
Study on the Association between LRRC8B Gene InDel and Sheep Body Conformation Traits

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Sample Collection and Measurement of Body Size
2.3. Primer Design
2.4. The PCR Amplification System and Procedures
2.5. TA Cloning Experiment
2.6. Data Collation and Analysis
3. Results
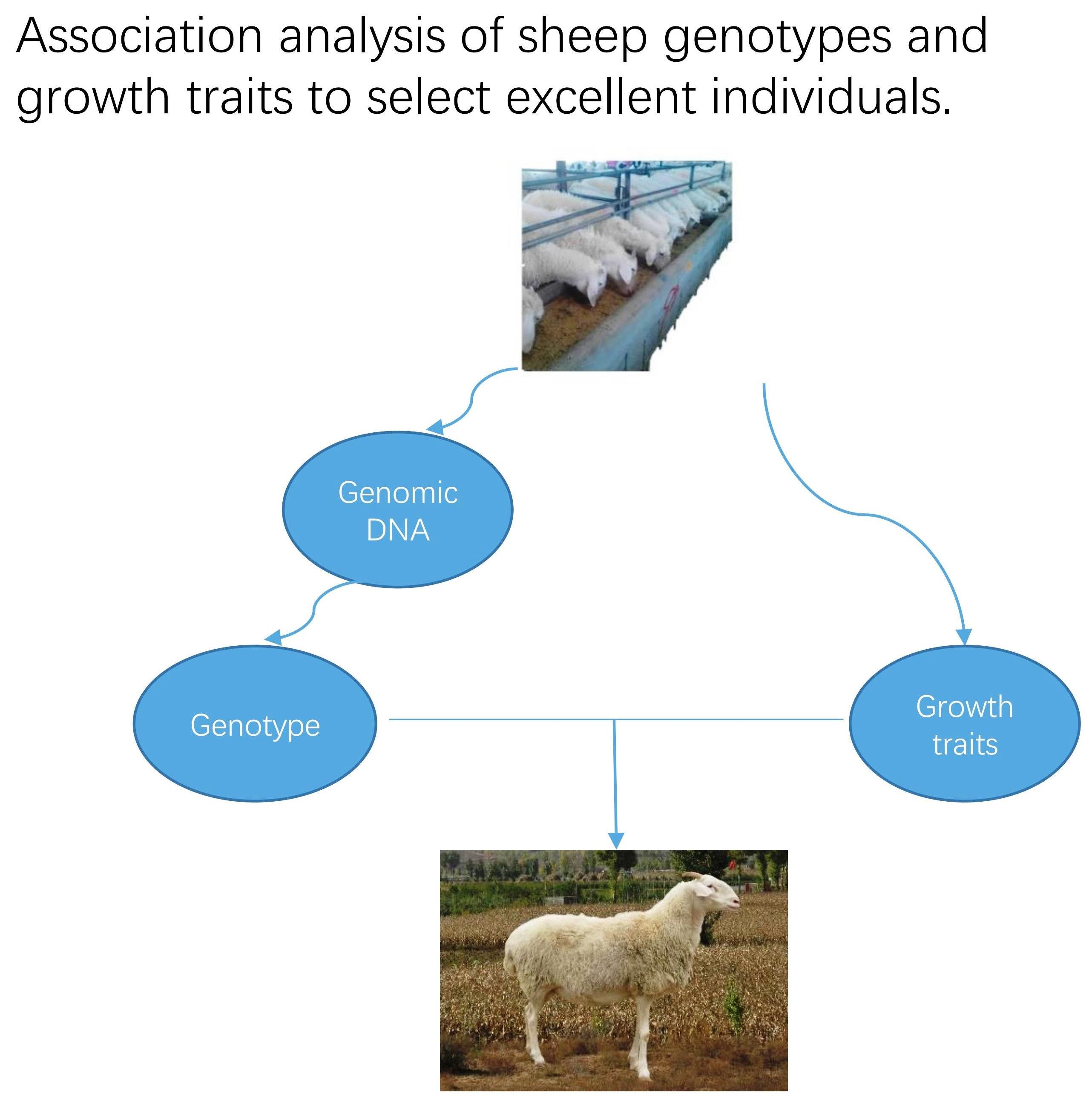
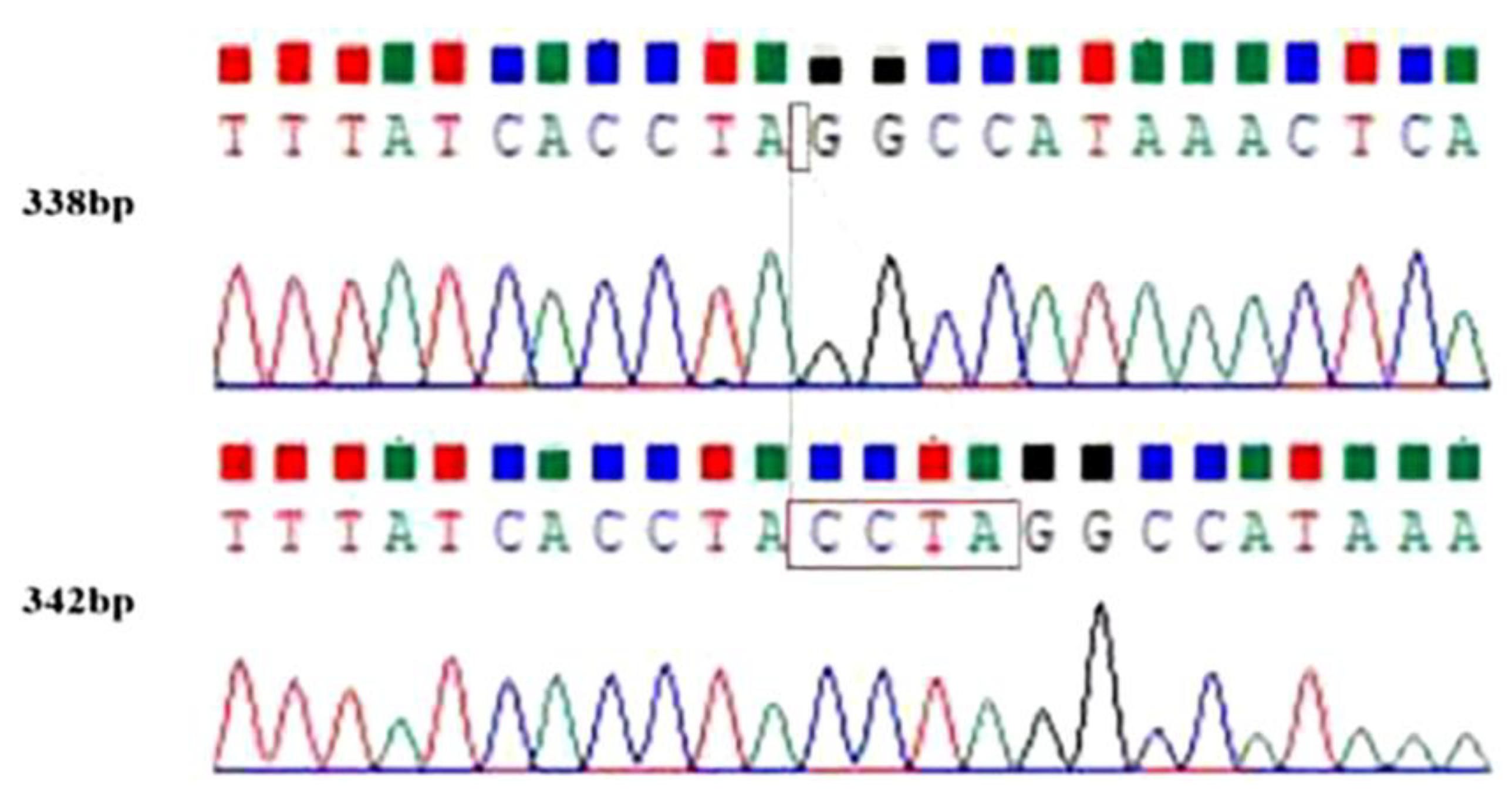
3.1. Screening of the InDel Locus of the LRRC8B Gene
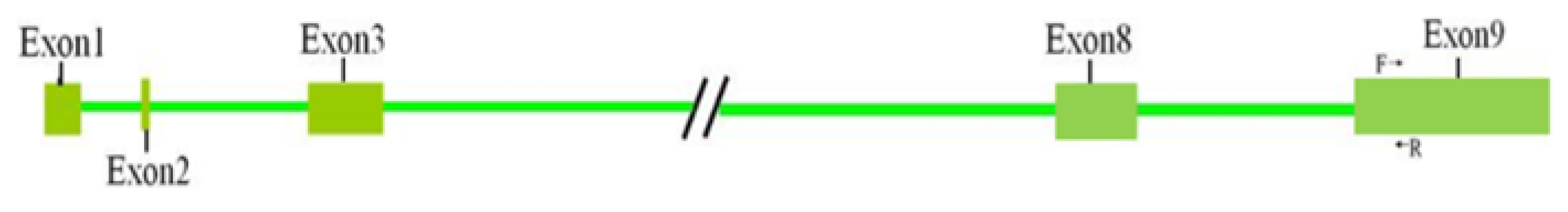
3.2. PCR Results and Genotyping
3.3. InDel Genetic Parameter Analysis of the LRRC8B Gene
3.4. Association Analysis between LRRC8B Gene Polymorphism and Body Conformation Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abascal, F.; Zardoya, R. LRRC8 proteins share a common ancestor with pannexins, and may form hexameric channels involved in cell-cell communication. BioEssays 2012, 34, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, F.K.; Ullrich, F.; Münch, J.; Lazarow, K.; Lutter, D.; Mah, N.; Andrade-Navarro, M.A.; von Kries, J.P.; Stauber, T.; Jentsch, T.J. Identification of LRRC8 heteromers as an essential component of the volume-regulated anion channel VRAC. Science 2014, 344, 634–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Dubin, A.E.; Mathur, J.; Tu, B.; Reddy, K.; Miraglia, L.J.; Reinhardt, J.; Orth, A.P.; Patapoutian, A. SWELL1, a plasma membrane protein, is an essential component of volume-regulated anion channel. Cell 2014, 157, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutter, D.; Ullrich, F.; Lueck, J.C.; Kempa, S.; Jentsch, T.J. Selective transport of neurotransmitters and modulators by distinct volume-regulated LRRC8 anion channels. J. Cell Sci. 2017, 130, 1122–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, B.; Stauber, T. Biophysics and Structure-Function Relationships of LRRC8-Formed Volume-Regulated Anion Channels. Biophys. J. 2019, 116, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Pervaiz, S.; Kopp, A.; von Kleist, L.; Stauber, T. Absolute Protein Amounts and Relative Abundance of Volume-regulated Anion Channel (VRAC) LRRC8 Subunits in Cells and Tissues Revealed by Quantitative Immunoblotting. Int. J. Mol. Sci. 2019, 20, 5879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, Y.; Sato, K.; Numata, T. Pathophysiology and puzzles of the volume-sensitive outwardly rectifying anion channel. J. Physiol. 2009, 587, 2141–2149. [Google Scholar] [CrossRef] [PubMed]
- Lang, F.; Hoffmann, E.K. Role of ion transport in control of apoptotic cell death. Compr. Physiol. 2012, 2, 2037–2061. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.F.; Hoffmann, E.K.; Novak, I. Cell volume regulation in epithelial physiology and cancer. Front. Physiol. 2013, 4, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akita, T.; Okada, Y. Characteristics and roles of the volume-sensitive outwardly rectifying (VSOR) anion channel in the central nervous system. Neuroscience 2014, 275, 211–231. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Klausen, T.K.; Nilius, B. The identification of a volume-regulated anion channel: An amazing Odyssey. Acta physiologica 2015, 213, 868–881. [Google Scholar] [CrossRef] [PubMed]
- Stauber, T. The volume-regulated anion channel is formed by LRRC8 heteromers—Molecular identification and roles in membrane transport and physiology. Biol. Chem. 2015, 396, 975–990. [Google Scholar] [CrossRef] [PubMed]
- Osei-Owusu, J.; Yang, J.; Vitery, M.; Qiu, Z. Molecular Biology and Physiology of Volume-Regulated Anion Channel (VRAC). Curr. Top. Membr. 2018, 81, 177–203. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Wondergem, R.; Morrison, R.; Yin, V.P.; Strange, K. Leucine-rich repeat containing protein LRRC8A is essential for swelling-activated Cl- currents and embryonic development in zebrafish. Physiol. Rep. 2016, 4, e12940. [Google Scholar] [CrossRef]
- Syeda, R.; Qiu, Z.; Dubin, A.E.; Murthy, S.E.; Florendo, M.N.; Mason, D.E.; Mathur, J.; Cahalan, S.M.; Peters, E.C.; Montal, M.; et al. LRRC8 Proteins Form Volume-Regulated Anion Channels that Sense Ionic Strength. Cell 2016, 164, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Hyzinski-García, M.C.; Rudkouskaya, A.; Mongin, A.A. LRRC8A protein is indispensable for swelling-activated and ATP-induced release of excitatory amino acids in rat astrocytes. J. Physiol. 2014, 592, 4855–4862. [Google Scholar] [CrossRef] [PubMed]
- Strange, K.; Yamada, T.; Denton, J.S. A 30-year journey from volume-regulated anion currents to molecular structure of the LRRC8 channel. J. Gen. Physiol. 2019, 151, 100–117. [Google Scholar] [CrossRef] [Green Version]
- Schober, A.L.; Wilson, C.S.; Mongin, A.A. Molecular composition and heterogeneity of the LRRC8-containing swelling-activated osmolyte channels in primary rat astrocytes. J. Physiol. 2017, 595, 6939–6951. [Google Scholar] [CrossRef] [Green Version]
- Gradogna, A.; Gaitán-Peñas, H.; Boccaccio, A.; Estévez, R.; Pusch, M. Cisplatin activates volume sensitive LRRC8 channel mediated currents in Xenopus oocytes. Channels 2017, 11, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Khandelwal, N.; Kumar, A.; Bera, A.K. Leucine-rich repeat-containing 8B protein is associated with the endoplasmic reticulum Ca2+ leak in HEK293 cells. J. Cell Sci. 2017, 130, 3818–3828. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, H.; Kang, J.S.; Zhang, L.; Su, J.; Li, H.Y.; Sun, L.K. 5-Nitro-2-(3-phenylpropylamino) benzoic acid induced drug resistance to cisplatin in human erythroleukemia cell lines. Anat. Rec. 2011, 294, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Xu, Y.; Zhou, L.; Yu, H.M.; Kang, J.S.; Liu, N.; Quan, C.S.; Sun, L.K. Suppression of chloride channel 3 expression facilitates sensitivity of human glioma U251 cells to cisplatin through concomitant inhibition of Akt and autophagy. Anat. Rec. 2013, 296, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, B.H.; Dam, C.S.; Stürup, S.; Lambert, I.H. Dual role of LRRC8A-containing transporters on cisplatin resistance in human ovarian cancer cells. J. Inorg. Biochem. 2016, 160, 287–295. [Google Scholar] [CrossRef]
- Gasparoni, G.; Bultmann, S.; Lutsik, P.; Kraus, T.; Sordon, S.; Vlcek, J.; Dietinger, V.; Steinmaurer, M.; Haider, M.; Mulholland, C.B.; et al. DNA methylation analysis on purified neurons and glia dissects age and Alzheimer’s disease-specific changes in the human cortex. Epigenetics Chromatin 2018, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, T.; Ma, S.F.; Machado, R.F.; Bhorade, S.M.; Garcia, J.G. MicroRNAs Implicated in Dysregulation of Gene Expression Following Human Lung Transplantation. Transl. Respir. Med. 2013, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Liao, X.; Stothard, P.; Chung, W.H.; Jeon, H.J.; Miller, S.P.; Choi, S.Y.; Lee, J.K.; Yang, B.; Lee, K.T.; et al. Whole-genome analyses of Korean native and Holstein cattle breeds by massively parallel sequencing. PLoS ONE 2014, 9, e101127. [Google Scholar] [CrossRef]
- Weber, J.L.; David, D.; Heil, J.; Fan, Y.; Zhao, C.F.; Marth, G. Human diallelic insertion/deletion polymorphisms. Am. J. Hum. Genet. 2002, 71, 854–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jander, G.; Norris, S.R.; Rounsley, S.D.; Bush, D.F.; Levin, I.M.; Last, R.L. Arabidopsis map-based cloning in the post-genome era. Plant Physiol. 2002, 129, 440–450. [Google Scholar] [CrossRef] [Green Version]
- Varshney, R.K.; Nayak, S.N.; May, G.D.; Jackson, S.A. Next-generation sequencing technologies and their implications for crop genetics and breeding. Trends Biotechnol. 2009, 27, 522–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, Z.; Liu, X.; Chai, Y.; Yang, P.; Li, J.; Huang, Y.; Li, L.; Huang, W.; Yang, G.; et al. Copy number variation of WBP1L gene revealed its association with growth traits across chinese cattle populations. J. Agric. Sci. 2022. [CrossRef]
- Xu, W.; He, H.; Zheng, L.; Xu, J.W.; Lei, C.Z.; Zhang, G.M.; Dang, R.H.; Niu, H.; Qi, X.L.; Chen, H.; et al. Detection of 19-bp deletion within PLAG1 gene and its effect on growth traits in cattle. Gene 2018, 675, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.; Yang, P.; Liu, Y.; Song, T.; Zhang, Z.; Shi, Q.; Chen, F.; Liu, X.; Li, Z.; Ru, B.; et al. Genetic effects of MOGAT1 gene SNP in growth traits of Chinese cattle. Gene 2021, 769, 145201. [Google Scholar] [CrossRef] [PubMed]
- Kvikstad, E.M.; Tyekucheva, S.; Chiaromonte, F.; Makova, K.D. A macaque’s-eye view of human insertions and deletions: Differences in mechanisms. PLoS Comput. Biol. 2007, 3, 1772–1782. [Google Scholar] [CrossRef]
- Britten, R.J.; Rowen, L.; Williams, J.; Cameron, R.A. Majority of divergence between closely related DNA samples is due to indels. Proc. Natl. Acad. Sci. USA 2003, 100, 4661–4665. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Zhao, H.; Chen, N.; Cao, X.; Hanif, Q.; Pi, L.; Hu, L.; Chaogetu, B.; Huang, Y.; Lan, X.; et al. Population structure, genetic diversity, and selective signature of Chaka sheep revealed by whole genome sequencing. BMC Genom. 2020, 21, 520. [Google Scholar] [CrossRef]
- Bai, M.; Sun, L.; Jia, C.; Li, J.; Han, Y.; Liu, H.; Chen, Y.; Jiang, H. Integrated Analysis of miRNA and mRNA Expression Profiles Reveals Functional miRNA-Targets in Development Testes of Small Tail Han Sheep. G3 2019, 9, 523–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Chen, X.; Liu, C.; Han, X.; Xiao, C.; Yi, S.; Huang, D. Genetic analysis of 32 InDels in four ethnic minorities from Chinese Xinjiang. PLoS ONE 2021, 16, e0250206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jin, Y.; Jiang, F.; Cheng, H.; Wang, Y.; Lan, X.; Song, E. Relationship between an indel mutation within the SIRT4 gene and growth traits in Chinese cattle. Anim. Biotechnol. 2019, 30, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Q.; Wang, K.; Zhang, S.; Pan, C.; Chen, H.; Qu, L.; Yan, H.; Lan, X. A novel 12-bp indel polymorphism within the GDF9 gene is significantly associated with litter size and growth traits in goats. Anim. Genet. 2017, 48, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Su, P.; Akhatayeva, Z.; Pan, C.; Zhang, Q.; Lan, X. Novel InDel variations of the Cry2 gene are associated with litter size in Australian White sheep. Theriogenology 2022, 179, 155–161. [Google Scholar] [CrossRef]
- Ren, F.; Yu, S.; Chen, R.; Lv, X.; Pan, C. Identification of a novel 12-bp insertion/deletion (indel) of iPS-related Oct4 gene and its association with reproductive traits in male piglets. Anim. Reprod. Sci. 2017, 178, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.; Ren, T.; Li, W.; Liang, J.; Mo, G.; Luo, W.; He, D.; Liang, S.; Zhang, X. A Novel 65-bp Indel in the GOLGB1 Gene Is Associated with Chicken Growth and Carcass Traits. Animals 2020, 10, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Xu, H.; Akhatayeva, Z.; Liu, H.; Lin, C.; Han, X.; Lu, X.; Lan, X.; Zhang, Q.; Pan, C. Novel indel variations of the sheep FecB gene and their effects on litter size. Gene 2021, 767, 145176. [Google Scholar] [CrossRef] [PubMed]
- Erdenee, S.; Akhatayeva, Z.; Pan, C.; Cai, Y.; Xu, H.; Chen, H.; Lan, X. An insertion/deletion within the CREB1 gene identified using the RNA-sequencing is associated with sheep body morphometric traits. Gene 2021, 775, 145444. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | Amplification Length/bp | |
|---|---|---|---|
| LRRC8B | Forward primer | ACTTGGAGGGTAGAATGGGG | 342 bp |
| Reverse primer | ACAGGCAGGCACTTTCTCAG |
| Number | Genotype Frequencies | Gene Frequencies | HWec | Genetic Parameter Estimation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ID | DD | I | D | Ho | He | Ne | PIC | |||
| CKS | 269 | 0.572 (154) | 0.428 (115) | 0.286 | 0.714 | p < 0.05 | 0.591 | 0.409 | 1.691 | 0.409 |
| STHS | 149 | 0.409 (61) | 0.591 (88) | 0.205 | 0.795 | p < 0.05 | 0.674 | 0.326 | 1.483 | 0.326 |
| Body Conformation Traits | Genotype (Mean ± SE) | p Value | |
|---|---|---|---|
| ID | DD | ||
| Wither height/cm | 66.29 ± 4.70 | 65.57 ± 4.50 | 0.470 |
| Body length/cm | 72.18 ± 6.80 | 72.14 ± 7.30 | 0.314 |
| Chest circumference/cm | 89.95 ± 8.63 | 88.63 ± 8.26 | 0.589 |
| Body weight/cm | 54.87 ± 13.42 | 53.26 ± 13.18 | 0.367 |
| Body Conformation Traits | Genotype (Mean ± SE) | p Value | |
|---|---|---|---|
| ID | DD | ||
| Body length/cm | 59.60 ± 5.63 | 59.44 ± 6.17 | 0.946 |
| Chest width/cm | 19.28 ± 2.67 | 19.51 ± 3.53 | 0.907 |
| Wither height/cm | 63.61 ± 4.09 | 63.36 ± 4.44 | 0.841 |
| Chest depth/cm | 27.23 ± 2.02 b | 28.13 ± 2.85 a | 0.044 * |
| Chest circumference/cm | 72.11 ± 5.74 | 72.64 ± 6.47 | 0.685 |
| Cannon bone circumference/cm | 7.24 ± 0.66 | 7.10 ± 0.79 | 0.382 |
| Height at hip cross/cm | 63.59 ± 3.85 | 62.87 ± 4.46 | 0.406 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Toremurat, Z.; Liang, Y.; Cheng, J.; Sun, Z.; Huang, Y.; Liu, J.; Chaogetu, B.; Ren, G.; Chen, H. Study on the Association between LRRC8B Gene InDel and Sheep Body Conformation Traits. Genes 2023, 14, 356. https://doi.org/10.3390/genes14020356
Zhang J, Toremurat Z, Liang Y, Cheng J, Sun Z, Huang Y, Liu J, Chaogetu B, Ren G, Chen H. Study on the Association between LRRC8B Gene InDel and Sheep Body Conformation Traits. Genes. 2023; 14(2):356. https://doi.org/10.3390/genes14020356
Chicago/Turabian StyleZhang, Jiaqiang, Zhansaya Toremurat, Yilin Liang, Jie Cheng, Zhenzhen Sun, Yangming Huang, Junxia Liu, BUREN Chaogetu, Gang Ren, and Hong Chen. 2023. "Study on the Association between LRRC8B Gene InDel and Sheep Body Conformation Traits" Genes 14, no. 2: 356. https://doi.org/10.3390/genes14020356