
Role of miR-143 and miR-146 in Risk Evaluation of Coronary Artery Diseases in Autopsied Samples

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. SNP Genotyping
2.3. RNA Isolation from the Plasma
2.4. Reverse Transcription
2.5. miRNA Expression
2.6. Statistical Analysis
3. Results
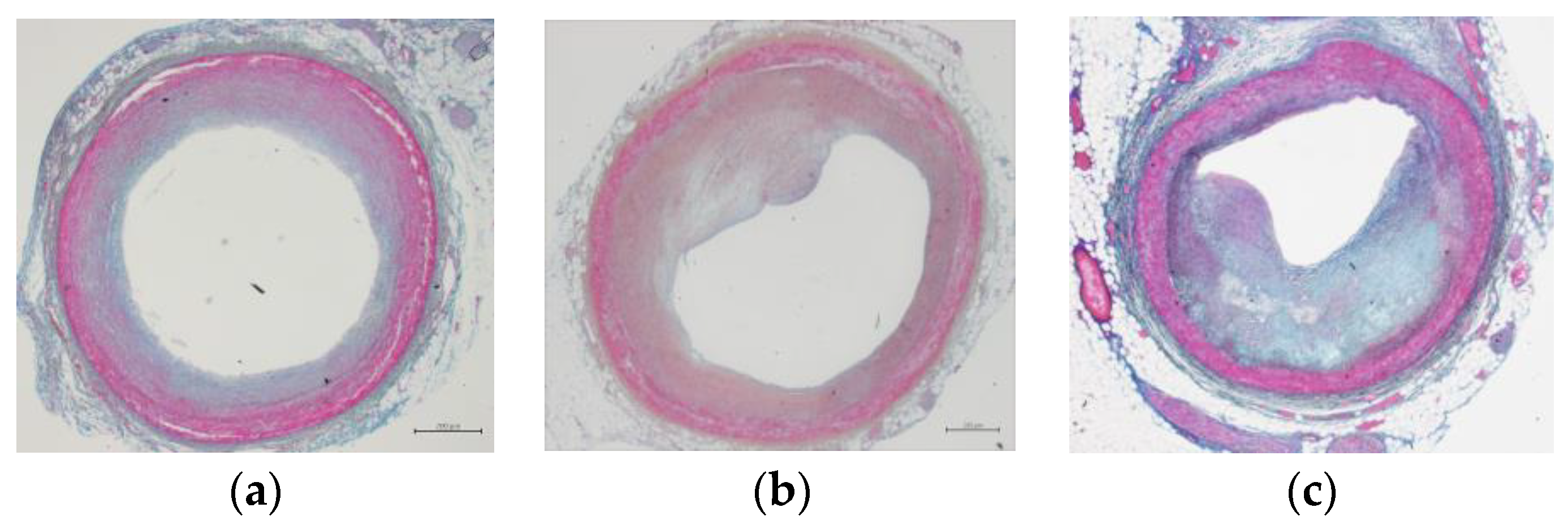
3.1. Characteristics of the Subjects in Coronary Artery Occlusion Status
3.2. Gene Distribution of rs2910164a and rs41291957
3.3. Changes in the miRNA Expression of the Plasma of the Control Samples Kept at Room Temperature
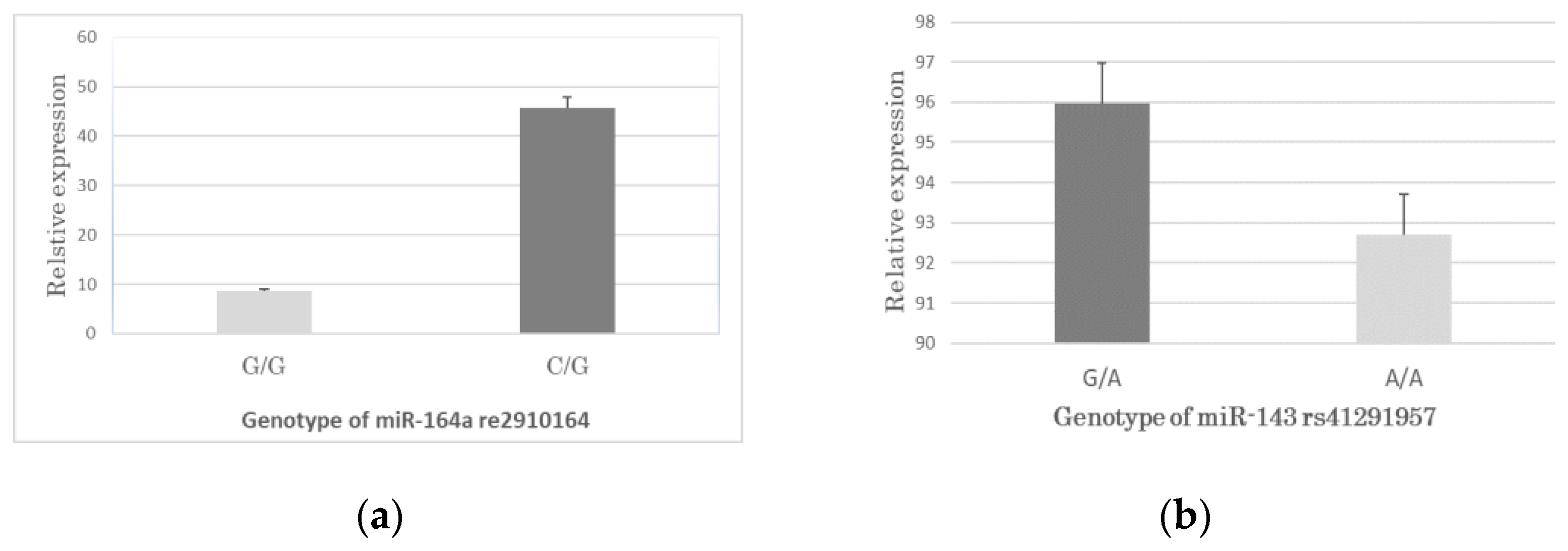
3.4. Association between the SNPs Genotypes and miRNA Expression in CAD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malakar, A.K.; Choudhury, D.; Halder, B.; Paul, B.; Uddin, A. A review on coronary artery disease, its risk factors, and therapeutics. J. Cell Physiol. 2019, 234, 16812–16823. [Google Scholar] [CrossRef]
- Klein, L.; Gheorghiade, M. Coronary artery disease and prevention of heart failure. Med. Clin. N. Am. 2004, 88, 1209–1235. [Google Scholar] [CrossRef] [PubMed]
- Stanford, W.; Thompson, B.H.; Weiss, R.M. Coronary artery calcification: Clinical significance and current methods of detection. AJR Am. J. Roentgenol. 1993, 161, 1139–1146. [Google Scholar] [CrossRef]
- Hill, J.S.; Hayden, M.R.; Frohlich, J.; Pritchard, P.H. Genetic and environmental factors affecting the incidence of coronary artery disease in heterozygous familial hypercholesterolemia. Arter. Thromb. 1991, 11, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Heiba, I.M.; DeMeester, C.A.; Xia, Y.R.; Diep, A.; George, V.T.; Amos, C.I.; Srinivasan, S.R.; Berenson, G.S.; Elston, R.C.; Lusis, A.J. Genetic contributions to quantitative lipoprotein traits associated with coronary artery disease: Analysis of a large pedigree from the Bogalusa Heart Study. Am. J. Med. Genet. 1993, 47, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Tong, Y.; Zhang, H.M.; Wang, K.; Hu, T.; Shan, G.; Sun, J.; Guo, A.Y. Genome-wide identification of SNPs in microRNA genes and the SNP effects on microRNA target binding and biogenesis. Hum. Mutat. 2012, 33, 254–263. [Google Scholar] [CrossRef]
- Wu, D.F.; Yin, R.X.; Cao, X.L.; Chen, W.X. Association between single nucleotide polymorphism rs1044925 and the risk of coronary artery disease and ischemic stroke Figure legends. Int. J. Mol. Sci. 2014, 15, 3546–3559. [Google Scholar] [CrossRef] [PubMed]
- Sarosh, M.; Nurulain, S.M.; Shah, S.T.A.; Jadoon, K.M.; Muneer, Z.; Bibi, N.; Shah, S.F.A.; Hussain, S. Association analysis of single nucleotide polymorphisms in autophagy related 7 (ATG7) gene in patients with coronary artery disease. Medicine 2022, 101, e29776. [Google Scholar] [CrossRef] [PubMed]
- Hombach, S.; Kretz, M. Non-coding RNAs: Classification, Biology and Functioning. Adv. Exp. Med. Biol. 2016, 937, 3–17. [Google Scholar] [PubMed]
- Stępień, E.; Marina, C.C.; Szczepan, K.; Anna, D.; Nuno, C.D.; Francisco, J.E. The circulating non-coding RNA landscape for biomarker research: Lessons and prospects from cardiovascular diseases. Acta. Pharmacol. Sin. 2018, 39, 1085–1099. [Google Scholar]
- Viereck, J.; Thum, T. Circulating Noncoding RNAs as Biomarkers of Cardiovascular Disease and Injury. Circ. Res. 2017, 120, 381–399. [Google Scholar] [CrossRef] [Green Version]
- Fengqiong Liu, F.; Li, R.; Zhang, Y.; Qiu, J.; Ling, W. Association of Plasma MiR-17-92 With Dyslipidemia in Patients with Coronary Artery Disease. Medicine 2014, 93, e98. [Google Scholar]
- Momtazi, A.A.; Banach, M.; Pirro, M.; Evan, A.; Stein, E.A.; Sahebkar, A. MicroRNAs: New Therapeutic Targets for Familial Hypercholesterolemia? Clin. Rev. Allergy Immunol. 2018, 54, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. An overview of microRNAs. Adv. Drug. Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef]
- Huang, Y.; Li, J. MicroRNA 208 family in cardiovascular diseases: Therapeutic implication and potential biomarker. J. Physiol. Biochem. 2015, 71, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, J. MicroRNAs: A novel class of potential therapeutic targets for cardiovascular diseases. Acta. Pharmacol. Sin. 2010, 31, 1–9. [Google Scholar]
- Çakmak, H.A.; Demir, M. MicroRNA and Cardiovascular Diseases. Balkan. Med. J. 2020, 37, 60–71. [Google Scholar] [CrossRef]
- Chen, J.; Wang, D.Z. microRNAs in cardiovascular development. J. Mol. Cell Cardiol. 2012, 52, 949–957. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, J.; Baker, A.H.; Dimmeler, S.; Heymans, S.; Mayr, M.; Thum, T. Non-coding RNAs in vascular disease—From basic science to clinical applications: Scientific update from the Working Group of Myocardial Function of the European Society of Cardiology. Cardiovasc. Res. 2018, 114, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, B.; Yang, Y.X.; Jia, Q.J.; Zhang, A.; Qi, Z.W.; Zhang, J.P. Long Noncoding RNAs in Pathological Cardiac Remodeling: A Review of the Update Literature. Biomed. Res. Int. 2019, 2019, 7159592. [Google Scholar] [CrossRef] [PubMed]
- Correia de Sousa, M.; Gjorgjieva, M.; Dolicka, D.; Sobolewski, C.; Foti, M. Deciphering miRNAs’ Action through miRNA Editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef]
- Sorop, A.; Constantinescu, D.; Cojocaru, F.; Dinischiotu, A.; Cucu, D.; Simona Olimpia Dima, S.O. Exosomal microRNAs as Biomarkers and Therapeutic Targets for Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 4997. [Google Scholar] [CrossRef] [PubMed]
- Deyell, M.W.; Krahn, A.D.; Goldberger, J.J. Sudden cardiac death risk stratification. Circ. Res. 2015, 116, 1907–1918. [Google Scholar] [CrossRef]
- McElwee, S.K.; Velasco, A.; Doppalapudi, H. Mechanisms of sudden cardiac death. J. Nucl. Cardiol. 2016, 23, 1368–1379. [Google Scholar] [CrossRef]
- Tseng, Z.H.; Olgin, J.E.; Vittinghoff, E.; Ursell, P.C.; Kim, A.S.; Karl Sporer, K.; Yeh, C.; Colburn, B.; Clark, N.M.; Khan, R.; et al. Prospective Countywide Surveillance and Autopsy Characterization of Sudden Cardiac Death: POST SCD Study. Circulation 2018, 137, 2689–2700. [Google Scholar] [CrossRef] [PubMed]
- Pinchi, E.; Frati, P.; Aromatario, M.; Cipolloni, L.; Fabbri, M.; Russa, R.L.; Maiese, A.; Neri, M.; Santurro, A.; Scopetti, M.; et al. miR-1, miR-499 and miR-208 are sensitive markers to diagnose sudden death due to early acute myocardial infarction. J. Cell Mol. Med. 2019, 23, 6005–6016. [Google Scholar] [CrossRef]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. Methods Mol. Biol. 2017, 509, 1–10. [Google Scholar]
- Romaine, S.P.R.; Tomaszewski, M.; Condorelli, G.; Samani, N.J. MicroRNAs in cardiovascular disease: An introduction for clinicians. Heart 2015, 101, 921–928. [Google Scholar] [CrossRef]
- Hang, P.; Guo, J.; Sun, C.; Du, Z. MicroRNAs as Candidate Drug Targets for Cardiovascular Diseases. Curr. Drug Targets. 2017, 18, 463–472. [Google Scholar] [CrossRef]
- Rocchi, A.; Chiti, E.; Maiese, A.; Turillazzi, E.; Spinetti, I. MicroRNAs: An Update of Applications in Forensic Science. Diagnostics 2020, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Austen, W.G.; Edwards, J.E.; Frye, R.L.; Gensini, G.G.; Gott, V.L.; Griffith, L.S.; McGoon, D.C.; Murphy, M.L.; Roe, B.B. A reporting system on patients evaluated for coronary artery disease. Report of the Ad Hoc Committee for Grading of Coronary Artery Disease, Council on Cardiovascular Surgery, American Heart Association. Circulation 1975, 51, 5–40. [Google Scholar] [CrossRef]
- Hall, I.F.; Climent, M.; Viviani Anselmi, C.; Papa, L.; Tragante, V.; Lambroia, L.; Farina, F.M.; Kleber, M.E.; März, W.; Biguori, C.; et al. rs41291957 controls miR-143 and miR-145 expression and impacts coronary artery disease risk. EMBO Mol. Med. 2021, 13, e14060. [Google Scholar] [CrossRef] [PubMed]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Zhu, Y.; Wei, Q.K.; Yuan, Y.; Zhou, F.; Ge, Y.Y.; Yang, J.R.; Su, H.; Zhuang, S.M. A functional polymorphism in the miR-146a gene is associated with the risk for hepatocellular carcinoma. Carcinogenesis 2008, 29, 2126–2131. [Google Scholar] [CrossRef]
- Yang, L.; Gao, X.; Luo, H.; Huang, Q.; Wei, Y.; Zhang, G.; Huang, G.; Su, D.; Chen, L.; Lu, C.; et al. No association of pri-miR-143 rs41291957 polymorphism with the risk of congenital heart disease in a Chinese population. Pediatr. Cardiol. 2014, 35, 1057–1061. [Google Scholar] [CrossRef]
- Xie, W.Q.; Tan, S.Y.; Wang, X.F. MiR-146a rs2910164 polymorphism increases risk of gastric cancer: A meta-analysis. World J. Gastroenterol. 2014, 20, 15440–15447. [Google Scholar] [CrossRef]
- Shareef, S.; Ebrahimi, S.O.; Reiisi, S. Contribution of hsa-miR-146a and hsa-miR-223 gene variations in patients with multiple sclerosis reveals association of rs2910164 and rs1044165 with risk of multiple sclerosis susceptibility. J. Investig. Med. 2021, 69, 1015–1021. [Google Scholar] [CrossRef]
- Zhu, F.; Chu, T.; Ruan, Z.; Zhang, M.; Zhou, M.; Zhang, Q.; Zhang, R.; Wu, L. Inflammation-Related MicroRNAs Are Associated with Plaque Stability Calculated by IVUS in Coronary Heart Disease Patients. J. Interv. Cardiol. 2019, 2019, 9723129. [Google Scholar] [CrossRef] [PubMed]
- Verit, F.F.; Yucel, O. Endometriosis, leiomyoma and adenomyosis: The risk of gynecologic malignancy. Asian. Pac. J. Cancer Prev. 2013, 14, 5589–5597. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.S.; Besla, R.; Li, A.; Chen, Z.; Shikatani, E.A.; Nazari-Jahantigh, M.; Hammoutène, A.; Nguyen, M.A.; Geoffrion, M.; Cai, L.; et al. Paradoxical Suppression of Atherosclerosis in the Absence of microRNA-146a. Circ. Res. 2017, 121, 354–367. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Closure Rate (%) | Male | Female | Total |
|---|---|---|---|
| <10 | 28 | 26 | 54 |
| 10–30 | 14 | 14 | 28 |
| 30–50 | 16 | 20 | 36 |
| >50 | 23 | 10 | 33 |
| Total | 81 | 70 | 151 |
| Model | Genotype | CAD Group (%) | Non-CAD Group (%) | OR (95% CI) | p-Value |
|---|---|---|---|---|---|
| rs2910164 C>G | |||||
| Codominant | G/G | 13 (14) | 2 (3.5) | 1 | |
| C/G | 60 (64.5) | 16 (27.6) | 1.61 (0.91–3.35) | 0.006 | |
| C/C | 20 (21.5) | 40 (69) | 1.18 (0.82–2.31) | ||
| Dominant | C/G-G/G | 73 (78.5) | 18 (31) | 1 | |
| C/C | 20 (21.5) | 40 (69) | 1.59 (0.76–2.81) | 0.014 | |
| Recessive | G/G | 13 (14) | 2 (3.5) | 1 | |
| C/C-C/G | 80 (86) | 56 (96.5) | 0.91 (0.53–2.04) | 0.32 | |
| Overdominant | C/G | 60 (64.5) | 16 (27.6) | 1 | |
| C/C-G/G | 33 (35.5) | 42 (72.4) | 1.12 (0.75–2.73) | 0.0049 | |
| Log-additive | 0.0025 | ||||
| rs41291957 G>A | |||||
| Codominant | A/A | 10 (15.2) | 5 (5.9) | 1 | |
| G/A | 20 (30.3) | 34 (40) | 0.81 (0.62–1.23) | 0.26 | |
| G/G | 36 (54.5) | 46 (54.1) | |||
| Dominant | G/A-A/A | 30 (45.5) | 39 (45.9) | 1 | 0.68 |
| G/G | 36 (54.5) | 46 (54.1) | 0.71 (0.53–1.35) | ||
| Recessive | A/A | 10 (15.2) | 5 (5.9) | 1 | 0.17 |
| G/G-G/A | 56 (84.8) | 80 (94.1) | 0.66 (0.21–1.12) | ||
| Overdominant | G/A | 20 (30.3) | 34 (40) | 1 | 0.21 |
| G/G-A/A | 46 (69.7) | 51 (60) | 0.41 (0.19–0.87) | ||
| Log-additive | 0.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tie, J.; Takanari, H.; Ota, K.; Okuda, T. Role of miR-143 and miR-146 in Risk Evaluation of Coronary Artery Diseases in Autopsied Samples. Genes 2023, 14, 471. https://doi.org/10.3390/genes14020471
Tie J, Takanari H, Ota K, Okuda T. Role of miR-143 and miR-146 in Risk Evaluation of Coronary Artery Diseases in Autopsied Samples. Genes. 2023; 14(2):471. https://doi.org/10.3390/genes14020471
Chicago/Turabian StyleTie, Jian, Hiroki Takanari, Koya Ota, and Takahisa Okuda. 2023. "Role of miR-143 and miR-146 in Risk Evaluation of Coronary Artery Diseases in Autopsied Samples" Genes 14, no. 2: 471. https://doi.org/10.3390/genes14020471