
Detection of Genes Encoding Microbial Surface Component Recognizing Adhesive Matrix Molecules in Methicillin-Resistant Staphylococcus aureus Isolated from Pyoderma Patients
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of MRSA Strains
2.2. Antibiotic Susceptibility Testing
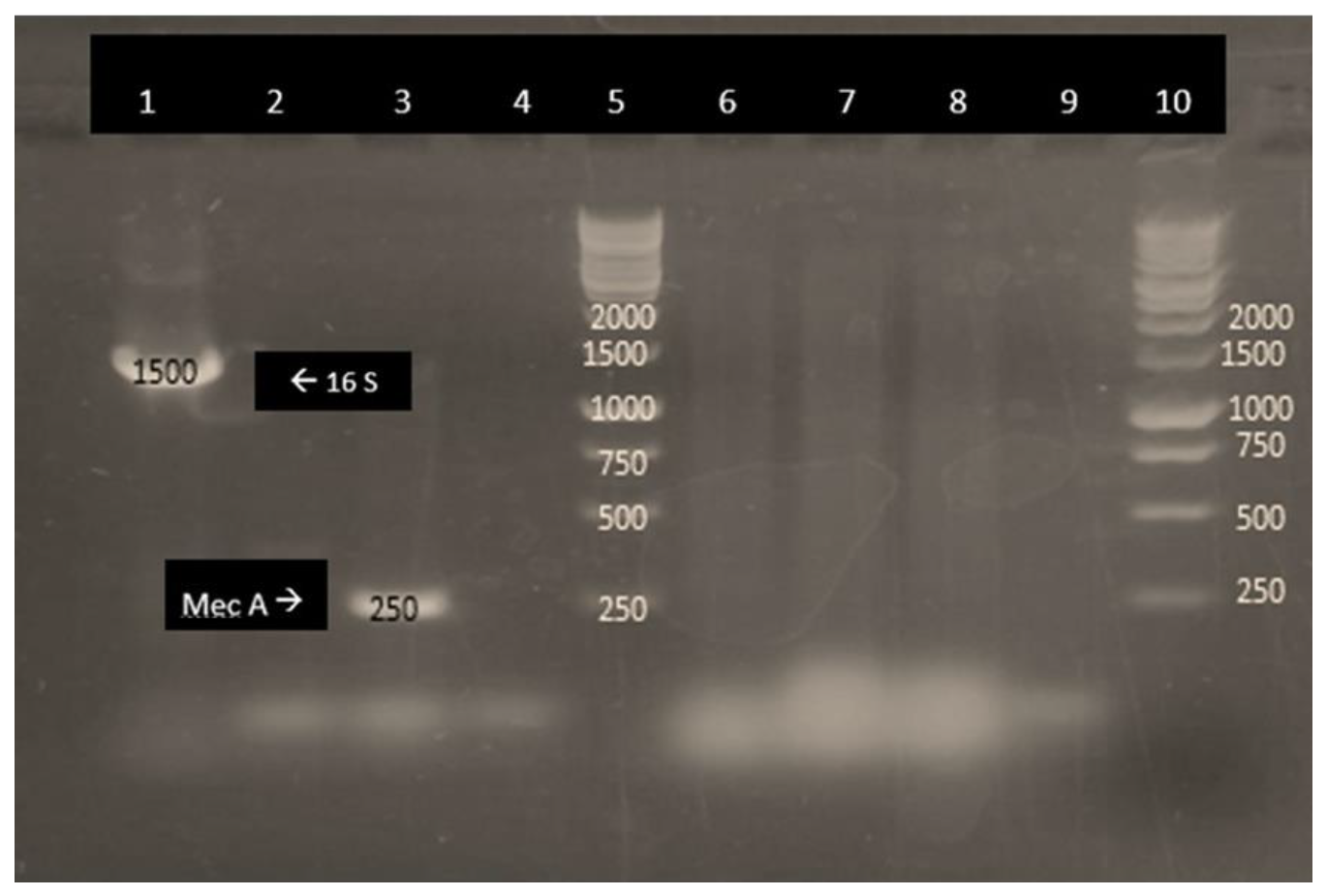
2.3. Amplification of MSCRAMMs
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Firoozeh, F.; Omidi, M.; Saffari, M.; Sedaghat, H.; Zibaei, M. Molecular Analysis of Methicillin-Resistant Staphylococcus aureus Isolates from Four Teaching Hospitals in Iran: The Emergence of Novel MRSA Clones. Antimicrob. Resist. Infect. Control 2020, 9, 1–8. [Google Scholar] [CrossRef]
- Romero, L.C.; de Souza da Cunha, M.d.L.R. Insights into the Epidemiology of Community-Associated Methicillin-Resistant Staphylococcus aureus in Special Populations and at the Community-Healthcare Interface. Braz. J. Infect. Dis. 2021, 25, 101636. [Google Scholar] [CrossRef]
- Gandhi, S.; Ojha, A.K.; Ranjan, K.P. Neelima Clinical and Bacteriological Aspects of Pyoderma. N. Am. J. Med. Sci. 2012, 4, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Speziale, P.; Pietrocola, G. The Multivalent Role of Fibronectin-Binding Proteins A and B (FnBPA and FnBPB) of Staphylococcus aureus in Host Infections. Front. Microbiol. 2020, 11, 2054. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. The MSCRAMM Family of Cell-Wall-Anchored Surface Proteins of Gram-Positive Cocci. Trends Microbiol. 2019, 27, 927–941. [Google Scholar] [CrossRef] [PubMed]
- Foster, C.E.; Kok, M.; Flores, A.R.; Minard, C.G.; Luna, R.A.; Lamberth, L.B.; Kaplan, S.L.; Hulten, K.G. Adhesin Genes and Biofilm Formation among Pediatric Staphylococcus aureus Isolates from Implant-Associated Infections. PLoS ONE 2020, 15, e0235115. [Google Scholar] [CrossRef] [PubMed]
- Atshan, S.S.; Nor Shamsudin, M.; Sekawi, Z.; Lung, L.T.T.; Hamat, R.A.; Karunanidhi, A.; Ali, A.M.; Ghaznavi-Rad, E.; Ghasemzadeh-Moghaddam, H.; Seng, J.S.C.; et al. Prevalence of Adhesion and Regulation of Biofilm-Related Genes in Different Clones of Staphylococcus aureus. J. Biomed. Biotechnol. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, V.K.; Rivera, J.J.; Smeds, E.; Ko, Y.P.; Bowden, M.G.; Wann, E.R.; Gurusiddappa, S.; Fitzgerald, J.R.; Höök, M. A Structural Model of the Staphylococcus aureus ClfA-Fibrinogen Interaction Opens New Avenues for the Design of Anti-Staphylococcal Therapeutics. PLoS Pathog. 2008, 4, e1000226. [Google Scholar] [CrossRef]
- Kot, B.; Sytykiewicz, H.; Sprawka, I. Expression of the Biofilm-Associated Genes in Methicillin-Resistant Staphylococcus aureus in Biofilm and Planktonic Conditions. Int. J. Mol. Sci. 2018, 19, 3487. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Zhang, G.; Sun, B.; Liu, S.; Wang, Y.; Gao, M.; Fan, Y.; Zhang, G.; Shi, G.; Kang, X. Rapid Detection of MecA and FemA Genes by Loop-Mediated Isothermal Amplification in a Microfluidic System for Discrimination of Different Staphylococcal Species and Prediction of Methicillin Resistance. Front. Microbiol. 2020, 11, 1487. [Google Scholar] [CrossRef]
- Vitali, L.A.; Petrelli, D.; Lamikanra, A.; Prenna, M.; Akinkunmi, E.O. Diversity of Antibiotic Resistance Genes and Staphylococcal Cassette Chromosome Mec Elements in Faecal Isolates of Coagulase-Negative Staphylococci from Nigeria. BMC Microbiol. 2014, 14, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.Y.C.; McDonald, M.I.; Holt, D.C.; Currie, B.J. Global Implications of the Emergence of Community-Associated Methicillin-Resistant Staphylococcus aureus in Indigenous Populations. Clin. Infect. Dis. 2008, 46, 1871–1878. [Google Scholar] [CrossRef] [Green Version]
- Udo, E.E. Community-Acquired Methicillin-Resistant Staphylococcus aureus: The New Face of an Old Foe? Med. Princ. Pract. 2013, 22, 20–29. [Google Scholar] [CrossRef]
- Jukes, L.; Mikhail, J.; Bome-Mannathoko, N.; Hadfield, S.J.; Harris, L.G.; El-Bouri, K.; Davies, A.P.; Mack, D. Rapid Differentiation of Staphylococcus aureus, Staphylococcus Epidermidis and Other Coagulase-Negative Staphylococci and Meticillin Susceptibility Testing Directly from Growth-Positive Blood Cultures by Multiplex Real-Time PCR. J. Med. Microbiol. 2010, 59, 1456–1461. [Google Scholar] [CrossRef]
- Lim, W.W.; Wu, P.; Bond, H.S.; Wong, J.Y.; Ni, K.; Hong, W.; Jit, M.; Cowling, B.J. Determinants of methicillin-resistant Staphylococcus aureus (MRSA) prevalence in the Asia-Pacific region: A systematic review and meta-analysis. J. Glob. Antimicrob. Resist. 2020, 16, 17–27. [Google Scholar] [CrossRef]
- Lade, H.; Joo, H.S.; Kim, J.S. Molecular Basis of Non-β-Lactam Antibiotics Resistance in Staphylococcus aureus. Antibiotics 2022, 11, 1378. [Google Scholar] [CrossRef]
- Watkins, R.R.; Holubar, M.; David, M.Z. Antimicrobial Resistance in Methicillin-Resistant Staphylococcus aureus to Newer Antimicrobial Agents. Antimicrob. Agents Chemother. 2019, 63, e01216-19. [Google Scholar] [CrossRef]
- Swathi, G.; Chandan, K.S.; Murgesh, S.B. Clinical and Bacteriological Study of Pyoderma with Reference to Antibiotic Susceptibility Profile. J. Pak. Assoc. Dermatol. 2019, 29, 83–88. [Google Scholar]
- Ghadage, D.; Sali, Y. Bacteriological Study of Pyoderma with Special Reference to Antibiotic Susceptibility to Newer Antibiotics. Indian J. Dermatol. Venereol. Leprol. 1999, 65, 177–181. [Google Scholar]
- Ghosh, S.; Sengupta, M.; Sarkar, S.; Pramanik, S.B.; Sengupta, M.; Bandyopadhyay, D. Bacteriologic Profile Along with Antimicrobial Susceptibility Pattern of Pediatric Pyoderma in Eastern India. Cureus 2022, 14. [Google Scholar] [CrossRef]
- Omidi, M.; Firoozeh, F.; Saffari, M.; Sedaghat, H.; Zibaei, M.; Khaledi, A. Ability of Biofilm Production and Molecular Analysis of Spa and Ica Genes among Clinical Isolates of Methicillin-Resistant Staphylococcus aureus. BMC Res. Notes 2020, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Yoon, E.J.; Kim, D.; Kim, J.W.; Lee, K.J.; Kim, H.S.; Kim, Y.R.; Shin, J.H.; Shin, J.H.; Shin, K.S.; et al. Ceftaroline Resistance by Clone-Specific Polymorphism in Penicillin-Binding Protein 2a of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2018, 62, e00485-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qodrati, M.; SeyedAlinaghi, S.A.; Manshadi, S.A.D.; Abdollahi, A.; Dadras, O. Antimicrobial Susceptibility Testing of Staphylococcus aureus Isolates from Patients at a Tertiary Hospital in Tehran, Iran, 2018–2019. Eur. J. Med. Res. 2022, 27, 152. [Google Scholar] [CrossRef] [PubMed]
- Habib, A.; Qadir, A. Frequency and Antibiotic Susceptibility Pattern of Community-Associated Methicillin-Resistant Staphylococcus aureus (CA-MRSA) in Uncomplicated Skin and Soft Tissue Infections. J. Coll. Physicians Surg. Pak. 2022, 32, 1398–1403. [Google Scholar] [CrossRef]
- Mehndiratta, P.; Bhalla, P.; Ahmed, A.; Sharma, Y. Molecular Typing of Methicillin-Resistant Staphylococcus aureus Strains by PCR-RFLP of SPA Gene: A Reference Laboratory Perspective. Indian J. Med. Microbiol. 2009, 27, 116–122. [Google Scholar] [CrossRef]
- Ahmad, M.K.; Asrar, A. Prevalence of Methicillin Resistant Staphylococcus aureus in Pyogenic Community and Hospital Acquired Skin and Soft Tissues Infections. J. Pak. Med. Assoc. 2014, 64, 892–895. [Google Scholar]
- Wongwanich, S.; Tishyadhigama, P.; Paisomboon, S.; Ohta, T. Epidemiological Analysis of Methicillin Resistant. Southeast Asia Microbiol. J. 2000, 31, 72–76. [Google Scholar]
- Zuniga, E.; Benites, N.R.; da Hora, A.S.; Mello, P.L.; Laes, M.A.; de Oliveira, L.A.R.; Brandão, P.E.; de Souza Silva, S.O.; Taniwaki, S.A.; Melville, P.A. Expression of Genes Encoding Resistance in Staphylococcus Spp. Isolated from Bovine Subclinical Mastitis in Brazil. J. Infect. Dev. Ctries. 2020, 14, 772–780. [Google Scholar] [CrossRef]




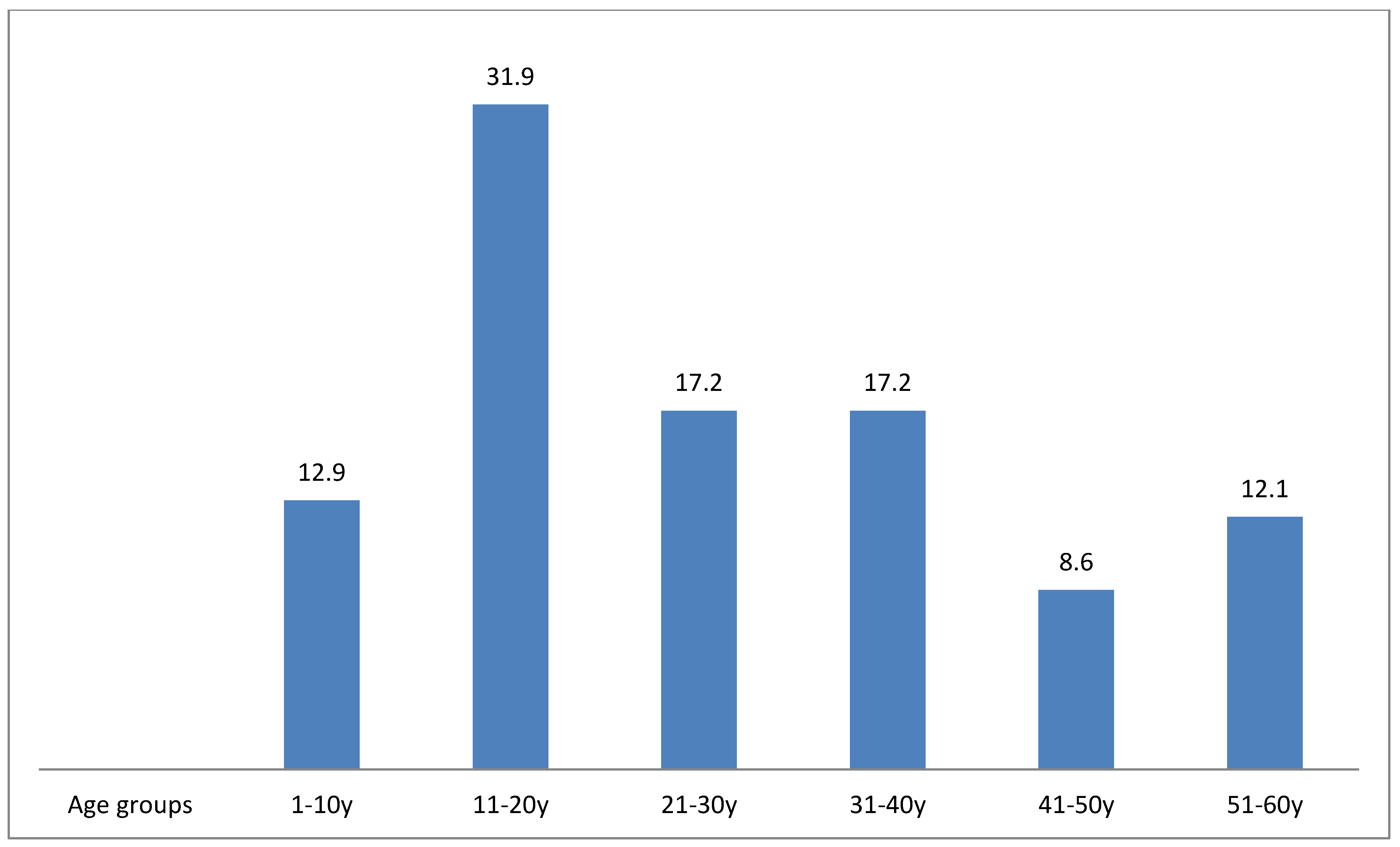{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | Frequency | Percent |
|---|---|---|
| AMC | 11 | 9.5 |
| AMC, SXT + Antifungal | 1 | 0.9 |
| AMC + FD | 1 | 0.9 |
| AMC + SXT | 27 | 23.3 |
| AMP, SXT | 2 | 1.7 |
| Antifungal | 3 | 2.6 |
| CD | 4 | 3.4 |
| CF | 11 | 9.5 |
| CF + VA | 2 | 1.7 |
| CF + SXT | 19 | 16.4 |
| CRO/SXT | 5 | 4.3 |
| LNZ | 1 | 0.9 |
| No | 16 | 13.8 |
| Steroids | 2 | 1.7 |
| SXT | 9 | 7.8 |
| VA, CF | 1 | 0.9 |
| Total | 116 | 100.0 |
| Antibiotics | Number of Sensitive Strains | Percent |
|---|---|---|
| Penicillin | 27 | 23.3 |
| Cefoxitin | 43 | 37.06 |
| Ceftaroline | 94 | 81.0 |
| Gentamicin | 96 | 82.8 |
| Erythromycin | 49 | 42.2 |
| Tetracycline | 53 | 45.7 |
| Ciprofloxacin | 44 | 37.9 |
| Clindamycin | 103 | 88.8 |
| Trimethoprim–sulfamethoxazole | 62 | 53.4 |
| Chloramphenicol | 103 | 88.8 |
| Linezolid | 116 | 100.0 |
| Rifampin | 108 | 93.1 |
| Antibiotic | Susceptibility | MSSA (43) | MRSA (73) | p-Value |
|---|---|---|---|---|
| Ceftaroline | R | 3 (6.97) | 19 (26.03) | 0.01 |
| S | 40 (93.03) | 54 (73.97) | ||
| Gentamicin | R | 5 (11.62) | 15 (20.55) | 0.2 |
| S | 38 (88.38) | 58 (79.45) | ||
| Erythromycin | R | 21 (48.84) | 46 (63.01) | 0.1 |
| S | 22 (51.16) | 27 (36.97) | ||
| Linezolid * | R | - | - | - |
| S | 43 (100) | 73 (100) | ||
| Rifampin | R | - | 8 (10.96) | 0.02 |
| S | 43 (100) | 65 (89.04) | ||
| Tetracycline | R | 18 (41.86) | 45 (61.64) | 0.03 |
| S | 25 (58.14) | 28 (38.36) | ||
| Ciprofloxacin | R | 14 (32.56) | 58 (79.45) | 0.00 |
| S | 29 (67.44) | 15 (20.55) | ||
| Clindamycin | R | - | 13 (17.81) | 0.00 |
| S | 43 (100) | 60 (82.19) | ||
| Trimethoprim sulfamethoxazole | R | 9 (20.93) | 45 (61.64) | 0.00 |
| S | 34 (79.07) | 28 (38.36) | ||
| Chloramphenicol | R | - | 13 (17.81) | 0.00 |
| S | 43 (100) | 60 (82.19) |
| Prevalence | Frequency | Percent | Valid Percent |
|---|---|---|---|
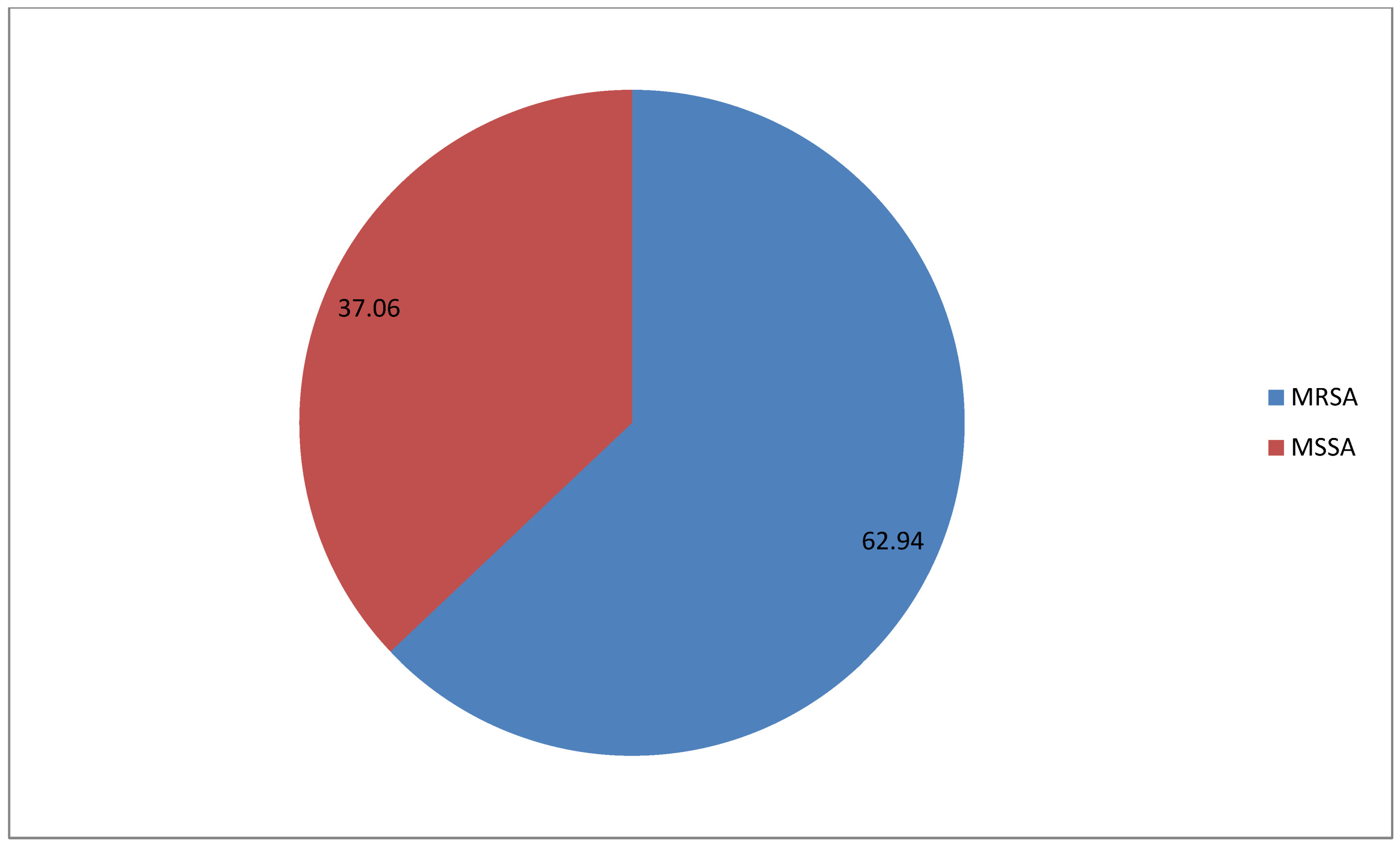
| Negative | 43 | 37.07 | 37.07 |
| Positive | 73 | 62.93 | 62.93 |
| Total | 116 | 100.0 | 100.0 |
| MSCRAMMs | Number | Percent |
|---|---|---|
| clfA | 115 | 99.1 |
| bbp | 116 | 100.0 |
| fnbA | 115 | 99.1 |
| fnbB | 116 | 100.0 |
| eno | 86 | 74.1 |
| fib | 85 | 73.3 |
| cna | 114 | 98.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alorabi, M.; Ejaz, U.; Khoso, B.K.; Uddin, F.; Mahmoud, S.F.; Sohail, M.; Youssef, M. Detection of Genes Encoding Microbial Surface Component Recognizing Adhesive Matrix Molecules in Methicillin-Resistant Staphylococcus aureus Isolated from Pyoderma Patients. Genes 2023, 14, 783. https://doi.org/10.3390/genes14040783
Alorabi M, Ejaz U, Khoso BK, Uddin F, Mahmoud SF, Sohail M, Youssef M. Detection of Genes Encoding Microbial Surface Component Recognizing Adhesive Matrix Molecules in Methicillin-Resistant Staphylococcus aureus Isolated from Pyoderma Patients. Genes. 2023; 14(4):783. https://doi.org/10.3390/genes14040783
Chicago/Turabian StyleAlorabi, Mohammed, Uroosa Ejaz, Bahram Khan Khoso, Fakhur Uddin, Samy F. Mahmoud, Muhammad Sohail, and Mona Youssef. 2023. "Detection of Genes Encoding Microbial Surface Component Recognizing Adhesive Matrix Molecules in Methicillin-Resistant Staphylococcus aureus Isolated from Pyoderma Patients" Genes 14, no. 4: 783. https://doi.org/10.3390/genes14040783