

DNA Barcoding, Phylogenetic Analysis and Secondary Structure Predictions of Nepenthes ampullaria, Nepenthes gracilis and Nepenthes rafflesiana

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
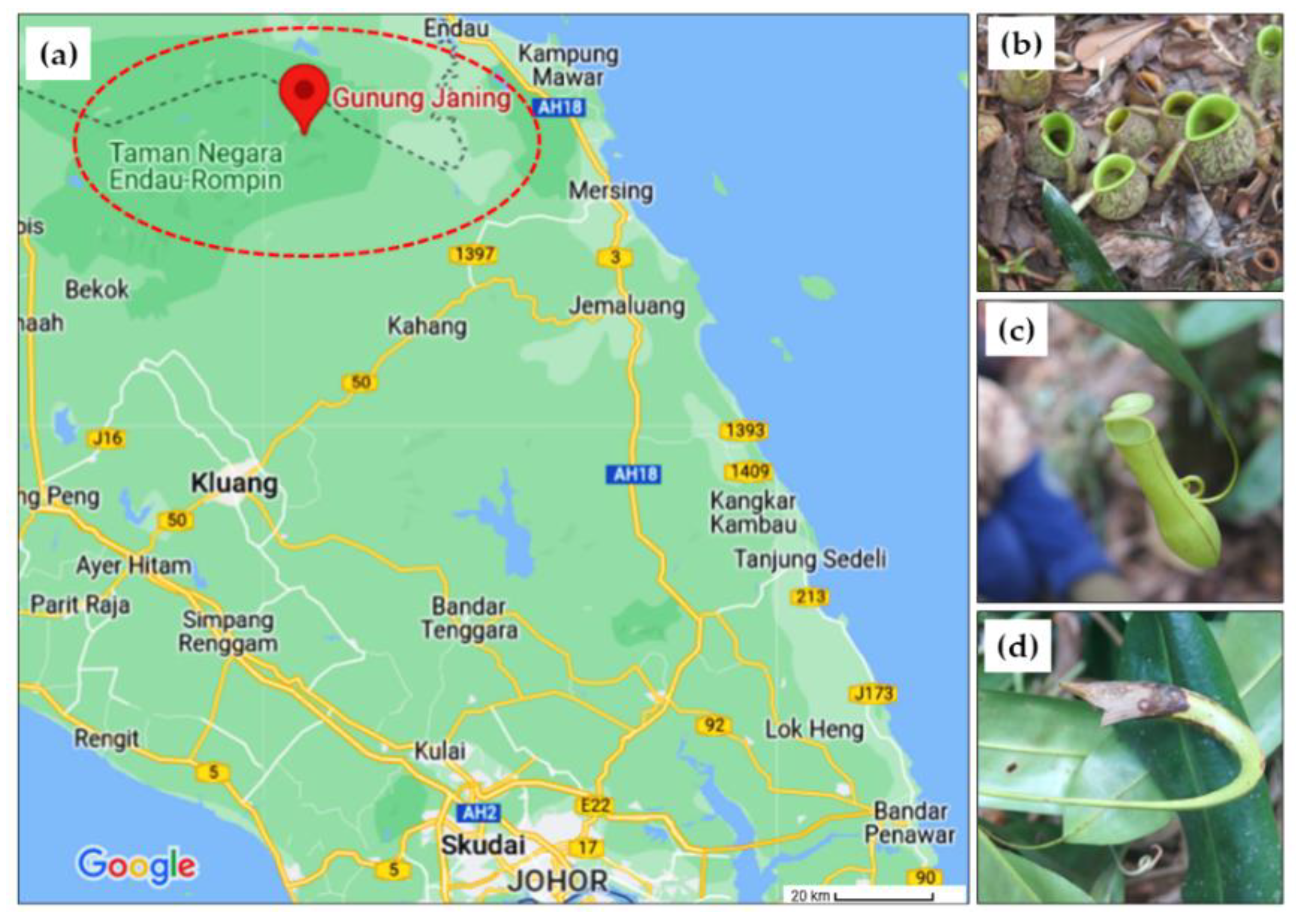
2.1. Sample Collection
2.2. Genomic DNA Extraction and PCR Amplification
2.3. Bioinformatics Analysis and Phylogenetic Tree
2.4. Secondary Structure Predictions
3. Results
3.1. Amplification, Sequencing, Multiple Sequence Alignment and Species Identification
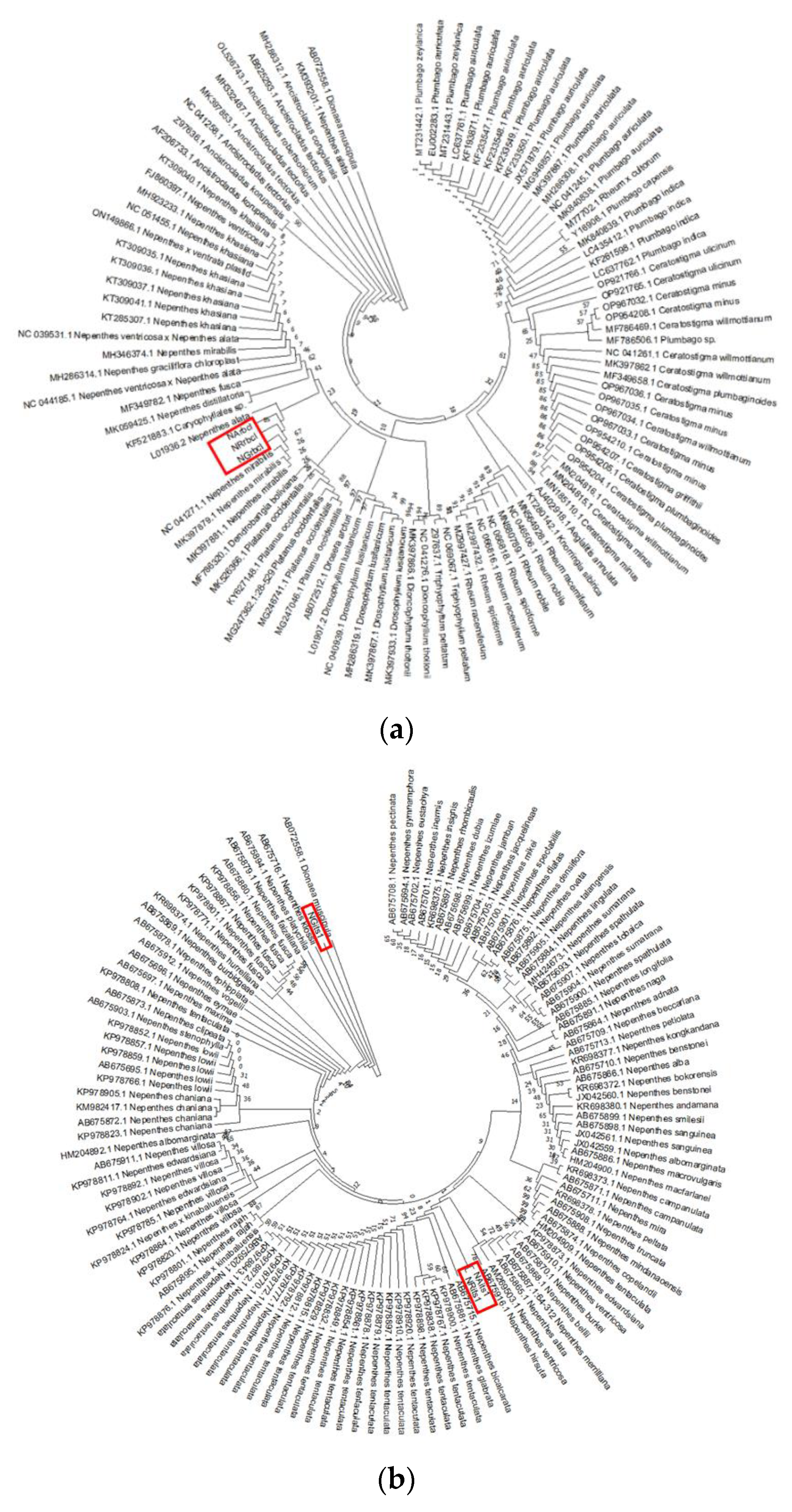
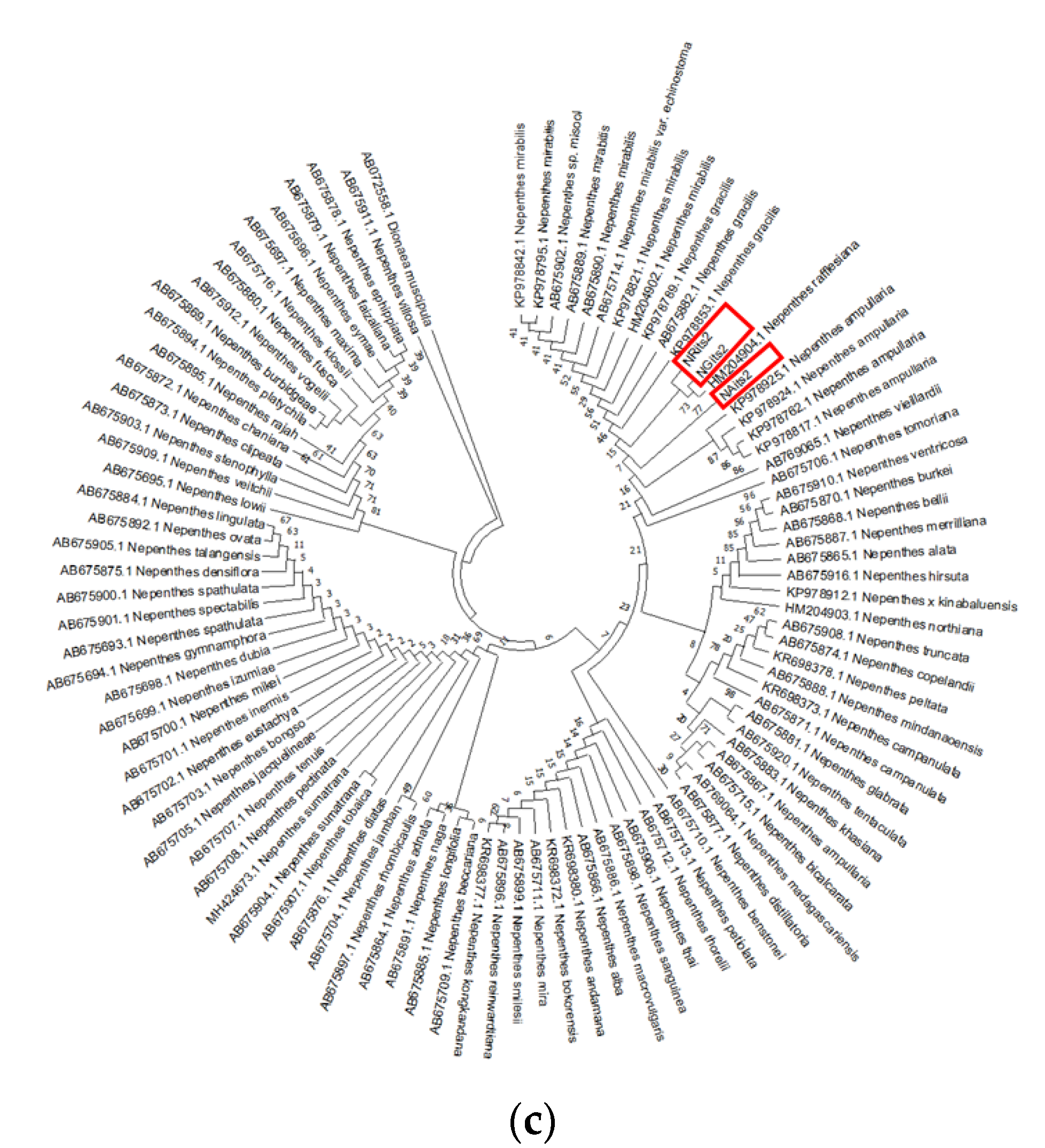
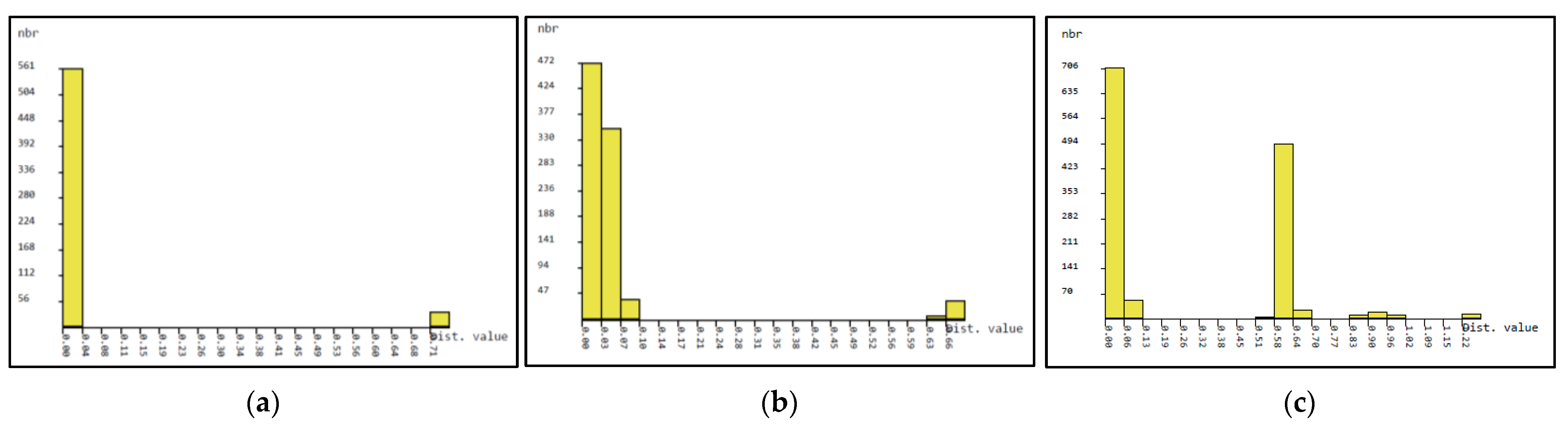
3.2. Phylogenetic Studies, Intraspecific Variation, Interspecific Divergence and DNA Barcoding Gap
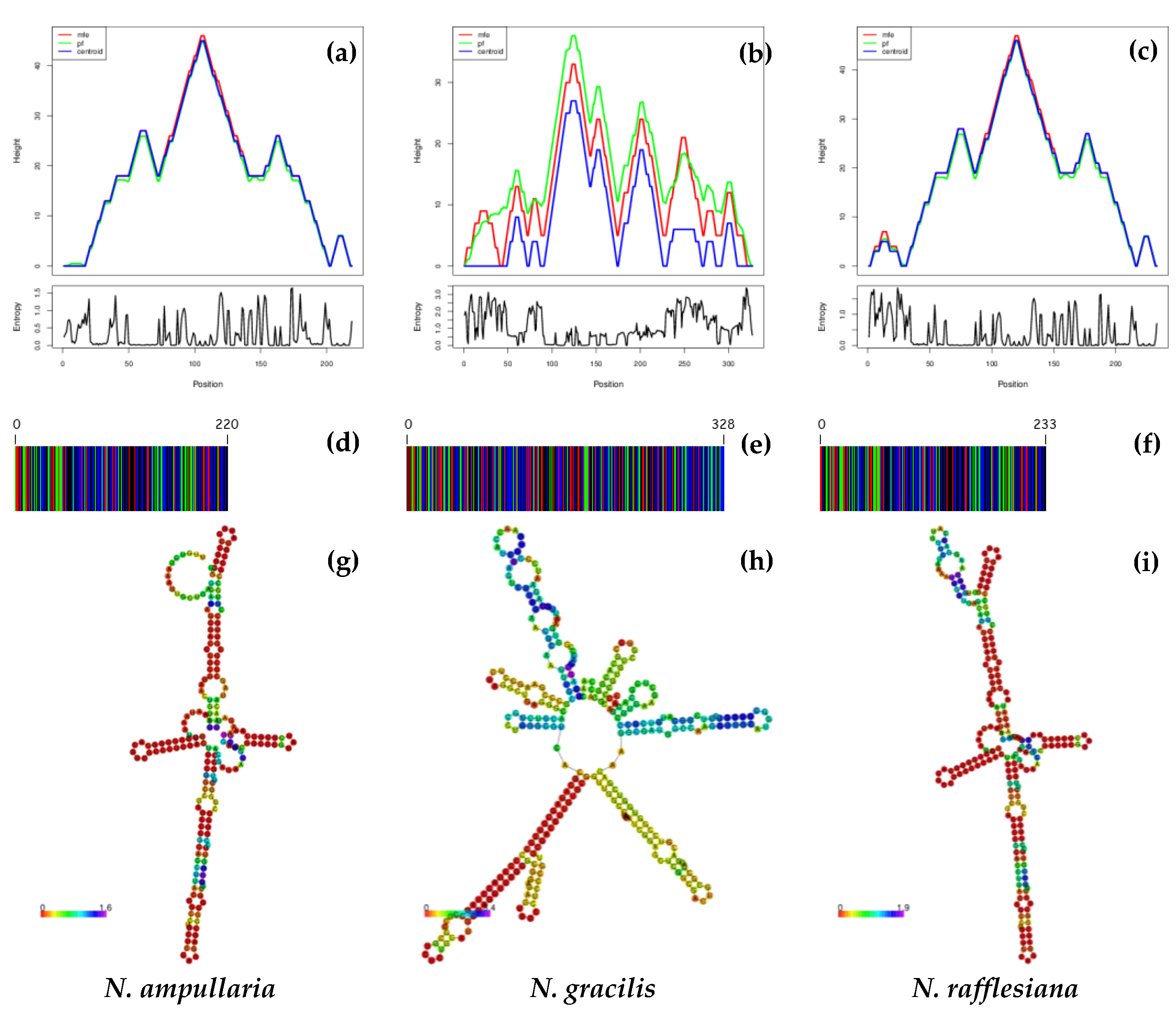
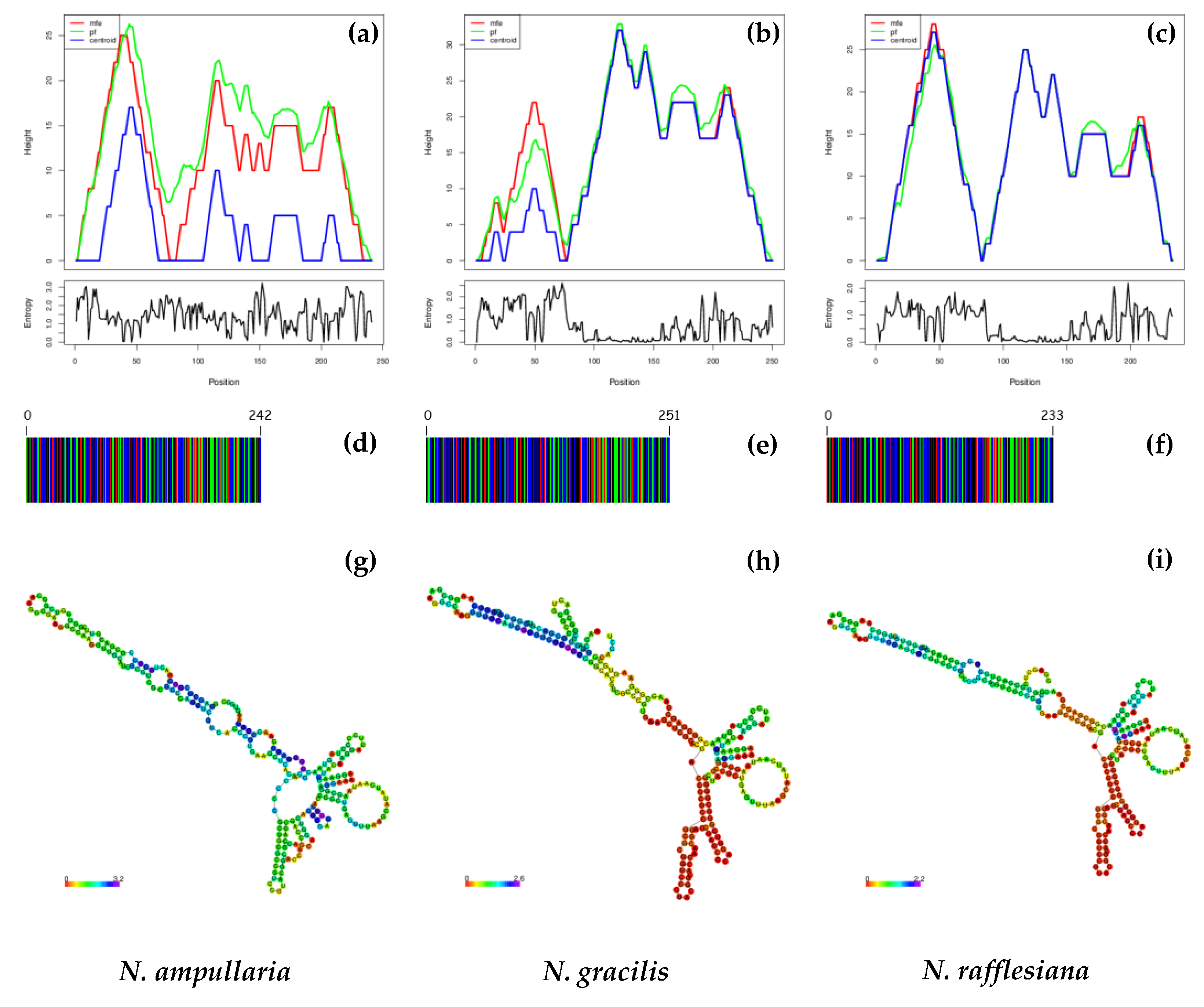
3.3. DNA Barcodes, ITS1 and ITS2 Secondary Structure Predictions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McPherson, S.; Robinson, A.S. Field Guide to the Pitcher Plants of Peninsular Malaysia and Indochina; Redfern Natural History: Poole, UK, 2012. [Google Scholar]
- Clarke, C. Nepenthes of Borneo; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 1997. [Google Scholar]
- Clarke, C. Nepenthes of Sumatra, and Peninsular Malaysia. In Nepenthes of Sumatra and Peninsular Malaysia; Natural History Publications (Borneo): Kota Kinabalu, Malaysia, 2001; p. 336. [Google Scholar]
- Cheek, M.; Jebb, M. Flora Malesiana, Series 1: Volume 15: Nepenthaceae. In Flora Malesiana; National Herbarium of the Netherlands: Wageningen, The Netherlands, 2001. [Google Scholar]
- Jebb, M.; Cheek, M. A skeletal revision of nepenthes (Nepenthaceae). Blumea J. Plant Taxon. Plant Geogr. 1997, 42, 1–106. [Google Scholar]
- Lestari, W.; Jumari, J.; Ferniah, R.S. Identification and Cluster Analysis of Pitcher Plant (Nepenthes spp.) from South Sumatera Indonesia. Biosaintifika J. Biol. Biol. Educ. 2018, 10, 245–251. [Google Scholar] [CrossRef]
- Phillipps, A.; Lamb, A. Pitcher-Plants of Borneo, 1st ed.; Royal Botanic Gardens: Sydney, Australia, 1996. [Google Scholar]
- Beveridge, N.G.P.; Rauch, C.; Ke, P.J.A.; van Vugt, R.R. A new way to identify living species of Nepenthes (Nepenthaceae ): More data needed ! Carniv. Plant Newsl. 2013, 42, 122–128. [Google Scholar] [CrossRef]
- Aung, H.H.; Chia, L.S.; Goh, N.K.; Chia, T.F.; Ahmed, A.A.; Pare, P.W.; Mabry, T.J. Phenolic constituents from the leaves of the carnivorous plant Nepenthes gracilis. Fitoterapia 2002, 73, 445–447. [Google Scholar] [CrossRef]
- Mainasara, M.M.; Fadzelly, M.; Bakar, A.; Mohamed, M.; Linatoc, A.C.; Sanusi, S.B. Ethnomedical Knowledge of Plants Used for the Treatment of Breast Cancer by Jakun community in Kampung Peta Endau Rompin Johor, Malaysia. J. Sci. Technol. 2017, 9, 42–49. [Google Scholar]
- Thao, N.P.; Luyen, B.T.T.; Koo, J.E.; Kim, S.; Koh, Y.S.; van Thanh, N.; Cuong, N.X.; van Kiem, P.; van Minh, C.; Kim, Y.H. In vitro anti-inflammatory components isolated from the carnivorous plant Nepenthes mirabilis (Lour.) Rafarin. Pharm. Biol. 2016, 54, 588–594. [Google Scholar] [CrossRef]
- Sabran, S.F.; Mohamed, M.; Abu Bakar, M.F. Ethnomedical Knowledge of Plants Used for the Treatment of Tuberculosis in Johor, Malaysia. Evid. -Based Complement. Altern. Med. 2016, 2016, 2850845. [Google Scholar] [CrossRef] [PubMed]
- Sanusi, S.B.; Abu Bakar, M.F.; Mohamed, M.; Sabran, S.F.; Mainasara, M.M. Ethnobotanical, phytochemical, and pharmacological properties of nepenthes species: A review. Asian J. Pharm. Clin. Res. 2017, 10, 16–19. [Google Scholar] [CrossRef]
- Schwallier, R.; de Boer, H.J.; Visser, N.; van Vugt, R.R.; Gravendeel, B. Traps as treats: A traditional sticky rice snack persisting in rapidly changing Asian kitchens. J. Ethnobiol. Ethnomed. 2015, 11, 24. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, M.; Chen, X.; Chen, X.; Hu, Y.; Gao, J.; Pan, W.; Xin, Y.; Wu, J.; Du, Y.; et al. Developing an efficient DNA barcoding system to differentiate between Lilium species. BMC Plant Biol. 2021, 21, 465. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, B.; Bhau, B.S. DNA barcoding of the genus Nepenthes (Pitcher plant): A preliminary assessment towards its identification. BMC Plant Biol. 2018, 18, 153. [Google Scholar] [CrossRef]
- Bunawan, H.; Yen, C.C.; Yaakop, S.; Noor, N.M. Phylogenetic inferences of Nepenthes species in Peninsular Malaysia revealed by chloroplast (trnL intron) and nuclear (ITS) DNA sequences. BMC Res. Notes 2017, 10, 67. [Google Scholar] [CrossRef]
- Acharya, G.C.; Mohanty, S.; Dasgupta, M.; Sahu, S.; Singh, S.; Koundinya, A.V.V.; Kumari, M.; Naresh, P.; Sahoo, M.R. Molecular Phylogeny, DNA Barcoding, and ITS2 Secondary Structure Predictions in the Medicinally Important Eryngium Genotypes of East Coast Region of India. Genes 2022, 13, 1678. [Google Scholar] [CrossRef]
- Liu, Z.W.; Gao, Y.Z.; Zhou, J. Molecular authentication of the medicinal species of Ligusticum (ligustici rhizoma et radix, “gao-ben”) by integrating non-coding internal transcribed spacer 2 (ITS2) and its secondary structure. Front. Plant Sci. 2019, 10, 429. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Kumar, A.; Rodrigues, V.; Prabhu, K.N.; Kaushik, A.; Mani, D.N.; Shukla, A.K.; Sundaresan, V. DNA barcoding coupled with secondary structure information enhances Achyranthes species resolution. J. Appl. Res. Med. Aromat. Plants 2020, 19, 100269. [Google Scholar] [CrossRef]
- Fadzil, N.F.; Wagiran, A.; Mohd Salleh, F.; Abdullah, S.; Izham, M.N.H. Authenticity and Detection of Eurycoma longifolia herbal products using Bar-High Resolution Melting Analysis. Genes 2018, 9, 408. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, D.O.; Speranskaya, A.S.; Ayginin, A.A.; Khafizov, K.; Krinitsina, A.A.; Fedotova, A.V.; Pozdyshev, D.V.; Shtratnikova, V.Y.; Kupriyanova, E.V.; Shipulin, G.A.; et al. Improved Protocols of ITS1-Based Metabarcoding and Their Application in the Analysis of Plant-Containing Products. Genes 2019, 10, 122. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Newmaster, S.G.; Fazekas, A.J.; Steeves, R.A.D.; Janovec, J. Testing candidate plant barcode regions in the Myristicaceae. Mol. Ecol. Resour. 2008, 8, 480–490. [Google Scholar] [CrossRef]
- Yu, H.; Wu, K.; Song, J.; Zhu, Y.; Yao, H.; Luo, K.; Dai, Y.; Xu, S.; Lin, Y. Expedient identification of Magnoliaceae species by DNA barcoding. Plant OMICS 2014, 7, 47–53. [Google Scholar]
- Hollingsworth, P.M. DNA barcoding plants in biodiversity hot spots: Progress and outstanding questions. Heredity 2008, 101, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and Using a Plant DNA Barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Müller, T.; Achtziger, M.; Seibel, P.N.; Dandekar, T.; Wolf, M. The internal transcribed spacer 2 database—A web server for (not only) low level phylogenetic analyses. Nucleic Acids Res. 2006, 34, 704–707. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Li, D.Z.; van der Bank, M.; Twyford, A.D. Telling plant species apart with DNA: From barcodes to genomes. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150338. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270 (suppl. 1), 96–99. [Google Scholar] [CrossRef]
- Fazekas, A.J.; Burgess, K.S.; Kesanakurti, P.R.; Graham, S.W.; Newmaster, S.G.; Husband, B.C.; Percy, D.M.; Hajibabaei, M.; Barrett, S.C.H. Multiple Multilocus DNA Barcodes from the Plastid Genome Discriminate Plant Species Equally Well. PLoS ONE 2008, 3, e2802. [Google Scholar] [CrossRef]
- Kress, W.J.; Erickson, D.L. A Two-Locus Global DNA Barcode for Land Plants: The Coding rbcL Gene Complements the Non-Coding trnH-psbA Spacer Region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- Sass, C.; Little, D.P.; Stevenson, D.W.; Specht, C.D. DNA barcoding in the Cycadales: Testing the potential of proposed barcoding markers for species identification of Cycads. PLoS ONE 2007, 2, e1154. [Google Scholar] [CrossRef]
- Ismail, M.; Ahmad, A.; Nadeem, M.; Javed, M.A.; Khan, S.H.; Khawaish, I.; Sthanadar, A.A.; Qari, S.H.; Alghanem, S.M.; Khan, K.A.; et al. Development of DNA barcodes for selected Acacia species by using rbcL and matK DNA markers. Saudi J. Biol. Sci. 2020, 27, 3735–3742. [Google Scholar] [CrossRef] [PubMed]
- Coleman, A.W. ITS2 is a double-edged tool for eukaryote evolutionary comparisons. Trends Genet. 2003, 19, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Edinburgh, R.B.G.; Group, C.P.W.; Hollingsworth, P.M.; Forrest, L.L.; Spouge, J.L.; Hajibabaei, M.; Ratnasingham, S.; van der Bank, M.; Chase, M.W.; Cowan, R.S.; et al. A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12794–12797. [Google Scholar]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Gielly, L.; Miquel, C.; Valentini, A.; Vermat, T.; Corthier, G.; Brochmann, C.; Willerslev, E. Power and limitations of the chloroplast trnL (UAA) intron for plant DNA barcoding. Nucleic Acids Res. 2007, 35, e14. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Jiang, M.; Shi, Y.; Jiao, K.; Shen, C.; Lu, J.; Ying, Q.; Wang, H. Application of the ribosomal DNA ITS2 region of physalis (Solanaceae): DNA barcoding and phylogenetic study. Front. Plant Sci. 2016, 7, 1047. [Google Scholar] [CrossRef]
- Anaz, M.; Sasidharan, N.; Remakanthan, A.; Dilsha, M.V. ITS2 and RNA secondary structure-based analysis reveals a clear picture on phylogeny of South Indian Salacia spp. Comput. Biol. Chem. 2021, 91, 107438. [Google Scholar] [CrossRef]
- Selvaraj, D.; Ramalingam, S. Identification of morphologically similar species of Tribulus (Zygophyllaceae) by employing DNA barcodes and rRNA secondary structures. Ecol. Genet. Genom. 2021, 18, 100072. [Google Scholar] [CrossRef]
- Coleman, A.W. Pan-eukaryote ITS2 homologies revealed by RNA secondary structure. Nucleic Acids Res. 2007, 35, 3322–3329. [Google Scholar] [CrossRef]
- Wolf, M.; Achtziger, M.; Schultz, J.; Dandekar, T.; Müller, T. Homology modeling revealed more than 20,000 rRNA internal transcribed spacer 2 (ITS2) secondary structures. RNA 2005, 11, 1616–1623. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Liu, D.; Zhang, H.; Zhang, Y. Molecular authentication of medicinal and edible plant Gnaphalium affine (cudweed herb, “Shu-qu-cao”) based on DNA barcode marker ITS2. Acta Physiol. Plant. 2021, 43, 119. [Google Scholar] [CrossRef]
- Biswal, D.K.; Debnath, M.; Konhar, R.; Yanthan, S.; Tandon, P. Phylogeny and Biogeography of Carnivorous Plant Family Nepenthaceae with Reference to the Indian Pitcher Plant Kepenthes Khasiana Reveals an Indian Subcontinent Origin of Nepenthes Colonization in Southeast Asia During the Miocene Epoch. Front. Ecol. Evol. 2018, 6, 108. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Steps | No. of Cycles | rbcL 1 | ITS1 2 | ITS2 3 | |||
|---|---|---|---|---|---|---|---|
| Temperature (°C) | Duration (s) | Temperature (°C) | Duration (s) | Temperature (°C) | Duration (s) | ||
| Denaturation | 35 | 98 | 10 | 98 | 10 | 98 | 10 |
| Annealing | 53 | 15 | 55 | 15 | 55 | 10 | |
| Elongation | 72 | 20 | 72 | 40 | 72 | 30 | |
| Hold | 1 | 4 | ∞ | 4 | ∞ | 4 | ∞ |
| Region | Primer Name | Primer Sequence (5′ to 3′) | Primer Length (bp) | Reference |
|---|---|---|---|---|
| rbcL | Rbcla_fwd | ATGTCACCACAAACAGAGACTAAAGC | 26 | [21] |
| Rbclb_rvs | GTAAAATCAAGTCCACCRCG | 20 | ||
| ITS1 | ITS1_fwd | GGAAGGAGAAGTCGTAACAAGG | 22 | [21] |
| ITS1_rvs | AGATATCCGTTGCCGAGAGT | 20 | ||
| ITS2 | ITS2_fwd | GGGGCGGATATTGGCCTCCCCTTG | 24 | [22] |
| ITS2_rvs | GACGCTTCTCCAGACTACAAT | 21 |
| Barcode Genes | DNA Best-Fit Substitution Model |
|---|---|
| rbcL | Kimura-2-parameter |
| ITS1 | Tamura-3-parameter |
| ITS2 | Kimura-2-parameter + γ distribution |
| Barcode Genes | Sample ID | Scientific Name | Accession Number | E Value | Query Coverage (%) | Percent Identity (%) |
|---|---|---|---|---|---|---|
| rbcL | NArbcL | N. mirabilis | NC_041271.1 | 0.0 | 100 | 100 |
| NGrbcL | N. mirabilis | NC_041271.1 | 0.0 | 100 | 100 | |
| NRrbcL | N. mirabilis | NC_041271.1 | 0.0 | 100 | 100 | |
| ITS1 | NAits1 | Nepenthes x intermedia | HM204899.1 | 1e-103 | 100 | 98.64 |
| NGits1 | N. ventricosa | AB675910.1 | 2e-168 | 100 | 100 | |
| NRits1 | Nepenthes x intermedia | HM204899.1 | 2e-116 | 100 | 100 | |
| ITS2 | NAits2 | N. gracilis | AB675882.1 | 4e-104 | 100 | 95.85 |
| NGits2 | N. gracilis | AB675882.1 | 2e-122 | 100 | 99.20 | |
| NRits2 | N. gracilis | AB675882.1 | 5e-113 | 100 | 99.14 |
| DNA Barcode | Species | AT (bp) | GC (bp) | Total (bp) | AT (%) | GC (%) |
|---|---|---|---|---|---|---|
| rbcL | N. gracilis | 280 | 229 | 509 | 55 | 45 |
| N. ampullaria | 278 | 224 | 502 | 55.4 | 44.6 | |
| N. rafflesiana | 278 | 224 | 502 | 55.4 | 44.6 | |
| ITS2 | N. gracilis | 85 | 114 | 250 | 34.0 | 66.0 |
| N. ampullaria | 89 | 152 | 241 | 36.9 | 63.1 | |
| N. rafflesiana | 78 | 155 | 233 | 33.5 | 66.5 | |
| ITS1 | N. gracilis | 111 | 216 | 327 | 33.9 | 66.1 |
| N. ampullaria | 69 | 150 | 219 | 31.5 | 68.5 | |
| N. rafflesiana | 76 | 157 | 233 | 32.6 | 67.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saidon, N.A.; Wagiran, A.; Samad, A.F.A.; Mohd Salleh, F.; Mohamed, F.; Jani, J.; Linatoc, A.C. DNA Barcoding, Phylogenetic Analysis and Secondary Structure Predictions of Nepenthes ampullaria, Nepenthes gracilis and Nepenthes rafflesiana. Genes 2023, 14, 697. https://doi.org/10.3390/genes14030697
Saidon NA, Wagiran A, Samad AFA, Mohd Salleh F, Mohamed F, Jani J, Linatoc AC. DNA Barcoding, Phylogenetic Analysis and Secondary Structure Predictions of Nepenthes ampullaria, Nepenthes gracilis and Nepenthes rafflesiana. Genes. 2023; 14(3):697. https://doi.org/10.3390/genes14030697
Chicago/Turabian StyleSaidon, Nur Azreen, Alina Wagiran, Abdul Fatah A. Samad, Faezah Mohd Salleh, Farhan Mohamed, Jaeyres Jani, and Alona C Linatoc. 2023. "DNA Barcoding, Phylogenetic Analysis and Secondary Structure Predictions of Nepenthes ampullaria, Nepenthes gracilis and Nepenthes rafflesiana" Genes 14, no. 3: 697. https://doi.org/10.3390/genes14030697