
Genome-Wide Identification and Characterization of the PPO Gene Family in Cotton (Gossypium) and Their Expression Variations Responding to Verticillium Wilt Infection

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of PPO Genes
2.2. Phylogenetic Analysis and Classification of PPO Genes
2.3. Analysis of PPO Gene Structure and Protein Domain in Cotton
2.4. Expression Pattern Analysis of PPO Genes in Cotton
2.5. RNA Extraction and Gene Expression Analysis by Real-Time Quantitative PCR
3. Results
3.1. Identification and Characteristics of PPO Genes in Gossypium
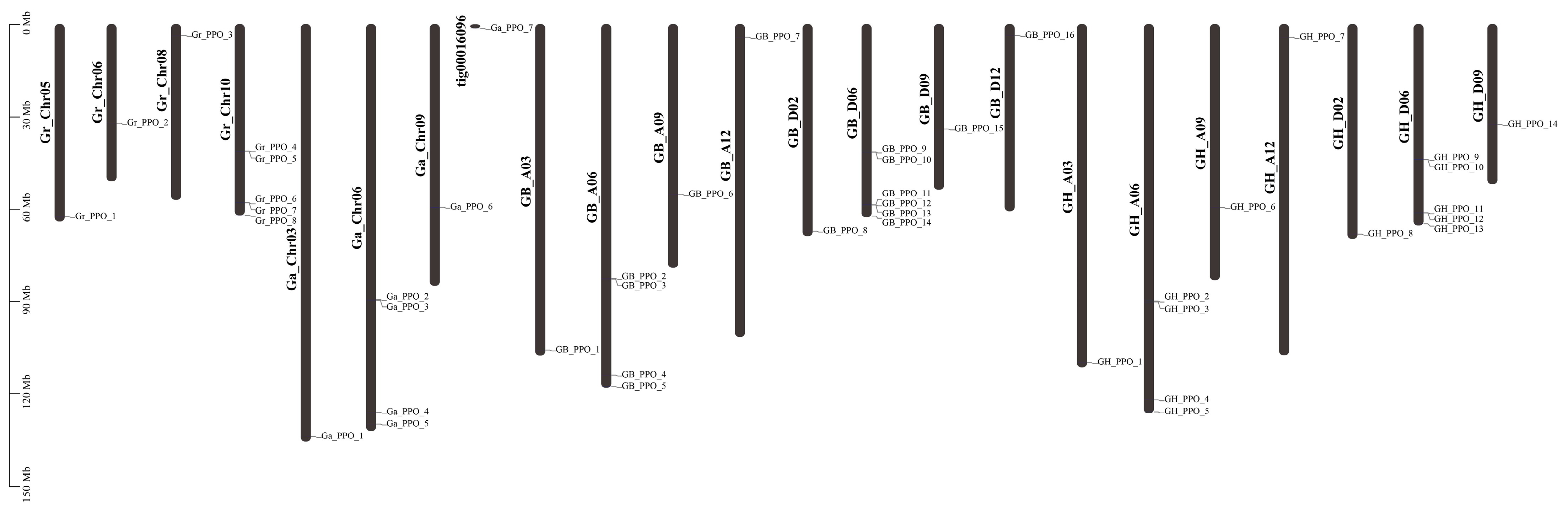
3.2. Chromosome Distribution of PPO Gene Family in Cotton
3.3. Structure Analysis of PPO Gene Family
3.4. Analyses of Gene Structure and Protein Domain of Cotton PPOs
3.5. Expression Analysis of PPO Gene Family in Diploid Cotton
3.6. Expression Investigation of GhPPO Genes in Response to VW Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, S.M.; Eannetta, N.T.; Yu, H.; Prince, J.P.; de Vicente, M.C.; Tanksley, S.D.; Steffens, J.C. Organisation of the tomato polyphenol oxidase gene family. Plant Mol. Biol. 1993, 21, 1035–1051. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, X. Recent advances in polyphenol oxidase-mediated plant stress responses. Phytochemistry 2021, 181, 112588. [Google Scholar] [CrossRef] [PubMed]
- Vámos-Vigyázó, L. Polyphenol oxidase and peroxidase in fruits and vegetables. Crit. Rev. Food Sci. Nutr. 1981, 15, 49–127. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef]
- Thipyapong, P.; Hunt, M.D.; Steffens, J.C. Antisense downregulation of polyphenol oxidase results in enhanced disease susceptibility. Planta 2004, 220, 105–117. [Google Scholar] [CrossRef]
- Parveen, I.; Threadgill, M.D.; Moorby, J.M.; Winters, A. Oxidative phenols in forage crops containing polyphenol oxidase enzymes. J. Agric. Food Chem. 2010, 58, 1371–1382. [Google Scholar] [CrossRef]
- Massa, A.N.; Beecher, B.; Morris, C.F. Polyphenol oxidase (PPO) in wheat and wild relatives: Molecular evidence for a multigene family. Theor. Appl. Genet. 2007, 114, 1239–1247. [Google Scholar] [CrossRef]
- Winters, A.; Heywood, S.; Farrar, K.; Donnison, I.; Thomas, A.; Webb, K.J. Identification of an extensive gene cluster among a family of PPOs in Trifolium pratense L. (red clover) using a large insert BAC library. BMC Plant Biol. 2009, 9, 94. [Google Scholar] [CrossRef]
- Li, C.; Li, D.; Li, J.; Shao, F.; Lu, S. Characterization of the polyphenol oxidase gene family reveals a novel microRNA involved in posttranscriptional regulation of PPOs in Salvia miltiorrhiza. Sci. Rep. 2017, 7, 44622. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, J.; Cui, D.; Zhao, D.; Benedito, V.A.; Zhao, J. Genome-wide analysis and metabolic profiling unveil the role of peroxidase CsGPX3 in theaflavin production in black tea processing. Food Res. Int. 2020, 137, 109677. [Google Scholar] [CrossRef]
- He, F.; Shi, Y.J.; Zhao, Q.; Zhao, K.J.; Cui, X.L.; Chen, L.H.; Yang, H.B.; Zhang, F.; Mi, J.X.; Huang, J.L.; et al. Genome-wide investigation and expression profiling of polyphenol oxidase (PPO) family genes uncover likely functions in organ development and stress responses in Populus trichocarpa. BMC Genom. 2021, 22, 731. [Google Scholar] [CrossRef]
- Desai, A.; Chee, P.W.; Rong, J.; May, O.L.; Paterson, A.H. Chromosome structural changes in diploid and tetraploid A genomes of Gossypium. Genome 2006, 49, 336–345. [Google Scholar] [CrossRef]
- Li, P.T.; Wang, M.; Lu, Q.W.; Ge, Q.; Rashid, M.H.O.; Liu, A.Y.; Gong, J.W.; Shang, H.H.; Gong, W.K.; Li, J.W.; et al. Comparative transcriptome analysis of cotton fiber development of Upland cotton (Gossypium hirsutum) and Chromosome Segment Substitution Lines from G. hirsutum × G. barbadense. BMC Genom. 2017, 18, 705. [Google Scholar] [CrossRef]
- Ma, A.; Zhang, D.; Wang, G.; Wang, K.; Li, Z.; Gao, Y.; Li, H.; Bian, C.; Cheng, J.; Han, Y.; et al. Verticillium dahliae effector VDAL protects MYB6 from degradation by interacting with PUB25 and PUB26 E3 ligases to enhance Verticillium wilt resistance. Plant Cell 2021, 33, 3675–3699. [Google Scholar] [CrossRef]
- Tong, S.; Yuan, M.; Liu, Y.; Li, X.; Jin, D.; Cheng, X.; Lin, D.; Ling, H.; Yang, D.; Wang, Y.; et al. Ergosterol-targeting fusion antifungal peptide significantly increases the Verticillium wilt resistance of cotton. Plant Biotechnol. J. 2021, 19, 926–936. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Ding, C.; Xu, W.; Wang, X. Temporal patterns of cotton Fusarium and Verticillium wilt in Jiangsu coastal areas of China. Sci. Rep. 2017, 7, 12581. [Google Scholar] [CrossRef]
- Xi, H.; Zhang, X.; Qu, Z.; Yang, D.; Alariqi, M.; Yang, Z.; Nie, X.; Zhu, L. Effects of cotton-maize rotation on soil microbiome structure. Mol. Plant Pathol. 2021, 22, 673–682. [Google Scholar] [CrossRef]
- Castro, D.; Torres, M.; Sampedro, I.; Martínez-Checa, F.; Torres, B.; Béjar, V. Biological control of Verticillium wilt on olive trees by the salt-tolerant strain Bacillus velezensis XT1. Microorganisms 2020, 8, 1080. [Google Scholar] [CrossRef]
- Berne, S.; Kovačević, N.; Kastelec, D.; Javornik, B.; Radišek, S. Hop Polyphenols in Relation to Verticillium wilt resistance and their antifungal activity. Plants 2020, 9, 1318. [Google Scholar] [CrossRef]
- Du, X.; Huang, G.; He, S.; Yang, Z.; Sun, G.; Ma, X.; Li, N.; Zhang, X.; Sun, J.; Liu, M.; et al. Resequencing of 243 diploid cotton accessions based on an updated A genome identifies the genetic basis of key agronomic traits. Nat. Genet. 2018, 50, 796–802. [Google Scholar] [CrossRef]
- Paterson, A.H.; Wendel, J.F.; Gundlach, H.; Guo, H.; Jenkins, J.; Jin, D.; Llewellyn, D.; Showmaker, K.C.; Shu, S.; Udall, J.; et al. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres. Nature 2012, 492, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, J.; Fang, L.; Zhang, Z.; Ma, W.; Niu, Y.; Ju, L.; Deng, J.; Zhao, T.; Lian, J.; et al. Gossypium barbadense and Gossypium hirsutum genomes provide insights into the origin and evolution of allotetraploid cotton. Nat. Genet. 2019, 51, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y.; Lee, S.; Van, K.; Kim, T.H.; Jeong, S.C.; Choi, I.Y.; Kim, D.S.; Lee, Y.S.; Park, D.; Ma, J.; et al. Whole-genome sequencing and intensive analysis of the undomesticated soybean (Glycine soja Sieb. and Zucc.) genome. Proc. Natl. Acad. Sci. USA 2010, 107, 22032–22037. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhou, Y.; Chen, J.; Shi, J.; Zhao, H.; Zhao, H.; Song, W.; Zhang, M.; Cui, Y.; Dong, X.; et al. Extensive intraspecific gene order and gene structural variations between Mo17 and other maize genomes. Nat. Genet. 2018, 50, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Branca, A.; Paape, T.D.; Zhou, P.; Briskine, R.; Farmer, A.D.; Mudge, J.; Bharti, A.K.; Woodward, J.E.; May, G.D.; Gentzbittel, L.; et al. Whole-genome nucleotide diversity, recombination, and linkage disequilibrium in the model legume Medicago truncatula. Proc. Natl. Acad. Sci. USA 2011, 108, E864–E870. [Google Scholar] [CrossRef]
- Yu, J.; Hu, S.; Wang, J.; Wong, G.K.; Li, S.; Liu, B.; Deng, Y.; Dai, L.; Zhou, Y.; Zhang, X.; et al. A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 2002, 296, 79–92. [Google Scholar] [CrossRef]
- Argout, X.; Salse, J.; Aury, J.M.; Guiltinan, M.J.; Droc, G.; Gouzy, J.; Allegre, M.; Chaparro, C.; Legavre, T.; Maximova, S.N.; et al. The genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar] [CrossRef]
- Tuskan, G.A.; Difazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 6: Recent updates and new developments. Nucleic Acids Res. 2009, 37, D229–D232. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Adobe Creative Team. Adobe Illustrator CS6 Classroom in a Book; Adobe Press: San Jose, CA, USA, 2012. [Google Scholar]
- Guo, A.Y.; Zhu, Q.H.; Chen, X.; Luo, J.C. GSDS: A gene structure display server. Yi Chuan 2007, 29, 1023–1026. (In Chinese) [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Huang, G.; Wu, Z.; Percy, R.G.; Bai, M.; Li, Y.; Frelichowski, J.E.; Hu, J.; Wang, K.; Yu, J.Z.; Zhu, Y. Genome sequence of Gossypium herbaceum and genome updates of Gossypium arboreum and Gossypium hirsutum provide insights into cotton A-genome evolution. Nat. Genet. 2020, 52, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Kolde, R. Pheatmap: Pretty Heatmaps, version 3.4+, R Package: San Francisco, CA, USA, 2015; volume 1.
- Li, P.T.; Rashid, M.H.O.; Chen, T.T.; Lu, Q.W.; Ge, Q.; Gong, W.K.; Liu, A.Y.; Gong, J.W.; Shang, H.H.; Deng, X.Y.; et al. Transcriptomic and biochemical analysis of upland cotton (Gossypium hirsutum) and a chromosome segment substitution line from G. hirsutum × G. barbadense in response to Verticillium dahliae infection. BMC Plant Biol. 2019, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Jiang, Y.; Cao, Y.; Lin, L.; He, S.; Zhou, W.; Rong, J. Gene expression profile analysis of Ligon lintless-1 (Li1) mutant reveals important genes and pathways in cotton leaf and fiber development. Gene 2014, 535, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shen, Y.; Li, Z.; Xie, X.; Gong, E.S.; Tian, J.; Si, X.; Wang, Y.; Gao, N.; Shu, C.; et al. Effects of high hydrostatic pressure and thermal processing on anthocyanin content, polyphenol oxidase and β-glucosidase activities, color, and antioxidant activities of blueberry (Vaccinium spp.) puree. Food Chem. 2021, 342, 128564. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of Silicon on Plant-Pathogen Interactions. Front Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef] [PubMed]
- Aziz, E.; Batool, R.; Akhtar, W.; Rehman, S.; Gregersen, P.L.; Mahmood, T. Expression analysis of the polyphenol oxidase gene in response to signaling molecules, herbivory and wounding in antisense transgenic tobacco plants. 3 Biotech 2019, 9, 55. [Google Scholar] [CrossRef]
- Chi, M.; Bhagwat, B.; Lane, W.D.; Tang, G.; Su, Y.; Sun, R.; Oomah, B.D.; Wiersma, P.A.; Xiang, Y. Reduced polyphenol oxidase gene expression and enzymatic browning in potato (Solanum tuberosum L.) with artificial microRNAs. BMC Plant Biol. 2014, 14, 62. [Google Scholar] [CrossRef]
- Thipyapong, P.; Joel, D.M.; Steffens, J.C. Differential Expression and Turnover of the Tomato Polyphenol Oxidase Gene Family during Vegetative and Reproductive Development. Plant Physiol. 1997, 113, 707–718. [Google Scholar] [CrossRef]
- Tran, L.T.; Constabel, C.P. The polyphenol oxidase gene family in poplar: Phylogeny, differential expression and identification of a novel, vacuolar isoform. Planta 2011, 234, 799–813. [Google Scholar] [CrossRef]
- Wendel, J.F.; Cronn, R.C. Polyploidy and the evolutionary history of cotton. Adv. Agron. 2003, 78, 139–186. [Google Scholar]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, D.; Hu, H.; Li, W.; Hu, Y.; Xie, J.; Huang, S.; Wang, W. Genome-wide characterization of a SRO gene family involved in response to biotic and abiotic stresses in banana (Musa spp.). BMC Plant Biol. 2019, 19, 211. [Google Scholar] [CrossRef]
- Narasimhan, M.L.; Damsz, B.; Coca, M.A.; Ibeas, J.I.; Yun, D.J.; Pardo, J.M.; Hasegawa, P.M.; Bressan, R.A. A plant defense response effector induces microbial apoptosis. Mol. Cell 2001, 8, 921–930. [Google Scholar] [CrossRef]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161. [Google Scholar] [CrossRef]
- Wang, J.; Constabel, C.P. Polyphenol oxidase overexpression in transgenic Populus enhances resistance to herbivory by forest tent caterpillar (Malacosoma disstria). Planta 2004, 220, 87–96. [Google Scholar] [CrossRef]
- Liu, D.; Meng, S.; Xiang, Z.; Yang, G.; He, N. An R1R2R3 MYB Transcription Factor, MnMYB3R1, Regulates the Polyphenol Oxidase Gene in Mulberry (Morus notabilis). Int. J. Mol. Sci. 2019, 20, 2602. [Google Scholar] [CrossRef]
- Thipyapong, P.; Steffens, J.C. Tomato Polyphenol Oxidase (Differential Response of the Polyphenol Oxidase F Promoter to Injuries and Wound Signals). Plant Physiol. 1997, 115, 409–418. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomics Position | Protein Length (aa) | Molecular Weight (kDa) | Isoelectric Point |
|---|---|---|---|---|---|
| Ga_PPO_1 [41] | Ga03G2532 | Chr03:133861218–133862947 | 548 | 63.195 | 7.086 |
| Ga_PPO_2 [41] | Ga06G1498 | Chr06:89406195–89407991 | 598 | 67.138 | 6.79 |
| Ga_PPO_3 [41] | Ga06G1499 | Chr06:89662332–89663647 | 413 | 47.373 | 5.922 |
| Ga_PPO_4 [41] | Ga06G2142 | Chr06:125985836–125987683 | 615 | 70.181 | 7.062 |
| Ga_PPO_5 [41] | Ga06G2393 | Chr06:129881213–129882703 | 496 | 56.889 | 6.209 |
| Ga_PPO_6 [41] | Ga09G0868 | Chr09:59384931–59386688 | 585 | 65.256 | 7.612 |
| Ga_PPO_7 [41] | Ga14G1905 | tig00016096:1234774–1236648 | 537 | 61.392 | 7.48 |
| Gr_PPO_1 [21] | Gorai.005G244400 | Chr05:62477068–62479105 | 548 | 63.326 | 6.866 |
| Gr_PPO_2 [21] | Gorai.006G086400 | Chr06:32091285–32093143 | 584 | 65.301 | 7.215 |
| Gr_PPO_3 [21] | Gorai.008G029600 | Chr08:3520590–3522462 | 561 | 64.067 | 7.818 |
| Gr_PPO_4 [21] | Gorai.010G151800 | Chr10:41097418–41099771 | 598 | 67.192 | 6.652 |
| Gr_PPO_5 [21] | Gorai.010G152000 | Chr10:41182365–41184434 | 599 | 67.596 | 6.879 |
| Gr_PPO_6 [21] | Gorai.010G213100 | Chr10:57917180–57919392 | 619 | 70.222 | 6.986 |
| Gr_PPO_7 [21] | Gorai.010G213300 | Chr10:57928797–57930609 | 568 | 63.512 | 5.472 |
| Gr_PPO_8 [21] | Gorai.010G254900 | Chr10:62004848–62006551 | 567 | 64.768 | 6.617 |
| GH_PPO_1 [22] | GH_A03G2196 | A03:109830718–109832447 | 548 | 63.247 | 7.266 |
| GH_PPO_2 [22] | GH_A06G1441 | A06:89879934–89881730 | 598 | 67.18 | 6.79 |
| GH_PPO_3 [22] | GH_A06G1442 | A06:90147702–90149552 | 616 | 69.307 | 7.186 |
| GH_PPO_4 [22] | GH_A06G2011 | A06:121988167–121990017 | 616 | 70.378 | 7.235 |
| GH_PPO_5 [22] | GH_A06G2315 | A06:125891271–125892974 | 567 | 64.513 | 7.05 |
| GH_PPO_6 [22] | GH_A09G0900 | A09:59466573–59468330 | 585 | 65.175 | 7.827 |
| GH_PPO_7 [22] | GH_A12G0274 | A12:4105742–4109367 | 593 | 67.96 | 7.523 |
| GH_PPO_8 [22] | GH_D02G2367 | D02:68093821–68095562 | 548 | 63.293 | 7.086 |
| GH_PPO_9 [22] | GH_D06G1461 | D06:43843765–43845561 | 598 | 67.166 | 6.519 |
| GH_PPO_10 [22] | GH_D06G1462 | D06:43930778–43932628 | 616 | 69.617 | 7.186 |
| GH_PPO_11 [22] | GH_D06G2044 | D06:61154513–61156396 | 627 | 71.531 | 6.51 |
| GH_PPO_12 [22] | GH_D06G2047 | D06:61198543–61200323 | 555 | 63.156 | 6.329 |
| GH_PPO_13 [22] | GH_D06G2351 | D06:64782818–64784521 | 567 | 64.815 | 6.719 |
| GH_PPO_14 [22] | GH_D09G0860 | D09:32552402–32554156 | 584 | 65.439 | 6.917 |
| GB_PPO_1 [22] | GB_A03G2281 | A03:105823405–105825134 | 548 | 63.225 | 7.081 |
| GB_PPO_2 [22] | GB_A06G1484 | A06:82535557–82536873 | 438 | 50.192 | 5.488 |
| GB_PPO_3 [22] | GB_A06G1485 | A06:82746888–82748738 | 616 | 69.378 | 7.022 |
| GB_PPO_4 [22] | GB_A06G2053 | A06:113959340–113961190 | 616 | 70.378 | 7.235 |
| GB_PPO_5 [22] | GB_A06G2345 | A06:117626828–117628531 | 567 | 64.513 | 7.05 |
| GB_PPO_6 [22] | GB_A09G1011 | A09:55231074–55232831 | 585 | 65.237 | 7.827 |
| GB_PPO_7 [22] | GB_A12G0273 | A12:4077941–4079816 | 571 | 65.345 | 7.224 |
| GB_PPO_8 [22] | GB_D02G2425 | D02:67144871–67146608 | 548 | 63.278 | 6.831 |
| GB_PPO_9 [22] | GB_D06G1511 | D06:41409523–41410830 | 435 | 49.822 | 5.647 |
| GB_PPO_10 [22] | GB_D06G1512 | D06:41500819–41502134 | 417 | 47.666 | 5.79 |
| GB_PPO_11 [22] | GB_D06G2136 | D06:58569149–58571008 | 619 | 70.365 | 7.235 |
| GB_PPO_12 [22] | GB_D06G2138 | D06:58617125–58618063 | 312 | 35.385 | 5.289 |
| GB_PPO_13 [22] | GB_D06G2142 | D06:58653169–58660371 | 609 | 69.141 | 7.156 |
| GB_PPO_14 [22] | GB_D06G2444 | D06:62265335–62272753 | 758 | 86.703 | 5.424 |
| GB_PPO_15 [22] | GB_D09G0864 | D09:33963391–33965145 | 584 | 65.334 | 7.242 |
| GB_PPO_16 [22] | GB_D12G0284 | D12:3557843–3559716 | 571 | 65.353 | 7.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Ge, Q.; Wan, S.; Sun, Z.; Chen, Y.; Li, Y.; Liu, Q.; Gong, J.; Xiao, X.; Lu, Q.; et al. Genome-Wide Identification and Characterization of the PPO Gene Family in Cotton (Gossypium) and Their Expression Variations Responding to Verticillium Wilt Infection. Genes 2023, 14, 477. https://doi.org/10.3390/genes14020477
Yang S, Ge Q, Wan S, Sun Z, Chen Y, Li Y, Liu Q, Gong J, Xiao X, Lu Q, et al. Genome-Wide Identification and Characterization of the PPO Gene Family in Cotton (Gossypium) and Their Expression Variations Responding to Verticillium Wilt Infection. Genes. 2023; 14(2):477. https://doi.org/10.3390/genes14020477
Chicago/Turabian StyleYang, Shuhan, Qun Ge, Sumei Wan, Zhihao Sun, Yu Chen, Yanfang Li, Qiankun Liu, Juwu Gong, Xianghui Xiao, Quanwei Lu, and et al. 2023. "Genome-Wide Identification and Characterization of the PPO Gene Family in Cotton (Gossypium) and Their Expression Variations Responding to Verticillium Wilt Infection" Genes 14, no. 2: 477. https://doi.org/10.3390/genes14020477