
Iroquois Family Genes in Gastric Carcinogenesis: A Comprehensive Review

Abstract
:1. Introduction
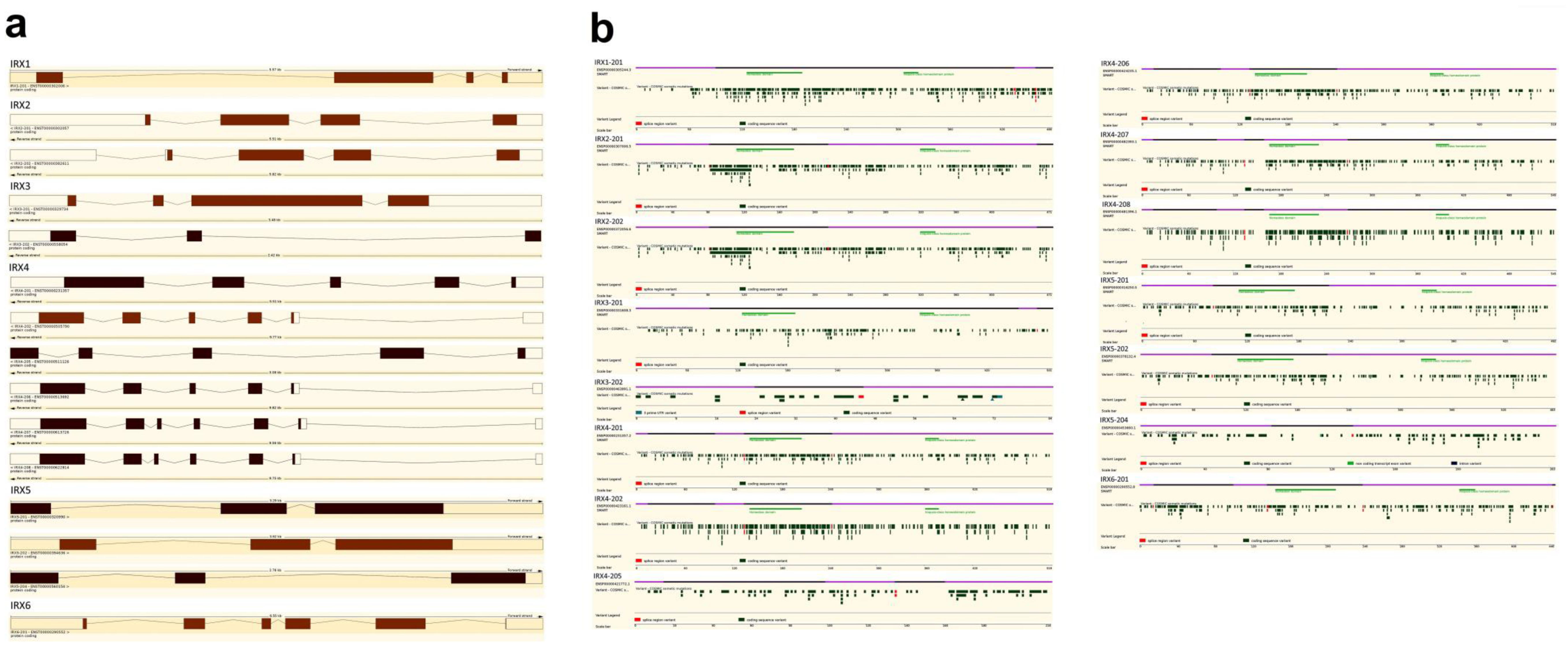
2. Overview of the Iroquois Gene Family
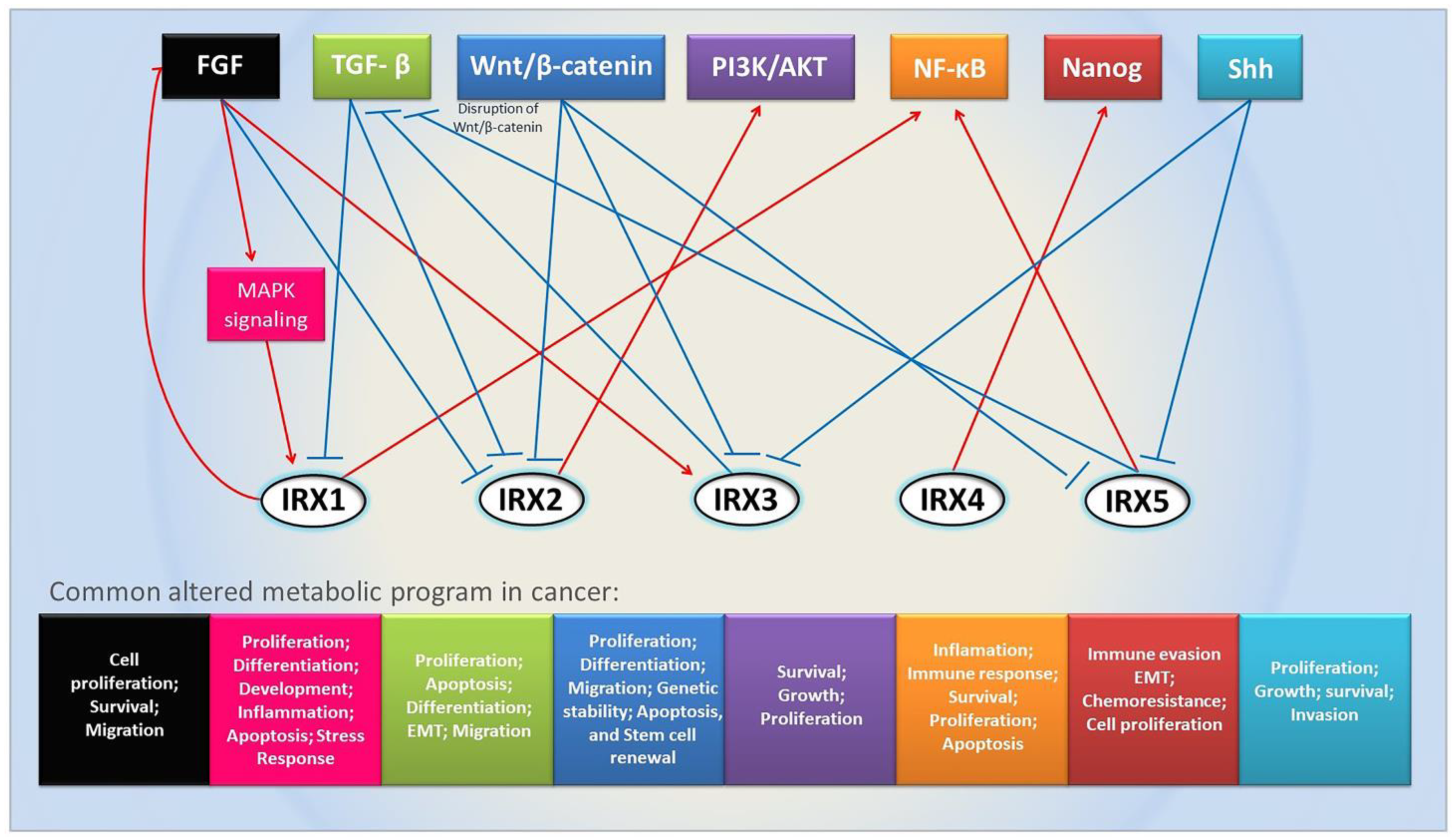
3. Important Pathways Related to Iroquois Genes
4. Iroquois Family Genes in Cancer
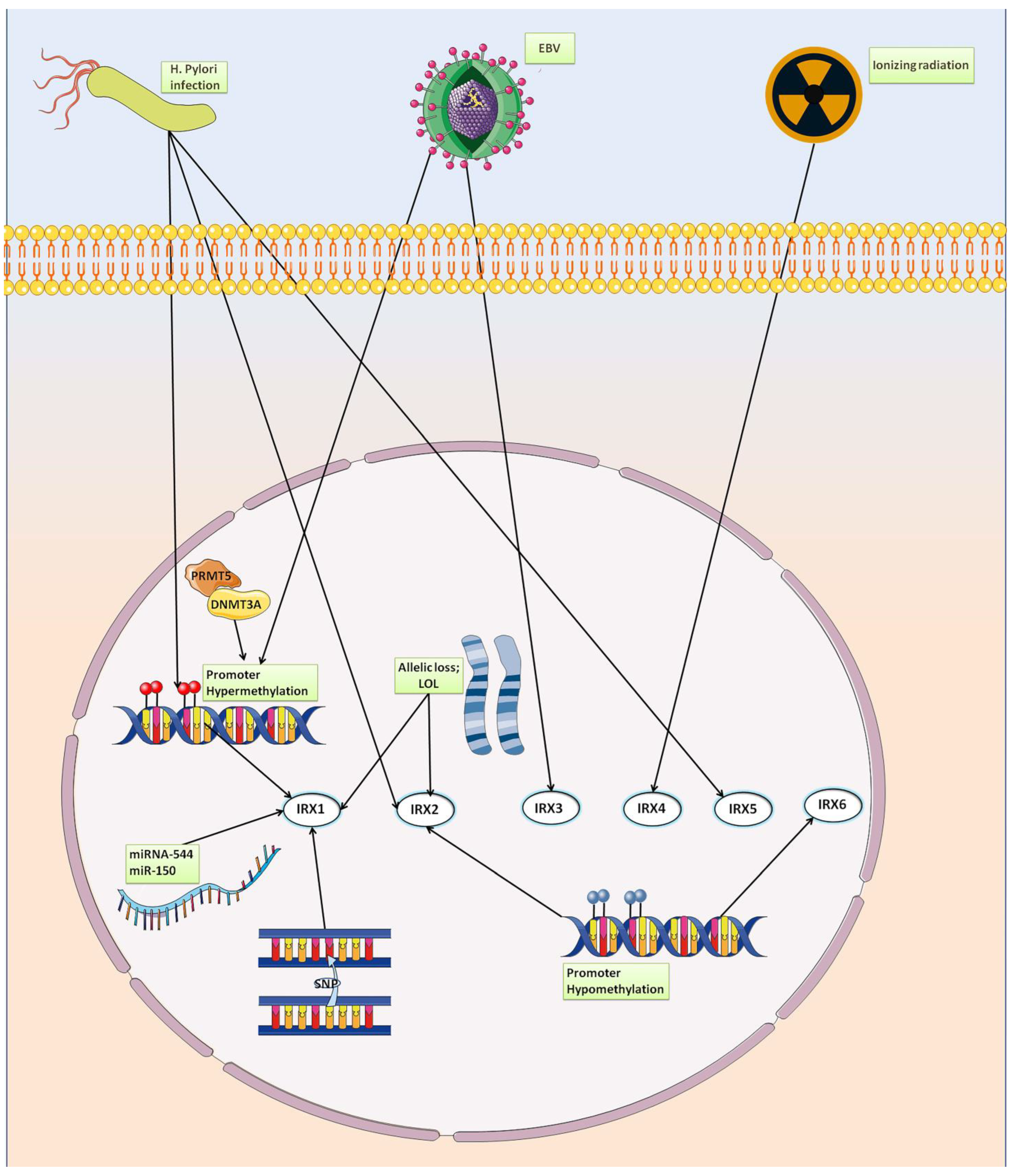
5. Iroquois Family Genes in Gastric Cancer
5.1. IRX1
5.2. IRX2
5.3. IRX3
5.4. IRX4
5.5. IRX5
5.6. IRX6
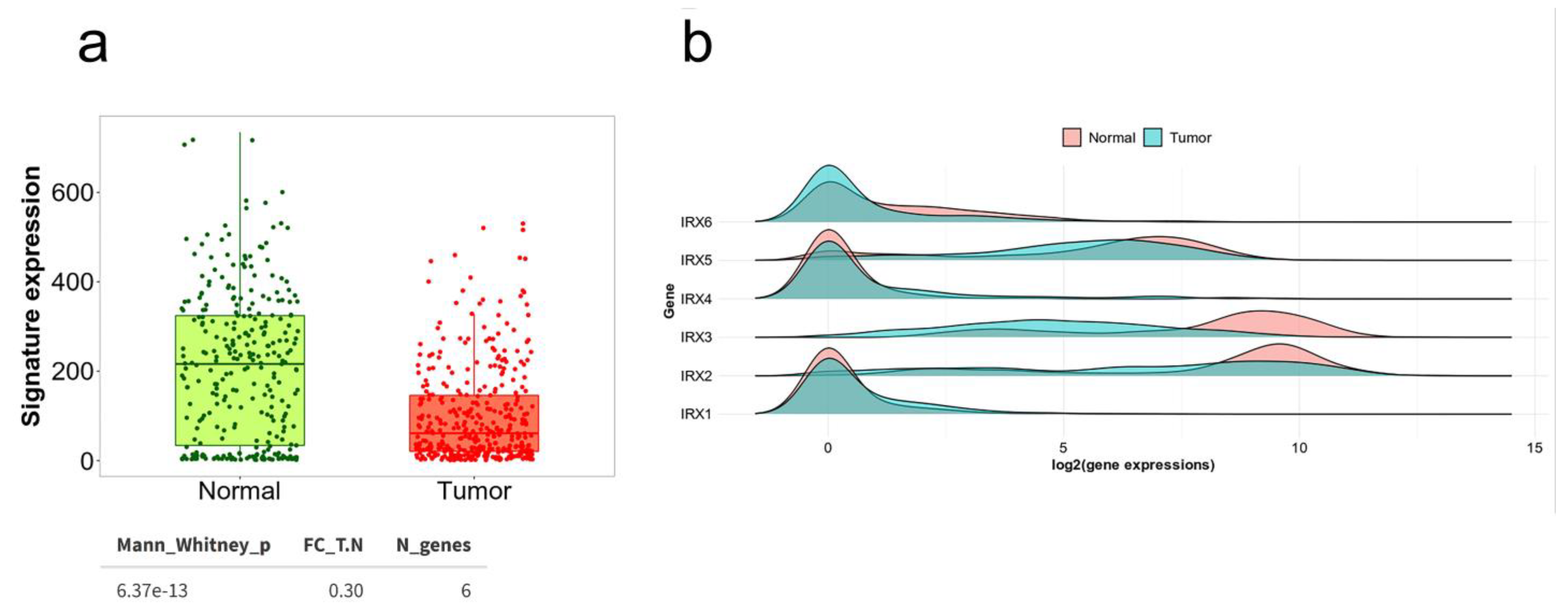
6. Status of Iroquois Family Genes in GC Patient Databases
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Nagini, S. Carcinoma of the Stomach: A Review of Epidemiology, Pathogenesis, Molecular Genetics and Chemoprevention. World J. Gastrointest. Oncol. 2012, 4, 156. [Google Scholar] [CrossRef]
- Kumar, R.K.; Raj, S.S.; Shankar, E.M.; Ganapathy, E.; Ebrahim, A.S.; Farooq, S.M. Gastric Carcinoma: A Review on Epidemiology, Current Surgical and Chemotherapeutic Options; InTech Open: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric Cancer: Descriptive Epidemiology, Risk Factors, Screening, and Prevention. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, X.; Tang, B.; Li, B.-S.; Xie, R.; Hu, C.-J.; Luo, G.; Qin, Y.; Dong, H.; Yang, S.-M. Helicobacter Pylori Virulence Factor CagA Promotes Tumorigenesis of Gastric Cancer via Multiple Signaling Pathways. Cell Commun. Signal. 2015, 13, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LAURÉN, P. The two histological main types of gastric carcinoma: Diffuse and so-called intestinal-type carcinoma. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.S.; Kim, W.H. Epstein-Barr Virus in Human Malignancy: A Special Reference to Epstein-Barr Virus Associated Gastric Carcinoma. Cancer Res. Treat. 2005, 37, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.; Kang, M.-S.; Kim, K.-M. Epstein-Barr Virus-Associated Gastric Carcinoma and Specific Features of the Accompanying Immune Response. J. Gastric Cancer 2016, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Jia, K.; Lv, H.; Wang, S.-Q.; Wu, Y.; Lei, H.; Chen, X. EBV-Positive Gastric Cancer: Current Knowledge and Future Perspectives. Front. Oncol. 2020, 10, 583463. [Google Scholar] [CrossRef]
- do Rosário Pinheiro, D.; Ferreira, W.A.S.; Barros, M.B.L.; Araújo, M.D.; Rodrigues-Antunes, S.; do Nascimento Borges, B. Perspectives on New Biomarkers in Gastric Cancer: Diagnostic and Prognostic Applications. World J. Gastroenterol. 2014, 20, 11574–11585. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-S. MicroRNAs as Potential Biomarkers for Gastric Cancer. World J. Gastroenterol. 2014, 20, 12007. [Google Scholar] [CrossRef]
- Dang, Y.; Lan, F.; Ouyang, X.; Wang, K.; Lin, Y.; Yu, Y.; Wang, L.; Wang, Y.; Huang, Q. Expression and Clinical Significance of Long Non-Coding RNA HNF1A-AS1 in Human Gastric Cancer. World J. Surg. Oncol. 2015, 13, 302. [Google Scholar] [CrossRef] [Green Version]
- Binato, R.; Santos, E.C.; Boroni, M.; Demachki, S.; Assumpção, P.; Abdelhay, E. A Common Molecular Signature of Intestinal-Type Gastric Carcinoma Indicates Processes Related to Gastric Carcinogenesis. Oncotarget 2017, 9, 7359–7371. [Google Scholar] [CrossRef] [Green Version]
- Santos, E.C.; Gomes, R.B.; Fernandes, P.V.; Ferreira, M.A.; Abdelhay, E.S.F.W. The Protein-Protein Interaction Network of Intestinal Gastric Cancer Patients Reveals Hub Proteins with Potential Prognostic Value. Cancer Biomark. 2021, 33, 83–96. [Google Scholar] [CrossRef]
- Burglin, T.R. Analysis of TALE Superclass Homeobox Genes (MEIS, PBC, KNOX, Iroquois, TGIF) Reveals a Novel Domain Conserved between Plants and Animals. Nucleic Acids Res. 1997, 25, 4173–4180. [Google Scholar] [CrossRef] [Green Version]
- Cavodeassi, F.; Modolell, J.; Gómez-Skarmeta, J.L. The Iroquois Family of Genes: From Body Building to Neural Patterning. Development 2001, 128, 2847–2855. [Google Scholar] [CrossRef]
- Dambly-Chaudière, C.; Leyns, L. The Determination of Sense Organs in Drosophila: A Search for Interacting Genes. Int. J. Dev. Biol. 1992, 36, 85–91. [Google Scholar]
- Gomez-Skarmeta, J.L.; Modolell, J. Araucan and Caupolican Provide a Link between Compartment Subdivisions and Patterning of Sensory Organs and Veins in the Drosophila Wing. Genes Dev. 1996, 10, 2935–2945. [Google Scholar] [CrossRef] [Green Version]
- Bellefroid, E.J. Xiro3 Encodes a Xenopus Homolog of the Drosophila Iroquois Genes and Functions in Neural Specification. EMBO J. 1998, 17, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Skarmeta, J.; de La Calle-Mustienes, E.; Modolell, J. The Wnt-Activated Xiro1 Gene Encodes a Repressor That Is Essential for Neural Development and Downregulates Bmp4. Development 2001, 128, 551–560. [Google Scholar] [CrossRef]
- Dildrop, R.; Rüther, U. Organization of Iroquois Genes in Fish. Dev. Genes Evol. 2004, 214, 267–276. [Google Scholar] [CrossRef]
- Feijóo, C.G.; Manzanares, M.; de la Calle-Mustienes, E.; Gómez-Skarmeta, J.L.; Allende, M.L. The Irx Gene Family in Zebrafish: Genomic Structure, Evolution and Initial Characterization of Irx5b. Dev. Genes Evol. 2004, 214, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Gehring, W.J. Exploring the Homeobox. Gene 1993, 135, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Bürglin, T.R. A Comprehensive Classification of Homeobox Genes. Guideb. Homeobox Genes 1994, 25, 25–71. [Google Scholar]
- Bürglin, T.R. Homeodomain Proteins. Encycl. Mol. Cell Biol. Mol. Med. 2005, 6, 179–222. [Google Scholar] [CrossRef]
- Boncinelli, E. Homeobox Genes and Disease. Curr. Opin. Genet. Dev. 1997, 7, 331–337. [Google Scholar] [CrossRef]
- Mukherjee, K.; Bürglin, T.R. Comprehensive Analysis of Animal TALE Homeobox Genes: New Conserved Motifs and Cases of Accelerated Evolution. J. Mol. Evol. 2007, 65, 137–153. [Google Scholar] [CrossRef]
- Jordan, K.C.; Clegg, N.J.; Blasi, J.A.; Morimoto, A.M.; Sen, J.; Stein, D.; McNeill, H.; Deng, W.-M.; Tworoger, M.; Ruohola-Baker, H. The Homeobox Gene Mirror Links EGF Signalling to Embryonic Dorso-Ventral Axis Formation through Notch Activation. Nat. Genet. 2000, 24, 429–433. [Google Scholar] [CrossRef]
- Netter, S.; Fauvarque, M.-O.; del Corral, R.D.; Dura, J.-M.; Coen, D. White+ Transgene Insertions Presenting a Dorsal/Ventral Pattern Define a Single Cluster of Homeobox Genes That Is Silenced by the Polycomb-Group Proteins in Drosophila Melanogaster. Genetics 1998, 149, 257–275. [Google Scholar] [CrossRef]
- Perovic, S.; Schroder, H.C.; Sudek, S.; Grebenjuk, V.A.; Batel, R.; Stifanic, M.; Muller, I.M.; Muller, W.E.G. Expression of One Sponge Iroquois Homeobox Gene in Primmorphs from Suberites Domuncula during Canal Formation. Evol. Dev. 2003, 5, 240–250. [Google Scholar] [CrossRef]
- Larroux, C.; Luke, G.N.; Koopman, P.; Rokhsar, D.S.; Shimeld, S.M.; Degnan, B.M. Genesis and Expansion of Metazoan Transcription Factor Gene Classes. Mol. Biol. Evol. 2008, 25, 980–996. [Google Scholar] [CrossRef] [Green Version]
- Irimia, M.; Maeso, I.; Garcia-Fernandez, J. Convergent Evolution of Clustering of Iroquois Homeobox Genes across Metazoans. Mol. Biol. Evol. 2008, 25, 1521–1525. [Google Scholar] [CrossRef] [Green Version]
- Peters, T.; Dildrop, R.; Ausmeier, K.; Rüther, U. Organization of Mouse Iroquois Homeobox Genes in Two Clusters Suggests a Conserved Regulation and Function in Vertebrate Development. Genome Res. 2000, 10, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Bosse, A.; Stoykova, A.; Nieselt-Struwe, K.; Chowdhury, K.; Copeland, N.G.; Jenkins, N.A.; Gruss, P. Identification of a Novel Mouse Iroquois Homeobox Gene, Irx5, and Chromosomal Localisation of All Members of the Mouse Iroquois Gene Family. Dev. Dyn. 2000, 218, 160–174. [Google Scholar] [CrossRef]
- Gómez-Skarmeta, J.L.; Glavic, A.; de la Calle-Mustienes, E.; Modolell, J.; Mayor, R. Xiro, a Xenopus Homolog of the Drosophila Iroquois Complex Genes, Controls Development at the Neural Plate. EMBO J. 1998, 17, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, P.; Rodríguez-Seguel, E.; Fernández-González, A.; Rubio, R.; Gómez-Skarmeta, J.L. A Dual Requirement for Iroquois Genes during Xenopus Kidney Development. Development 2008, 135, 3197–3207. [Google Scholar] [CrossRef] [Green Version]
- Reggiani, L.; Raciti, D.; Airik, R.; Kispert, A.; Brändli, A.W. The Prepattern Transcription Factor Irx3 Directs Nephron Segment Identity. Genes Dev. 2007, 21, 2358–2370. [Google Scholar] [CrossRef] [Green Version]
- Schwab, K.; Hartman, H.A.; Liang, H.-C.; Aronow, B.J.; Patterson, L.T.; Potter, S.S. Comprehensive Microarray Analysis of Hoxa11/Hoxd11 Mutant Kidney Development. Dev. Biol. 2006, 293, 540–554. [Google Scholar] [CrossRef] [Green Version]
- van Tuyl, M.; Liu, J.; Groenman, F.; Ridsdale, R.; Han, R.N.N.; Venkatesh, V.; Tibboel, D.; Post, M. Iroquois Genes Influence Proximo-Distal Morphogenesis during Rat Lung Development. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L777–L789. [Google Scholar] [CrossRef] [Green Version]
- Ragvin, A.; Moro, E.; Fredman, D.; Navratilova, P.; Drivenes, Ø.; Engström, P.G.; Alonso, M.E.; Mustienes, E.; de la Calle Mustienes, E.; Skarmeta, J.L.G.; et al. Long-Range Gene Regulation Links Genomic Type 2 Diabetes and Obesity Risk Regions to HHEX, SOX4, and IRX3. Proc. Natl. Acad. Sci. USA 2009, 107, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.W.; Chow, R.L.; Lebel, M.; Sakuma, R.; Cheung, H.O.-L.; Thanabalasingham, V.; Zhang, X.; Bruneau, B.G.; Birch, D.G.; Hui, C.; et al. The Iroquois Homeobox Gene, Irx5, Is Required for Retinal Cone Bipolar Cell Development. Dev. Biol. 2005, 287, 48–60. [Google Scholar] [CrossRef]
- Jorgensen, J.S.; Gao, L. Irx3 Is Differentially Up-Regulated in Female Gonads during Sex Determination. Gene Expr. Patterns 2005, 5, 756–762. [Google Scholar] [CrossRef]
- Kim, K.-H.; Rosen, A.; Bruneau, B.G.; Hui, C.; Backx, P.H. Iroquois Homeodomain Transcription Factors in Heart Development and Function. Circ. Res. 2012, 110, 1513–1524. [Google Scholar] [CrossRef] [Green Version]
- Christoffels, V.M.; Keijser, A.G.; Houweling, A.C.; Clout, D.E.; Moorman, A.F. Patterning the Embryonic Heart: Identification of Five Mouse Iroquois Homeobox Genes in the Developing Heart. Dev. Biol. 2000, 224, 263–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, F.; Allen, J.E.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Bennett, R.; et al. Ensembl 2022. Nucleic Acids Res. 2022, 50, D988–D995. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in the Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Rando, M.; Tutor, A.S.; Prieto-Sánchez, S.; González-Pérez, E.; Barrios, N.; Letizia, A.; Martín, P.; Campuzano, S.; Ruiz-Gómez, M. Drosophila Araucan and Caupolican Integrate Intrinsic and Signalling Inputs for the Acquisition by Muscle Progenitors of the Lateral Transverse Fate. PLoS Genet. 2011, 7, e1002186. [Google Scholar] [CrossRef] [Green Version]
- Barrios, N.; Campuzano, S. Expanding the Iroquois Genes Repertoire: A Non-Transcriptional Function in Cell Cycle Progression. Fly 2015, 9, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.C.; Clurman, B.E. Cyclin E in Normal and Neoplastic Cell Cycles. Oncogene 2005, 24, 2776–2786. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, K.; Nishihara, S.; Kamimura, M.; Shiraishi, T.; Otoguro, T.; Uehara, M.; Maeda, Y.; Ogura, K.; Lumsden, A.; Ogura, T. The Prepattern Transcription Factor Irx2, a Target of the FGF8/MAP Kinase Cascade, Is Involved in Cerebellum Formation. Nat. Neurosci. 2004, 7, 605–612. [Google Scholar] [CrossRef]
- Patel, N.S.; Rhinn, M.; Semprich, C.I.; Halley, P.A.; Dollé, P.; Bickmore, W.A.; Storey, K.G. FGF Signalling Regulates Chromatin Organisation during Neural Differentiation via Mechanisms That Can Be Uncoupled from Transcription. PLoS Genet. 2013, 9, e1003614. [Google Scholar] [CrossRef]
- McCracken, K.W.; Aihara, E.; Martin, B.; Crawford, C.M.; Broda, T.; Treguier, J.; Zhang, X.; Shannon, J.M.; Montrose, M.H.; Wells, J.M. Wnt/β-Catenin Promotes Gastric Fundus Specification in Mice and Humans. Nature 2017, 541, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Sakuma, R.; Vakili, N.A.; Mo, R.; Puviindran, V.; Deimling, S.; Zhang, X.; Hopyan, S.; Hui, C. Formation of Proximal and Anterior Limb Skeleton Requires Early Function of Irx3 and Irx5 and Is Negatively Regulated by Shh Signaling. Dev. Cell 2014, 29, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Hernández, M.E.; Bustamante, M.; Galván-Hernández, C.I.; Chimal-Monroy, J. Irx1 and Irx2 Are Coordinately Expressed and Regulated by Retinoic Acid, TGFβ and FGF Signaling during Chick Hindlimb Development. PLoS ONE 2013, 8, e58549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhou, W.; Cai, B.; Chu, J.; Shi, G.; Teng, H.; Xu, J.; Xiao, J.; Wang, Y. IRX2-Mediated Upregulation of MMP-9 and VEGF in a PI3K/AKT-Dependent Manner. Mol. Med. Rep. 2015, 12, 4346–4351. [Google Scholar] [CrossRef] [Green Version]
- Martorell, Ò.; Barriga, F.M.; Merlos-Suárez, A.; Stephan-Otto Attolini, C.; Casanova, J.; Batlle, E.; Sancho, E.; Casali, A. Iro/IRX Transcription Factors Negatively RegulateDPp/TGF-β Pathway Activity during Intestinal Tumorigenesis. EMBO Rep. 2014, 15, 1210–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Song, F.; Sun, H.; Zhang, L.; Huang, C. IRX 5 Promotes NF -ΚB Signalling to Increase Proliferation, Migration and Invasion via OPN in Tongue Squamous Cell Carcinoma. J. Cell. Mol. Med. 2018, 22, 3899–3910. [Google Scholar] [CrossRef]
- Jia, Z.; Zhang, Y.; Yan, A.; Wang, M.; Han, Q.; Wang, K.; Wang, J.; Qiao, C.; Pan, Z.; Chen, C.; et al. 1,25-Dihydroxyvitamin D3 Signaling-Induced Decreases in IRX4 Inhibits NANOG-Mediated Cancer Stem-like Properties and Gefitinib Resistance in NSCLC Cells. Cell Death Dis. 2020, 11, 670. [Google Scholar] [CrossRef]
- Lu, J.; Song, G.; Tang, Q.; Zou, C.; Han, F.; Zhao, Z.; Yong, B.; Yin, J.; Xu, H.; Xie, X.; et al. IRX1 Hypomethylation Promotes Osteosarcoma Metastasis via Induction of CXCL14/NF-ΚB Signaling. J. Clin. Investig. 2015, 125, 1839–1856. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.L.; Karpenko, M.; Lin, M.; Claus, R.; Arab, K.; Dyckhoff, G.; Plinkert, P.; Herpel, E.; Smiraglia, D.; Plass, C. Frequently Methylated Tumor Suppressor Genes in Head and Neck Squamous Cell Carcinoma. Cancer Res. 2008, 68, 4494–4499. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Lee, W.K.; Park, J.Y.; Kim, D.S. Prognostic Value of Iroquois Homeobox 1 Methylation in Non-Small Cell Lung Cancers. Genes Genom. 2020, 42, 571–579. [Google Scholar] [CrossRef]
- Sun, X.; Yi, J.; Yang, J.; Han, Y.; Qian, X.; Liu, Y.; Li, J.; Lu, B.; Zhang, J.; Pan, X.; et al. An Integrated Epigenomic-Transcriptomic Landscape of Lung Cancer Reveals Novel Methylation Driver Genes of Diagnostic and Therapeutic Relevance. Theranostics 2021, 11, 5346–5364. [Google Scholar] [CrossRef]
- He, Y.; Huang, Y.; Li, N.; Yan, H.; Yang, R.F.; Jiang, L.; Jiang, X.H.; Cao, B. Expression of IRX1 in Cervical Cancer and Its Correlation with Clinical Stage of Cervical Cancer. Zhonghua Yi Xue Za Zhi 2018, 98, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, N.; Xu, X.; Wang, Z.; Cheng, Y.; Jin, W.; Wang, X.; Yang, H.; Liu, H.; Zhang, Y.; et al. Clinical Significance of Iroquois Homeobox Gene—IRX1 in Human Glioma. Mol. Med. Rep. 2018, 17, 4651–4656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhou, W.; Zhang, F.; Shi, G.; Teng, H.; Xiao, J.; Wang, Y. Knockdown of IRX2 Inhibits Osteosarcoma Cell Proliferation and Invasion by the AKT/MMP9 Signaling Pathway. Mol. Med. Rep. 2014, 10, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Werner, S.; Stamm, H.; Pandjaitan, M.; Kemming, D.; Brors, B.; Pantel, K.; Wikman, H. Iroquois Homeobox 2 Suppresses Cellular Motility and Chemokine Expression in Breast Cancer Cells. BMC Cancer 2015, 15, 896. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Hussain, W.M.; Vijai, J.; Offit, K.; Rubin, M.A.; Demichelis, F.; Klein, R.J. Variants at IRX4 as Prostate Cancer Expression Quantitative Trait Loci. Eur. J. Hum. Genet. 2013, 22, 558–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha Nguyen, H.; Takata, R.; Akamatsu, S.; Shigemizu, D.; Tsunoda, T.; Furihata, M.; Takahashi, A.; Kubo, M.; Kamatani, N.; Ogawa, O.; et al. IRX4 at 5p15 Suppresses Prostate Cancer Growth through the Interaction with Vitamin D Receptor, Conferring Prostate Cancer Susceptibility. Hum. Mol. Genet. 2012, 21, 2076–2085. [Google Scholar] [CrossRef] [Green Version]
- Chakma, K.; Gu, Z.; Abudurexiti, Y.; Hata, T.; Motoi, F.; Unno, M.; Horii, A.; Fukushige, S. Epigenetic Inactivation of IRX4 Is Responsible for Acceleration of Cell Growth in Human Pancreatic Cancer. Cancer Sci. 2020, 111, 4594–4604. [Google Scholar] [CrossRef]
- Somerville, T.D.D.; Simeoni, F.; Chadwick, J.A.; Williams, E.L.; Spencer, G.J.; Boros, K.; Wirth, C.; Tholouli, E.; Byers, R.J.; Somervaille, T.C.P. Derepression of the Iroquois Homeodomain Transcription Factor Gene IRX3 Confers Differentiation Block in Acute Leukemia. Cell Rep. 2018, 22, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Holmquist Mengelbier, L.; Lindell-Munther, S.; Yasui, H.; Jansson, C.; Esfandyari, J.; Karlsson, J.; Lau, K.; Hui, C.; Bexell, D.; Hopyan, S.; et al. The Iroquois Homeobox Proteins IRX3 and IRX5 Have Distinct Roles in Wilms Tumour Development and Human Nephrogenesis. J. Pathol. 2018, 247, 86–98. [Google Scholar] [CrossRef]
- Zuo, S.; Dai, G.; Ren, X. Identification of a 6-Gene Signature Predicting Prognosis for Colorectal Cancer. Cancer Cell Int. 2019, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, C.; Dai, D.-Q. Identification of Differentially Expressed Genes Regulated by Methylation in Colon Cancer Based on Bioinformatics Analysis. World J. Gastroenterol. 2019, 25, 3392–3407. [Google Scholar] [CrossRef]
- Yuan, C.-L.; Jiang, X.-M.; Yi, Y.; Zhang, N.-D.; Luo, X.; Zou, N.; Wei, W.; Liu, Y.-Y. Identification of Differentially Expressed LncRNAs and MRNAs in Luminal-B Breast Cancer by RNA-Sequencing. BMC Cancer 2019, 19, 1171. [Google Scholar] [CrossRef] [Green Version]
- Palomeras, S.; Diaz-Lagares, Á.; Viñas, G.; Setien, F.; Ferreira, H.J.; Oliveras, G.; Crujeiras, A.B.; Hernández, A.; Lum, D.H.; Welm, A.L.; et al. Epigenetic Silencing of TGFBI Confers Resistance to Trastuzumab in Human Breast Cancer. Breast Cancer Res. 2019, 21, 79. [Google Scholar] [CrossRef] [Green Version]
- Asiedu, M.K.; Thomas, C.F.; Dong, J.; Schulte, S.C.; Khadka, P.; Sun, Z.; Kosari, F.; Jen, J.; Molina, J.; Vasmatzis, G.; et al. Pathways Impacted by Genomic Alterations in Pulmonary Carcinoid Tumors. Clin. Cancer Res. 2018, 24, 1691–1704. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yu, Y.; Zhu, Z.; Xu, H.; Ji, J.; Bu, L.; Liu, B.; Jiang, H.; Lin, Y.; Kong, X.; et al. Identification of a New Target Region by Loss of Heterozygosity at 5p15.33 in Sporadic Gastric Carcinomas: Genotype and Phenotype Related. Cancer Lett. 2005, 224, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ji, J.; Lu, Y.; Bu, L.; Liu, B.; Zhu, Z.; Lin, Y. High-Resolution Analysis of Chromosome 5 and Identification of Candidate Genes in Gastric Cancer. Zhonghua Zhong Liu Za Zhi [Chin. J. Oncol.] 2006, 28, 84–87. [Google Scholar] [PubMed]
- Guo, X.; Liu, W.; Pan, Y.; Ni, P.; Ji, J.; Guo, L.; Zhang, J.; Wu, J.; Jiang, J.; Chen, X.; et al. Homeobox Gene IRX1 Is a Tumor Suppressor Gene in Gastric Carcinoma. Oncogene 2010, 29, 3908–3920. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Li, Z.; Zhao, S.; Yang, B.; Liu, Z. Downregulated MicroRNA-150 Upregulates IRX1 to Depress Proliferation, Migration, and Invasion, but Boost Apoptosis of Gastric Cancer Cells. IUBMB Life 2019, 72, 476–491. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xu, Y.; Hou, P. Identifying Novel Biomarkers of Gastric Cancer through Integration Analysis of Single Nucleotide Polymorphisms and Gene Expression Profile. Int. J. Biol. Markers 2015, 30, 321–326. [Google Scholar] [CrossRef]
- Lim, B. Genomic and Epigenomic Heterogeneity in Molecular Subtypes of Gastric Cancer. World J. Gastroenterol. 2016, 22, 1190. [Google Scholar] [CrossRef]
- Chen, S.; Yu, Y.; Li, T.; Ruan, W.; Wang, J.; Peng, Q.; Yu, Y.; Cao, T.; Xue, W.; Liu, X.; et al. A Novel DNA Methylation Signature Associated with Lymph Node Metastasis Status in Early Gastric Cancer. Clin. Epigenetics 2022, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Liu, W.; Yu, Y.; Ni, P.; Wu, J.; Ji, J.; Zhang, J.; Chen, X.; Lu, B.; Zhu, Z. [Establishment of Experimental Angiogenic Models with Applications of Quantitative Digital Image Analysis]. Zhonghua Bing Li Xue Za Zhi [Chin. J. Pathol.] 2011, 40, 475–479. [Google Scholar]
- Jiang, J.; Liu, W.; Guo, X.; Zhang, R.; Zhi, Q.; Ji, J.; Zhang, J.; Chen, X.; Li, J.; Zhang, J.; et al. IRX1 Influences Peritoneal Spreading and Metastasis via Inhibiting BDKRB2-Dependent Neovascularization on Gastric Cancer. Oncogene 2011, 30, 4498–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamangar, F.; Sheikhattari, P.; Mohebtash, M. Helicobacter Pylori and Its Effects on Human Health and Disease. Arch. Iran. Med. 2011, 14, 192–199. [Google Scholar]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Schistosomes, Liver Flukes and Helicobacter Pylori. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC Press: Lyon, France, 1994; Volume 61, pp. 1–241. [Google Scholar]
- Guo, X.B.; Guo, L.; Zhi, Q.M.; Ji, J.; Jiang, J.L.; Zhang, R.J.; Zhang, J.N.; Zhang, J.; Chen, X.H.; Cai, Q.; et al. Helicobacter Pylori Induces Promoter Hypermethylation and Downregulates Gene Expression of IRX1 Transcription Factor on Human Gastric Mucosa. J. Gastroenterol. Hepatol. 2011, 26, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Q.; Guo, X.; Guo, L.; Zhang, R.; Jiang, J.; Ji, J.; Zhang, J.; Zhang, J.; Chen, X.; Cai, Q.; et al. Oncogenic MiR-544 Is an Important Molecular Target in Gastric Cancer. Anti-Cancer Agents Med. Chem. 2013, 13, 270–275. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.; Liu, L.; Jiang, Y.; Ji, J.; Yan, R.; Zhu, Z.; Yu, Y. Protein Arginine Methyltransferase 5-Mediated Epigenetic Silencing of IRX1 Contributes to Tumorigenicity and Metastasis of Gastric Cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2835–2844. [Google Scholar] [CrossRef]
- Eftang, L.L.; Esbensen, Y.; Tannæs, T.M.; Blom, G.P.; Bukholm, I.R.; Bukholm, G. Up-Regulation of CLDN1 in Gastric Cancer Is Correlated with Reduced Survival. BMC Cancer 2013, 13, 586. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yin, H.; Zheng, S.; Chu, A.; Li, Y.; Xing, C.; Yuan, Y.; Gong, Y. Differentially Expressed MRNAs and Their Long Noncoding RNA Regulatory Network with Helicobacter Pylori-Associated Diseases Including Atrophic Gastritis and Gastric Cancer. BioMed Res. Int. 2020, 2020, 3012193. [Google Scholar] [CrossRef]
- Wang, G.-J.; Huangfu, L.-T.; Gao, X.-Y.; Gan, X.-J.; Xing, X.-F.; Ji, J.-F. A Novel Classification and Scoring Method Based on Immune-Related Transcription Factor Regulation Patterns in Gastric Cancer. Front. Oncol. 2022, 12, 887244. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Xu, Y.; Chen, Y.; Lv, H.; Chen, G.; Chen, Y. Construction of MiRNA-MRNA-TF Regulatory Network for Diagnosis of Gastric Cancer. BioMed Res. Int. 2021, 2021, 9121478. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Pan, S.; Wu, H.; Zhou, J.; Huang, Y.; Wang, S.; Liu, A. ICAM1 Regulates the Development of Gastric Cancer and May Be a Potential Biomarker for the Early Diagnosis and Prognosis of Gastric Cancer. Cancer Manag. Res. 2020, 12, 1523–1534. [Google Scholar] [CrossRef] [Green Version]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H. WHO Classification of Tumours of the Digestive System: WHO Classification of Tumours; World Health Organization: Geneva, Switzerland, 2010; Volume 3.
- Correa, P.; Piazuelo, M.B. The Gastric Precancerous Cascade. J. Dig. Dis. 2011, 13, 2–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vastrad, B.; Vastrad, C. Investigation of the Underlying Hub Genes and Molecular Pathogenesis in Gastric Cancer by Integrated Bioinformatic Analyses. Available online: https://europepmc.org/article/ppr/ppr255988 (accessed on 20 January 2023).
- Qiu, J.; Sun, M.; Wang, Y.; Chen, B. Identification of Hub Genes and Pathways in Gastric Adenocarcinoma Based on Bioinformatics Analysis. Med. Sci. Monit. 2020, 26, e920261-1–e920261-12. [Google Scholar] [CrossRef]
- Zu, F.; Han, H.; Sheng, W.; Sun, J.; Zang, H.; Liang, Y.; Liu, Q. Identification of a Competing Endogenous RNA Axis Related to Gastric Cancer. Aging 2020, 12, 20540–20560. [Google Scholar] [CrossRef]
- Ma, Z.; Xu, J.; Ru, L.; Zhu, W. Identification of Pivotal Genes Associated with the Prognosis of Gastric Carcinoma through Integrated Analysis. Biosci. Rep. 2021, 41, BSR20203676. [Google Scholar] [CrossRef]
- Jing, J.; Wang, Z.; Li, H.; Sun, L.; Yuan, Y. Key Elements Involved in Epstein–Barr Virus-Associated Gastric Cancer and Their Network Regulation. Cancer Cell Int. 2018, 18, 146. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.G. Somatostatin and Somatostatin Analogues: Pharmacokinetics and Pharmacodynamic Effects. Gut 1994, 35, S1–S4. [Google Scholar] [CrossRef] [Green Version]
- Margheri, F.; Schiavone, N.; Papucci, L.; Magnelli, L.; Serratì, S.; Chillà, A.; Laurenzana, A.; Bianchini, F.; Calorini, L.; Torre, E.; et al. GDF5 Regulates TGFß-Dependent Angiogenesis in Breast Carcinoma MCF-7 Cells: In Vitro and in Vivo Control by Anti-TGFß Peptides. PLoS ONE 2012, 7, e50342. [Google Scholar] [CrossRef] [Green Version]
- Enescu, A.S.; Mărgăritescu, C.L.; Crăiţoiu, M.M.; Enescu, A.; Crăiţoiu, Ş. The Involvement of Growth Differentiation Factor 5 (GDF5) and Aggrecan in the Epithelial-Mesenchymal Transition of Salivary Gland Pleomorphic Adenoma. Rom. J. Morphol. Embryol. Rev. Roum. De Morphol. Embryol. 2013, 54, 969–976. [Google Scholar]
- Li, H.; Liu, J.; Liu, S.; Yuan, Y.; Sun, L. Bioinformatics-Based Identification of Methylated-Differentially Expressed Genes and Related Pathways in Gastric Cancer. Dig. Dis. Sci. 2017, 62, 3029–3039. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-Arévalo, S.; Hormaechea-Agulla, D.; Gómez-Gómez, E.; Requena, M.J.; Selth, L.A.; Gahete, M.D.; Castaño, J.P.; Luque, R.M. Somatostatin Receptor Subtype 1 as a Potential Diagnostic Marker and Therapeutic Target in Prostate Cancer. Prostate 2017, 77, 1499–1511. [Google Scholar] [CrossRef] [PubMed]
- Znalesniak, E.B.; Salm, F.; Hoffmann, W. Molecular Alterations in the Stomach of Tff1-Deficient Mice: Early Steps in Antral Carcinogenesis. Int. J. Mol. Sci. 2020, 21, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Feng, Y.; Sun, Z.; Gao, Y.; Wu, C.; Zhang, H.; Cao, J.; Chen, Z.; Cao, J.; Zhu, Y.; et al. mRNA and lncRNA Expression Profiling of Radiation-Induced Gastric Injury Reveals Potential Radiation-Responsive Transcription Factors. Dose-Response 2019, 17, 155932581988676. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Zhang, C.; Ai, S.; Liu, Z.; Lu, X. Identification of Hub Genes in Gastric Cancer by Integrated Bioinformatics Analysis. Transl. Cancer Res. 2021, 10, 2831–2840. [Google Scholar] [CrossRef]
- Tian, Y.; Xing, Y.; Zhang, Z.; Peng, R.; Zhang, L.; Sun, Y. Bioinformatics Analysis of Key Genes and CircRNA-MiRNA-MRNA Regulatory Network in Gastric Cancer. BioMed Res. Int. 2020, 2020, 2862701. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ding, Z.; Zhuang, Q.; Shen, L.; Yang, F.; Sah, S.; Wu, G. Analysis of Different Adipose Depot Gene Expression in Cachectic Patients with Gastric Cancer. Nutr. Metab. 2022, 19, 72. [Google Scholar] [CrossRef]
- Miao, R.; Guo, X.; Zhi, Q.; Shi, Y.; Li, L.; Mao, X.; Zhang, L.; Li, C. VEZT, a Novel Putative Tumor Suppressor, Suppresses the Growth and Tumorigenicity of Gastric Cancer. PLoS ONE 2013, 8, e74409. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Ishikawa, T.; Ichiishi, E.; Yoshida, Y.; Hanashiro, K.; Kuchide, M.; Uchiyama, K.; Kokura, S.; Ichikawa, H.; Naito, Y.; et al. The Effect of Rebamipide on Helicobacter Pylori Extract-Mediated Changes of Gene Expression in Gastric Epithelial Cells. Aliment. Pharmacol. Ther. 2003, 18 (Suppl. S1), 63–75. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Research Network. Comprehensive Molecular Characterization of Gastric Adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Bartha, Á.; Győrffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue of Somatic Mutations in Cancer. Nucleic Acids Res. 2018, 47, D941–D947. [Google Scholar] [CrossRef] [Green Version]
- Shihab, H.A.; Gough, J.; Cooper, D.N.; Stenson, P.D.; Barker, G.L.A.; Edwards, K.J.; Day, I.N.M.; Gaunt, T.R. Predicting the Functional, Molecular, and Phenotypic Consequences of Amino Acid Substitutions Using Hidden Markov Models. Hum. Mutat. 2012, 34, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IRX1 | ||||||
| Regulation | Methodology | Metilation Status | Tissue | Histology | Sample | Citation |
| Down | RT–qPCR | Metilated | GC | Intestinal | NCI-N87 Cell line | [79,80,89,90] |
| Down | RT–qPCR | Metilated | GC | Poorly differentiated | SNU-1 Cell line | |
| carcinoma | ||||||
| Down | RT–qPCR | Metilated | GC | Diffuse | SNU-16 Cell line | |
| Down | RT–qPCR | Metilated | GC | Intestinal | AGS Cell line | |
| Down | RT–qPCR | Metilated | GC | Diffuse | KATO-III Cell line | |
| Down | RT–qPCR | Metilated | GC | Diffuse | MKN-45 Cell line | |
| Down | RT–qPCR | Metilated | GC | Intestinal | SGC-7901 Cell line | |
| Control non cancer | RT–qPCR | Unmetilated | Epitelial | Normal | GES-1 (control cell) | |
| Control non cancer | Western blot/RT–qPCR | Epitelial | Normal | RGM-1 Cell line | [80] | |
| Down | Western blot/RT–qPCR | GC | Intestinal | BGC823 Cell line | [90] | |
| Down | Western blot/RT–qPCR | GC | Intestinal | MKN28 Cell line | ||
| Down | Western blot/RT–qPCR | GC | Diffuse | Hs746T Cell line | ||
| Down | Western blot/RT–qPCR | GC | Poorly differentiated | SUN-1 Cell line | ||
| carcinoma | ||||||
| Down | Western blot/RT–qPCR | CG | 36 patients cancer and adjacent | |||
| Down | Western blot/RT–qPCR | GC | 60 patients cancer and adjacent | [80] | ||
| genome-wide methylation sequencing | Hypermetilated | GC | GC | 23 lymph node metastasis positive (LNM+) and 24 lymph node metastasis negative (LNM−) | [83] | |
| IRX2 | ||||||
| Regulation | Methodology | Metilation status | Tissue | Histology | Sample | Citation |
| Down | cDNA microarrays | GC | Diffuse/intestinal/mixed | 20 patients cancer and adjacent | [91] | |
| Up | Gene expression array | EGC, LGIN | 58 patients | [95] | ||
| Down | Microarray | GC and normal | 210 gastric cancer tissues and 118 normal tissue | [94] | ||
| Down | LncRNA/mRNA analysis set | GC, normal mucosa and atrophic gastritis | 13 | [92] | ||
| Down | Microarray | GC | Diffuse | 107 diffuse type gastric cancer tissues samples and 10 normal gastric tissues samples | [98] | |
| Down | Microarray | GC | GC | 210 gastric cancer tissues 118 normal tissue samples | [94] | |
| Genome-wide methylation sequencing | Hypometilated | GC | GC | 23 lymph node metastasis positive (LNM+) and 24 lymph node metastasis negative (LNM−) | [83] | |
| IRX3 | ||||||
| Regulation | Methodology | Metilation status | Tissue | Histology | Sample | Citation |
| Down | cDNA microarrays | GC | Diffuse/intestinal/mixed | 20 patients cancer and adjacent | [91] | |
| Down | Microarray | GC | GC | 70 gastric adenocarcinoma samples and 68 matched normal samples | [99] | |
| Down | Microarray | GC | GC | 663 cancer samples and 223 normal cases | [100] | |
| Down | Microarray | GC | Diffuse | 107 diffuse type gastric cancer tissues samples and 10 normal gastric tissues samples | [98] | |
| Down | Microarray | GC | GC | 210 gastric cancer tissues 118 normal tissue samples | [94] | |
| IRX4 | ||||||
| Regulation | Methodology | Metilation status | Tissue | Histology | Sample | Citation |
| Down | microarray | Stomach | mice | [109] | ||
| Down | RNA-seq | GC | GC | 375 GC primary tumor samples and 32 solid normal tissue s | [110] | |
| IRX5 | ||||||
| Regulation | Methodology | Metilation status | Tissue | Histology | Sample | Citation |
| Down | Microarray | GC | GC | 63 GC tissues and 56 paracancer tissues | [111] | |
| Down | microarray | SAT and VAT | GC | 3 | [112] | |
| Down | microarray | GC | Diffuse | 107 diffuse type gastric cancer tissues samples and 10 normal gastric tissues samples | [98] | |
| IRX6 | ||||||
| Regulation | Methodology | Metilation status | Tissue | Histology | Sample | Citation |
| Genome-wide methylation sequencing | Hypometilated | GC | GC | 23 lymph node metastasis positive (LNM+) and 24 lymph node metastasis negative (LNM−) | [83] | |
| Transcript Name (Translation ID) | Homeobox Domain | Iroquois-Class Homeodomain Protein | Possible Pathogenic Mutation ** | Samples Tested | Mutated Samples | Homeobox Domain Possible Pathogenic Mutation ** | Iroquois-Class Homeodomain Protein Possible Pathogenic Mutation ** | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | Exon | Start | End | Exon | Mutation (Amino Acid) | |||||
| IRX1-201 (ENST00000302006.4) | 128 | 192 | 2 | 309 | 326 | 2 | p.A71V; p.A114V; p.K132T; p.Y154C; p.A179T; p.A181T; p.A181V; p.R182C; p.R196C; p.I223V; p.E290V; p.S377=; p.R389H; p.V394I; p.S433R; p.V442D; p.D461N; p.N462T; p.G469E; p.R472W | 63 | 19 | p.K132T; p.Y154C; p.A179T; p.A181T; p.A181V; p.R182C | - |
| IRX2-201 (ENST00000302057.6) | 114 | 179 | 2 | 322 | 339 | 3 | p.N113K; p.A116T; p.K130R; p.Y141=; p.T157A; p.R171H; p.D217=; p.E235 *; p.C250G; p.S254L; p.D265A; p.E274 *; p.S386P; p.N399=; p.Y410C; p.E458K; p.V463I | 50 | 16 | p.A116T; p.K130R; p.Y141=; p.T157A; p.R171H | - |
| IRX2-202 (ENST00000382611.10) | 114 | 179 | 2 | 322 | 339 | 3 | |||||
| IRX3-201 (ENST00000329734.4) | 128 | 192 | 2 | 342 | 359 | 2 | p.Y73=; p.G158=; p.R184L; p.R184=; p.G203E; p.P336S; p.R463H; p.V481= | 33 | 8 | p.G158=; p.R184L; p.R184= | - |
| IRX4-201 (ENST00000231357.7) | 142 | 207 | 4 | 361 | 378 | 5 | p.A35V; p.T88A; p.P124S; p.S168I; p.T170M; p.R177H; p.I207V; p.R225H; p.T233M; p.A271=; p.R348M; p.S439A; p.P457L; p.S499L; p.A502V | 41 | 14 | p.S168I; p.T170M; p.R177H; p.I207V; p.R225H; p.T233M | - |
| IRX4-202 (ENST00000505790.5) | 142 | 207 | 4 | 361 | 378 | 5 | |||||
| IRX4-206 (ENST00000513692.5) | 142 | 207 | 4 | 361 | 378 | 5 | |||||
| IRX4-207 (ENST00000613726.4) | 168 | 233 | 5 | 387 | 404 | 6 | |||||
| IRX4-208 (ENST00000622814.4) | 168 | 233 | 5 | 387 | 404 | 6 | |||||
| IRX5-201 (ENST00000320990.9) | 113 | 178 | 2 | 326 | 343 | 3 | p.P114T; p.T156=; p.L157I; p.R182Q; p.E188G; p.E201K; p.D212N; p.A341V; p.A452T; p.P457=; p.? | 25 | 11 | p.P114T; p.T156=; p.L157I | p.A341V |
| IRX5-202 (ENST00000394636.9) | 113 | 178 | 2 | 326 | 344 | 3 | |||||
| IRX6-201 (ENST00000290552.8) | 146 | 211 | 4 | 344 | 361 | 5 | p.V34I; p.A44V; p.R55Q; p.L56=; p.S145C; p.P349L; p.E432K | 33 | 7 | - | p.P349L |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, E.C.; Petrone, I.; Binato, R.; Abdelhay, E. Iroquois Family Genes in Gastric Carcinogenesis: A Comprehensive Review. Genes 2023, 14, 621. https://doi.org/10.3390/genes14030621
dos Santos EC, Petrone I, Binato R, Abdelhay E. Iroquois Family Genes in Gastric Carcinogenesis: A Comprehensive Review. Genes. 2023; 14(3):621. https://doi.org/10.3390/genes14030621
Chicago/Turabian Styledos Santos, Everton Cruz, Igor Petrone, Renata Binato, and Eliana Abdelhay. 2023. "Iroquois Family Genes in Gastric Carcinogenesis: A Comprehensive Review" Genes 14, no. 3: 621. https://doi.org/10.3390/genes14030621