

Functional Analyses of Rare Germline Missense BRCA1 Variants Located within and outside Protein Domains with Known Functions
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids and Construction of BRCA1 Variants
2.2. Cell Culture and Transfection
2.3. Assessment of BRCA1 Protein Expression by Immunoblotting
2.4. RNA Purification and qPCR
2.5. MG132 Assay for Assessment of Proteasomal Degradation
2.6. Cycloheximide Chase Assay for Measurement of BRCA1 Protein Stability
2.7. Fractionation Assay for Assessment of Subcellular Localisation
2.8. Co-Immunoprecipitation Assay
2.9. Statistics
2.10. Assessment of Variant Classifications
3. Results
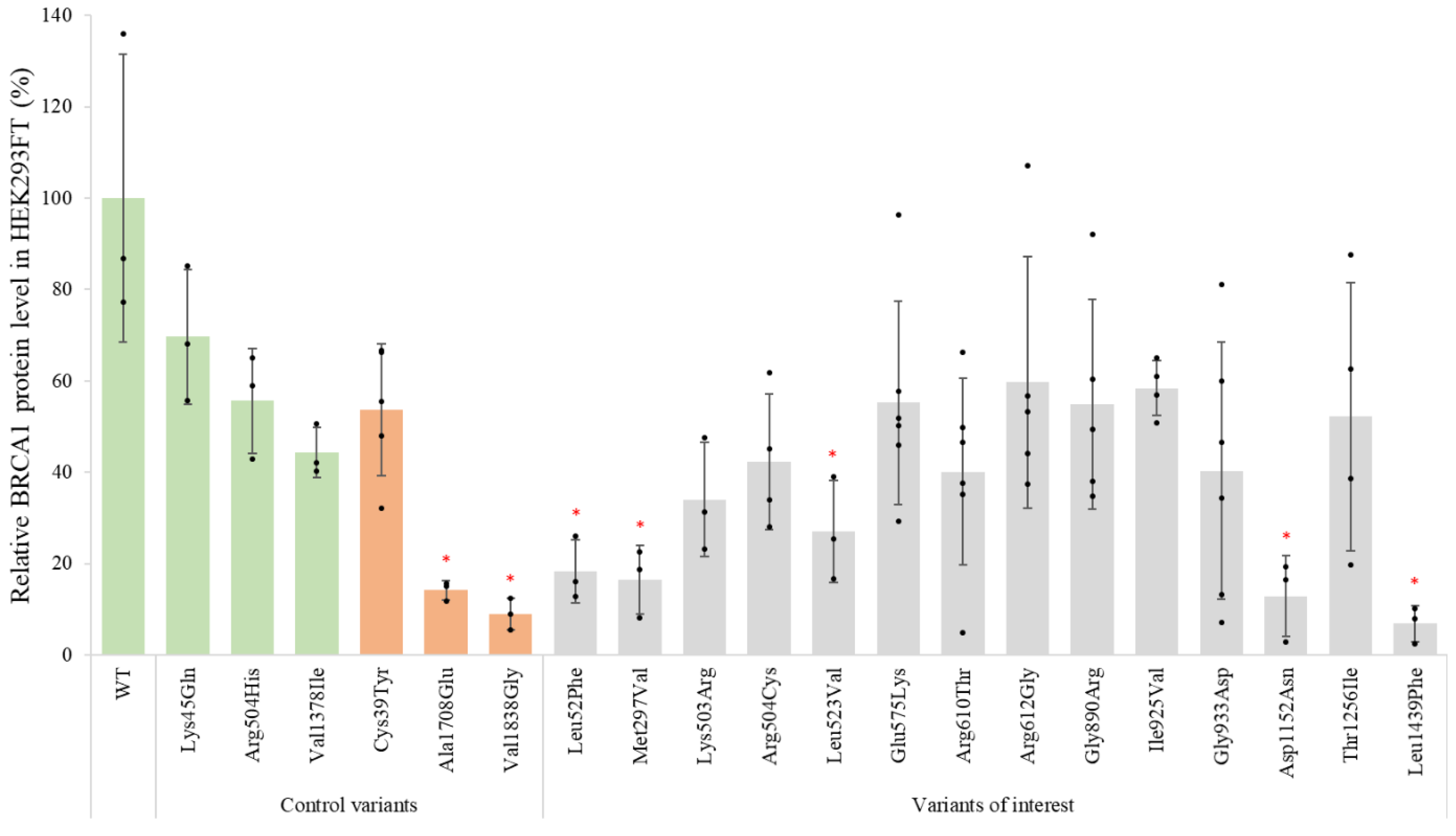
3.1. Effects of BRCA1 Variants on Protein Expression
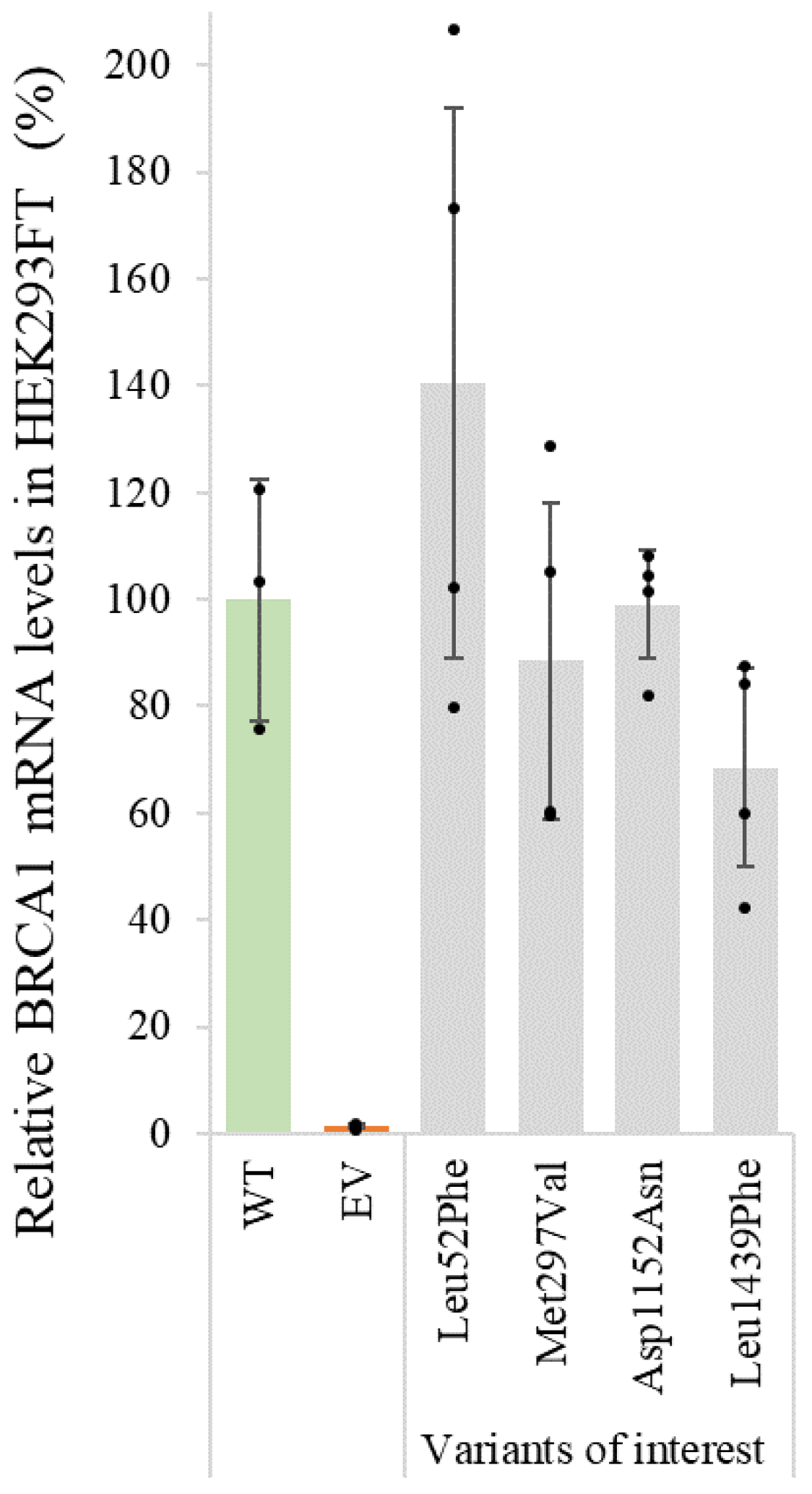
3.2. qPCR for Assessment of mRNA Levels
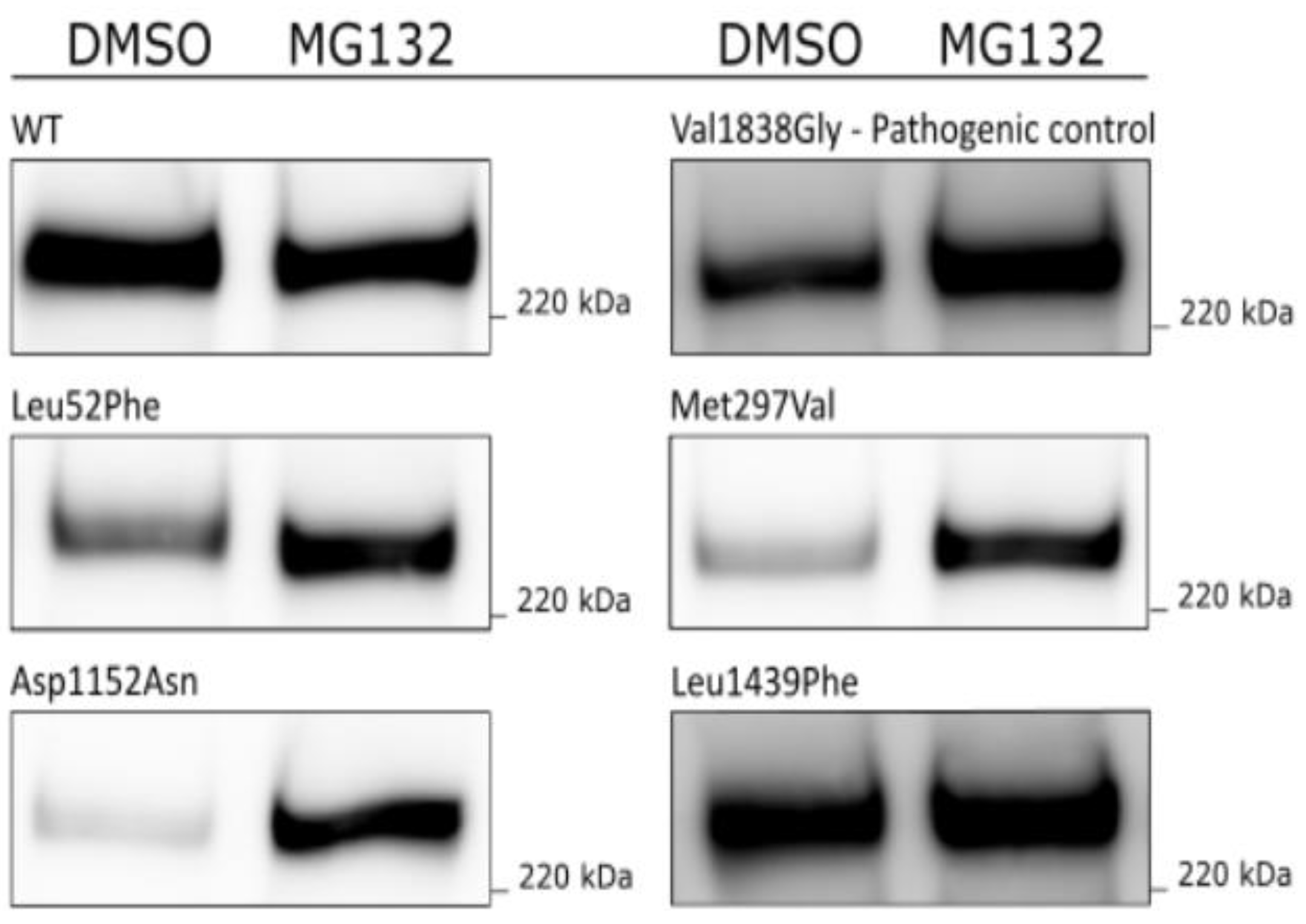
3.3. MG132 Assay for Assessment of Proteasomal Degradation
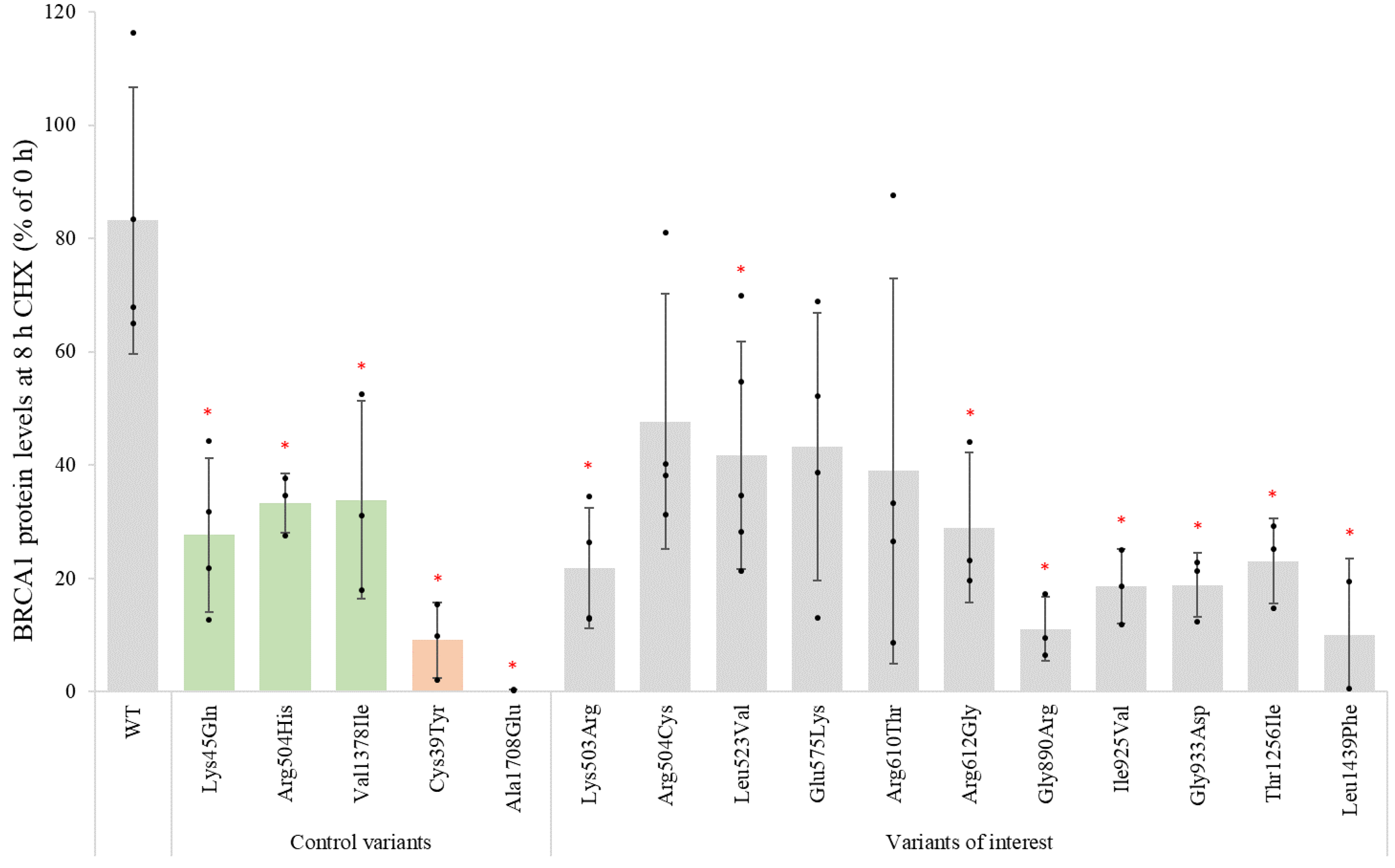
3.4. Cycloheximide Chase Assay for Assessment of Protein Stability
3.5. Assessment of Subcellular Localisation by Fractionation Assay
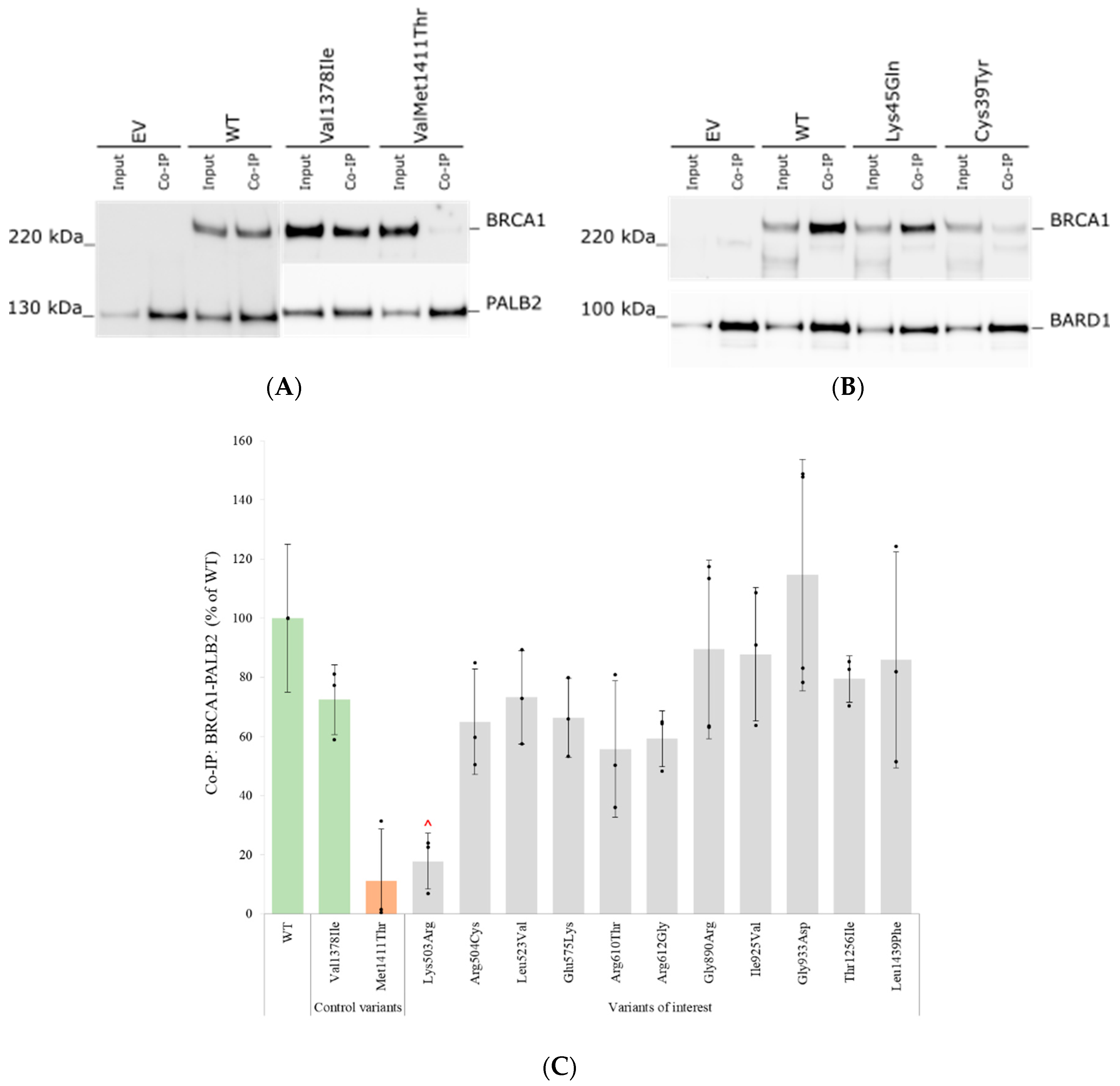
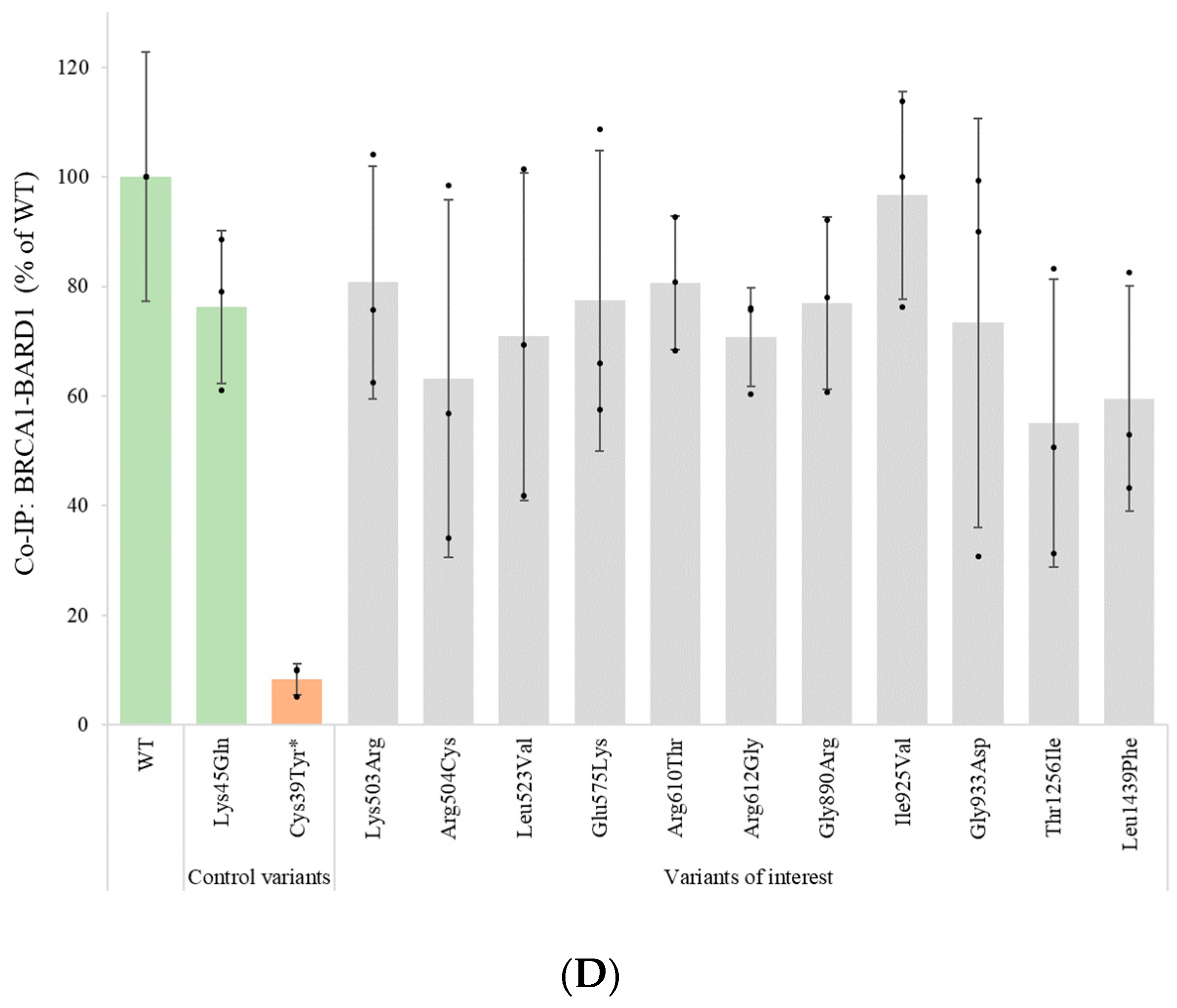
3.6. Assessment of Protein–Protein Interactions with BARD1 and PALB2 by Co-Immunoprecipitation
3.7. Summary of Functional Assays
4. Discussion
4.1. BRCA1 Protein Domains and Amino Acid Conservation
4.2. Protein Expression and Protein Stability of the BRCA1 Variants
4.3. The Effect of BRCA1 Variants on BARD1 and PALB2 Interaction
4.4. Variant Interpretation of the Investigated VUSs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antoniou, A.; Pharoah, P.D.P.; Narod, S.; Risch, H.A.; Eyfjord, J.E.; Hopper, J.L.; Loman, N.; Olsson, H.; Johannsson, O.; Borg, Å.; et al. Average Risks of Breast and Ovarian Cancer Associated with BRCA1 or BRCA2 Mutations Detected in Case Series Unselected for Family History: A Combined Analysis of 22 Studies. Am. J. Hum. Genet. 2003, 72, 1117–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, R.; Ludwig, T. The BRCA1/BARD1 heterodimer, a tumor suppressor complex with ubiquitin E3 ligase activity. Curr. Opin. Genet Dev. 2002, 12, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Mallery, D.L.; Vandenberg, C.J.; Hiom, K. Activation of the E3 ligase function of the BRCA1/BARD1 complex by polyubiquitin chains. EMBO J. 2002, 21, 6755–6762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Pao, G.M.; Chen, H.W.; Verma, I.M.; Hunter, T. Enhancement of BRCA1 E3 Ubiquitin Ligase Activity through Direct Interaction with the BARD1 Protein. J. Biol. Chem. 2003, 278, 5255–5263. [Google Scholar] [CrossRef] [Green Version]
- De Siervi, A.; De Luca, P.; Byun, J.S.; Di, L.J.; Fufa, T.; Haggerty, C.M.; Vazquez, E.; Moiola, C.; Longo, D.L.; Gardner, K. Transcriptional Autoregulation by BRCA1. Cancer Res. 2010, 70, 532–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, R.; Chun, J.; Powell, S.N. BRCA1 and BRCA2: Different roles in a common pathway of genome protection. Nat. Rev. Cancer 2011, 12, 68–78. [Google Scholar] [CrossRef] [Green Version]
- Mark, W.-Y.; Liao, J.C.C.; Lu, Y.; Ayed, A.; Laister, R.; Szymczyna, B.; Chakrabartty, A.; Arrowsmith, C.H. Characterization of Segments from the Central Region of BRCA1: An Intrinsically Disordered Scaffold for Multiple Protein–Protein and Protein–DNA Interactions? J. Mol. Biol. 2005, 345, 275–287. [Google Scholar] [CrossRef]
- Anantha, R.W.; Simhadri, S.; Foo, T.K.; Miao, S.; Liu, J.; Shen, Z.; Ganesan, S.; Xia, B. Functional and mutational landscapes of BRCA1 for homology-directed repair and therapy resistance. Elife 2017, 6, e21350. [Google Scholar] [CrossRef]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef] [Green Version]
- Brnich, S.E.A.T.; Ahmad, N.; Couch, F.J.; Cutting, G.R.; Greenblatt, M.S.; Heinen, C.D.; Kanavy, D.M.; Luo, X.; McNulty, S.M.; Starita, L.M.; et al. Recommendations for application of the functional evidence PS3/BS3 criterion using the ACMG/AMP sequence variant interpretation framework. Genome Med. 2019, 12, 3. [Google Scholar] [CrossRef]
- Findlay, G.M.; Daza, R.M.; Martin, B.; Zhang, M.D.; Leith, A.P.; Gasperini, M.; Janizek, J.D.; Huang, X.; Starita, L.M.; Shendure, J. Accurate classification of BRCA1 variants with saturation genome editing. Nature 2018, 562, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, P.; van der Heijden, I.; van der Gulden, H.; De Bruijn, R.; Braspenning, M.E.; Moghadasi, S.; Wessels, L.F.A.; Vreeswijk, M.P.G.; Jonkers, J. Functional categorization of BRCA1 variants of uncertain clinical significance in homologous recombination repair complementation assays. Clin. Cancer Res. 2020, 26, 4559–4568. [Google Scholar] [CrossRef]
- Fernandes, V.C.; Golubeva, V.A.; Di Pietro, G.; Shields, C.; Amankwah, K.; Nepomuceno, T.C.; de Gregoriis, G.; Abreu, R.B.V.; Harro, C.; Gomes, T.T.; et al. Impact of amino acid substitutions at secondary structures in the BRCT domains of the tumor suppressor BRCA1: Implications for clinical annotation. J. Biol. Chem. 2019, 294, 5980–5992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petitalot, A.; Dardillac, E.; Jacquet, E.; Nhiri, N.; Guirouilh-Barbat, J.; Julien, P.; Bouazzaoui, I.; Bonte, D.; Feunteun, J.; Schnell, J.; et al. Combining Homologous Recombination and Phosphopeptide-binding Data to Predict the Impact of BRCA1 BRCT Variants on Cancer Risk. Mol. Cancer Res. 2019, 17, 54–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starita, L.M.; Islam, M.M.; Banerjee, T.; Adamovich, A.I.; Gullingsrud, J.; Fields, S.; Shendure, J.; Parvin, J.D. A Multiplex Homology-Directed DNA Repair Assay Reveals the Impact of More Than 1,000 BRCA1 Missense Substitution Variants on Protein Function. Am. J. Hum. Genet. 2018, 103, 498–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CanVIG-UK Gene Specific Recommendations: BRCA1/BRCA2. Available online: https://www.cangene-canvaruk.org/gene-specific-recommendations (accessed on 1 June 2022).
- Langerud, J.; Jarhelle, E.; Van Ghelue, M.; Ariansen, S.L.; Iversen, N. Trans-activation-based risk assessment of BRCA1 BRCT variants with unknown clinical significance. Hum. Genom. 2018, 12, 51. [Google Scholar] [CrossRef]
- Jarhelle, E.; Stensland, H.M.F.R.; Mæhle, L.; Van Ghelue, M. Characterization of BRCA1 and BRCA2 variants found in a Norwegian breast or ovarian cancer cohort. Fam. Cancer 2016, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.R.; Pangon, L.; Boutell, C.; Katagiri, T.; Keep, N.H.; Solomon, E. Genetic analysis of BRCA1 ubiquitin ligase activity and its relationship to breast cancer susceptibility. Hum. Mol. Genet. 2006, 15, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Green, R.; Glover, J.N.M.; Marsillac, S.M.; Coquelle, N.; Williams, R.S.; Yeung, T.; Foo, D.; Hau, D.D.; Hui, B.; et al. Comprehensive Analysis of Missense Variations in the BRCT Domain of BRCA1 by Structural and Functional Assays. Cancer Res. 2010, 70, 4880–4890. [Google Scholar] [CrossRef] [Green Version]
- Glover, J.N.M.; Williams, R.S.; Lee, M.S.; Hau, D.D. Structural basis of phosphopeptide recognition by the BRCT domain of BRCA1. Nat. Struct. Mol. Biol. 2004, 11, 519–525. [Google Scholar]
- Adamovich, A.I.; Diabate, M.; Banerjee, T.; Nagy, G.; Smith, N.; Duncan, K.; Mendoza, M.E.; Prida, G.; Freitas, M.A.; Starita, L.M.; et al. The functional impact of BRCA1 BRCT domain variants using multiplexed DNA double-strand break repair assays. Am. J. Hum. Genet. 2022, 109, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Christou, C.M.; Hadjisavvas, A.; Kyratzi, M.; Flouri, C.; Neophytou, I.; Anastasiadou, V.; Loizidou, M.A.; Kyriacou, K. The BRCA1 Variant p.Ser36Tyr Abrogates BRCA1 Protein Function and Potentially Confers a Moderate Risk of Breast Cancer. PLoS ONE 2014, 9, e93400. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.F.; Deffenbaugh, A.M.; Pruss, D.; Frye, C.; Wenstrup, R.J.; Allen-Brady, K.; Tavtigian, S.V.; Monteiro, A.N.; Iversen, E.S.; Couch, F.J.; et al. A Systematic Genetic Assessment of 1,433 Sequence Variants of Unknown Clinical Significance in the BRCA1 and BRCA2 Breast Cancer–Predisposition Genes. Am. J. Hum. Genet. 2007, 81, 873–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavtigian, S.V.; Byrnes, G.B.; Goldgar, D.E.; Thomas, A. Classification of rare missense substitutions, using risk surfaces, with genetic- and molecular-epidemiology applications. Hum. Mutat. 2008, 29, 1342–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallée, M.P.; Di Sera, T.L.; Nix, D.A.; Paquette, A.M.; Parsons, M.T.; Bell, R.; Hoffman, A.; Hogervorst, F.B.L.; Goldgar, D.E.; Spurdle, A.B.; et al. Adding In Silico Assessment of Potential Splice Aberration to the Integrated Evaluation of BRCA Gene Unclassified Variants. Hum. Mutat. 2016, 37, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Dines, J.N.; Shirts, B.H.; Slavin, T.P.; Walsh, T.; King, M.-C.; Fowler, D.M.; Pritchard, C.C. Systematic misclassification of missense variants in BRCA1 and BRCA2 “coldspots”. Genet. Med. 2020, 22, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Garrett, A.; Callaway, A.; Durkie, M.; Cubuk, C.; Alikian, M.; Burghel, G.J.; Robinson, R.; Izatt, L.; Talukdar, S.; Side, L.; et al. Cancer Variant Interpretation Group UK (CanVIG-UK): An exemplar national subspecialty multidisciplinary network. J. Med. Genet. 2020, 57, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, I.; Pérez-Alvarado, G.C.; Parker, D.; Dyson, H.J.; Montminy, M.R.; Wright, P.E. Solution Structure of the KIX Domain of CBP Bound to the Transactivation Domain of CREB: A Model for Activator:Coactivator Interactions. Cell 1997, 91, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Rosen, M.K.; Kim, A.S.; Kakalis, L.T.; Abdul-Manan, N.; Liu, G.A. Autoinhibition and activation mechanisms of the Wiskott-Aldrich syndrome protein. Nature 2000, 404, 151–158. [Google Scholar]
- Kriwacki, R.W.; Lacy, E.R.; Filippov, I.; Lewis, W.S.; Otieno, S.; Xiao, L.; Weiss, S.; Hengst, L. p27 binds cyclin-CDK complexes through a sequential mechanism involving binding-induced protein folding. Nat. Struct. Mol. Biol. 2004, 11, 358–364. [Google Scholar]
- Kriwacki, R.W.; Hengst, L.; Tennant, L.; Reed, S.I.; Wright, P.E. Structural studies of p21Waf1/Cip1/Sdi1 in the free and Cdk2-bound state: Conformational disorder mediates binding diversity. Proc. Natl. Acad. Sci. USA 1996, 93, 11504–11509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovland, H.N.; Al-Adhami, R.; Ariansen, S.L.; Van Ghelue, M.; Sjursen, W.; Lima, S.; Bolstad, M.; Berger, A.; Høberg-Vetti, H.; Knappskog, P.; et al. BRCA1 Norway: Comparison of classification for BRCA1 germline variants detected in families with suspected hereditary breast and ovarian cancer between different laboratories. Fam. Cancer 2022, 21, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Orthwein, A.; Noordermeer, S.M.; Wilson, M.D.; Landry, S.; Enchev, R.I.; Sherker, A.; Munro, M.; Pinder, J.; Salsman, J.; Dellaire, G.; et al. A mechanism for the suppression of homologous recombination in G1 cells. Nature 2015, 528, 422–426. [Google Scholar] [CrossRef]
- Kurihara, M.; Mano, T.; Saito, Y.; Murayama, S.; Toda, T.; Iwata, A. Colocalization of BRCA1 with Tau Aggregates in Human Tauopathies. Brain Sci. 2019, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Evidence-based Network for the Interpretation of Germline Mutant Alleles. Available online: https://enigmaconsortium.org/library/general-documents/enigma-classification-criteria/z (accessed on 24 January 2022).
- Gaboriau, D.C.A.; Rowling, P.J.; Morrison, C.G.; Itzhaki, L.S. Protein stability versus function: Effects of destabilizing missense mutations on BRCA1 DNA repair activity. Biochem. J. 2015, 466, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Tarsounas, M.; Sung, P. The antitumorigenic roles of BRCA1-BARD1 in DNA repair and replication. Nat. Rev. Mol. Cell Biol. 2020, 21, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Ruffner, H.; Joazeiro, C.A.; Hemmati, D.; Hunter, T.; Verma, I.M. Cancer-Predisposing Mutations within the RING Domain of BRCA1: Loss of Ubiquitin Protein Ligase Activity and Protection from Radiation Hypersensitivity. Proc. Natl. Acad. Sci. USA 2001, 98, 5134–5139. [Google Scholar] [CrossRef] [Green Version]
- Malikova, J.; Kaci, A.; Dusatkova, P.; Aukrust, I.; Torsvik, J.; Vesela, K.; Kankova, P.D.; Njølstad, P.R.; Pruhova, S.; Bjørkhaug, L. Functional analyses of HNF1A-MODY variants refine the interpretation of identified sequence variants. J. Clin. Endocrinol. Metab. 2020, 105, e1377–e1386. [Google Scholar] [CrossRef]
- Althari, S.; Najmi, L.A.; Bennett, A.J.; Aukrust, I.; Rundle, J.K.; Colclough, K.; Molnes, J.; Kaci, A.; Nawaz, S.; van der Lugt, T.; et al. Unsupervised Clustering of Missense Variants in HNF1A Using Multidimensional Functional Data Aids Clinical Interpretation. Am. J. Hum. Genet. 2020, 107, 670–682. [Google Scholar] [CrossRef]
- Sy, S.M.H.; Huen, M.S.Y.; Chen, J. PALB2 Is an Integral Component of the BRCA Complex Required for Homologous Recombination Repair. Proc. Natl. Acad. Sci. USA 2009, 106, 7155–7160. [Google Scholar] [CrossRef] [Green Version]
- Woods, N.T.; Baskin, R.; Golubeva, V.; Jhuraney, A.; De-Gregoriis, G.; Vaclova, T.; Goldgar, D.E.; Couch, F.J.; Carvalho, M.A.; Iversen, E.S.; et al. Functional assays provide a robust tool for the clinical annotation of genetic variants of uncertain significance. NPJ Genom. Med. 2016, 1, 16001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, S.L.; Rodriguez, A.M.; Snyder, R.R.; Hankins, G.D.; Boehning, D. Structure-Function Of The Tumor Suppressor BRCA1. Comput. Struct. Biotechnol. J. 2012, 1, e201204005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. Combining evidence for and against pathogenicity for variants in cancer susceptibility genes: CanVIG-UK consensus recommendations. J. Med. Genet. 2021, 58, 297–304. [Google Scholar]
- CanVIG-UK Consensus Specification for Cancer Susceptibility Genes ACGS Best Practice Guidelines for Variant Classification (v2.16). Available online: https://www.cangene-canvaruk.org/canvig-uk-guidance (accessed on 1 June 2022).
- Ioannidis, N.M.; Rothstein, J.H.; Pejaver, V.; Middha, S.; McDonnell, S.K.; Baheti, S.; Musolf, A.; Li, Q.; Holzinger, E.; Karyadi, D.; et al. REVEL: An Ensemble Method for Predicting the Pathogenicity of Rare Missense Variants. Am. J. Hum. Genet. 2016, 99, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Yue, P.; Li, Z.; Moult, J. Loss of Protein Structure Stability as a Major Causative Factor in Monogenic Disease. J. Mol. Biol. 2005, 353, 459–473. [Google Scholar] [CrossRef]
- Shi, Z.; Sellers, J.; Moult, J. Protein stability and in vivo concentration of missense mutations in phenylalanine hydroxylase. Proteins 2012, 80, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Yates, C.M.; Sternberg, M.J.E. The Effects of Non-Synonymous Single Nucleotide Polymorphisms (nsSNPs) on Protein–Protein Interactions. J. Mol. Biol. 2013, 425, 3949–3963. [Google Scholar] [CrossRef]
- Nepomuceno, T.C.; dos Santos, A.P.P.; Fernandes, V.C.; Elias, A.B.R.; Gomes, T.T.; Suarez-Kurtz, G.; Iversen, E.S.; Couch, F.J.; Monteiro, A.N.A.; Carvalho, M.A. Assessment of small in-frame indels and C-terminal nonsense variants of BRCA1 using a validated functional assay. Sci. Rep. 2022, 12, 16203. [Google Scholar] [CrossRef]
- Kim, S.; Jin, H.; Seo, H.-R.; Lee, H.J.; Lee, Y.-S. Regulating BRCA1 protein stability by cathepsin S-mediated ubiquitin degradation. Cell Death Differ. 2019, 26, 812–825. [Google Scholar] [CrossRef] [Green Version]
- Rowling, P.J.E.; Cook, R.; Itzhaki, L.S. Toward Classification of BRCA1 Missense Variants Using a Biophysical Approach. J. Biol. Chem. 2010, 285, 20080–20087. [Google Scholar] [CrossRef] [Green Version]
- Glover, J.N.M.; Williams, R.S.; Green, R. Crystal structure of the BRCT repeat region from the breast cancer-associated protein BRCA1. Nat. Struct. Biol. 2001, 8, 838–842. [Google Scholar]
- Williams, R.S.; Chasman, D.I.; Hau, D.D.; Hui, B.; Lau, A.Y.; Glover, J.N. Detection of Protein Folding Defects Caused by BRCA1-BRCT Truncation and Missense Mutations. J. Biol. Chem. 2003, 278, 53007–53016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lu, G.; Li, L.; Yi, J.; Yan, K.; Wang, Y.; Zhu, B.; Kuang, J.; Lin, M.; Zhang, S.; et al. HUWE1 interacts with BRCA1 and promotes its degradation in the ubiquitin–proteasome pathway. Biochem. Biophys. Res. Commun. 2014, 444, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, K.; Takano, N.; Yamada, Y.; Kazama, H.; Tokuhisa, M.; Hino, H.; Fujita, K.; Barroga, E.; Hiramoto, M.; Handa, H.; et al. BRCA1 degradation in response to mitochondrial damage in breast cancer cells. Sci. Rep. 2021, 11, 8735. [Google Scholar] [CrossRef] [PubMed]
- Ransburgh, D.J.R.; Chiba, N.; Ishioka, C.; Toland, A.E.; Parvin, J.D. Identification of Breast Tumor Mutations in BRCA1 That Abolish Its Function in Homologous DNA Recombination. Cancer Res. 2010, 70, 988–995. [Google Scholar] [CrossRef]
- Starita, L.M.; Young, D.L.; Islam, M.; Kitzman, J.O.; Gullingsrud, J.; Hause, R.J.; Fowler, D.M.; Parvin, J.D.; Shendure, J.; Fields, S. Massively Parallel Functional Analysis of BRCA1 RING Domain Variants. Genetics 2015, 200, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Towler, W.I.; Zhang, J.; Ransburgh, D.J.R.; Toland, A.E.; Ishioka, C.; Chiba, N.; Parvin, J.D. Analysis of BRCA1 Variants in Double-Strand Break Repair by Homologous Recombination and Single-Strand Annealing. Hum. Mutat. 2013, 34, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Kais, Z.; Chiba, N.; Ishioka, C.; Parvin, J.D. Functional differences among BRCA1 missense mutations in the control of centrosome duplication. Oncogene 2012, 31, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Lu, C.; Xie, M.; Wendl, M.C.; Wang, J.; McLellan, M.D.; Leiserson, M.D.M.; Huang, K.-L.; Wyczalkowski, M.A.; Jayasinghe, R.; Banerjee, T.; et al. Patterns and functional implications of rare germline variants across 12 cancer types. Nat. Commun. 2015, 6, 10086. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant cDNA | Protein | Protein Expression (% of WT) | qPCR (% of WT) | % Reduction in Protein Levels (after 8 h CHX Treatment) | Inhibition of Proteolytic Degradation (MG132) | Co-IP BARD1 (% of WT) | Co-IP PALB2 (% of WT) | Nuclear Localization (% of Total) | Summary Functional Assays |
|---|---|---|---|---|---|---|---|---|---|
| WT protein | 100 ± 31 | 100 ± 23 | 83 ± 23 | Equal amounts | 100 ± 23 | 100 ± 25 | 84 ± 10 | ||
| c.154C > T | p.Leu52Phe | 18 ± 7 | 141 ± 51 | Increased protein levels | 77 ± 5 | Reduced protein expression | |||
| c.889A > G | p.Met297Val | 16 ± 8 | 89 ± 30 | Increased protein levels | 82 ± 7 | Reduced protein expression | |||
| c.1508A > G | p.Lys503Arg | 34 ± 12 | 22 ± 11 | 81 ± 21 | 18 ± 9 | 81 ± 12 | No deviations detected | ||
| c.1510C > T | p.Arg504Cys | 42 ± 15 | 48 ± 23 | 63 ± 33 | 65 ± 18 | 71 ± 16 | No deviations detected | ||
| c.1567T > G | p.Leu523Val | 27 ± 11 | 42 ± 20 | 71 ± 30 | 73 ± 16 | 88 ± 1 | No deviations detected | ||
| c.1723G > A | p.Glu575Lys | 55 ± 22 | 43 ± 24 | 77 ± 27 | 66 ± 13 | 87 ± 10 | No deviations detected | ||
| c.1829G > C | p.Arg610Thr | 40 ± 20 | 39 ± 34 | 81 ± 12 | 56 ± 23 | 87 ± 2 | No deviations detected | ||
| c.1834A > G | p.Arg612Gly | 60 ± 28 | 29 ± 13 | 71 ± 9 | 59 ± 9 | 88 ± 9 | No deviations detected | ||
| c.2668G > A | p.Gly890Arg | 55 ± 23 | 11 ± 6 | 77 ± 16 | 89 ± 30 | 90 ± 10 | Reduced protein stability | ||
| c.2773A > G | p.Ile925Val | 58 ± 6 | 19 ± 7 | 97 ± 19 | 88 ± 23 | 95 ± 6 | No deviations detected | ||
| c.2798G > A | p.Gly933Asp | 40 ± 28 | 19 ± 6 | 73 ± 37 | 115 ± 39 | 88 ± 12 | No deviations detected | ||
| c.3454G > A | p.Asp1152Asn | 13 ± 9 | 99 ± 10 | Increased protein levels | 88 ± 10 | Reduced protein expression | |||
| c.3767C > T | p.Thr1256Ile | 52 ± 29 | 23 ± 7 | 55 ± 26 | 79 ± 8 | 90 ± 2 | No deviations detected | ||
| c.4315C > T | p.Leu1439Phe | 7 ± 4 | 69 ± 19 | 10 ± 13 | Equal amounts | 60 ± 20 | 86 ± 37 | 82 ± 13 | Reduced protein expression and stability |
| Benign control variants | |||||||||
| c.133A > C | p.Lys45Gln | 70 ± 15 | 28 ± 14 | 76 ± 14 | |||||
| c.1511G > A | p.Arg504His | 56 ± 11 | 33 ± 5 | ||||||
| c.4132G > A | p.Val1378Ile | 44 ± 5 | 34 ± 17 | 72 ± 11 | |||||
| Pathogenic control variants | |||||||||
| c.116G > A | p.Cys39Tyr | 9 ± 7 | 8 ± 3 | ||||||
| c.4232T > C | p.Met1411Thr | 11 ± 18 | |||||||
| c.5513T > G | p.Val1838Gly | 9 ± 3 | Increased protein levels | ||||||
| c.5123C > A | p.Ala1708Glu | 14 ± 2 | 0 ± 0 | ||||||
| Variant | Region/Domain | Functional Studies Recommended by CanVIG-UK * | Results in Functional Assays in This Study | GnomAD MAF % (Allele Count) ** | REVEL *** | Splicing **** | ClinVar Classifications | CanVIG-UK Criteria | Original Class ***** | Revised Class |
|---|---|---|---|---|---|---|---|---|---|---|
| c.154C > T p.(Leu52Phe) | RING | Functional HRR [15], functional saturating genome assay [11] | Reduced protein expression and increased proteasomal degradation | 0.09354 (24) | 0.68 | Possible effect | VUSx7, LBx4, Bx2 | BS1_strong | VUS | VUS |
| c.889A > G p.(Met297Val) | Reduced protein expression and increased proteasomal degradation | - (1) | 0.58 | VUSx6 | BP1, PS3_sup, PM2_sup | LB, VUS | VUS | |||
| c.1508A > G p.(Lys503Arg) | NLS | No deviations detected | 0.0003240 (2) | 0.76 | VUSx4, LBx3 | BP1, BS3_sup, PP3, PM2_sup | LB, VUS | VUS | ||
| c.1510C > T p.(Arg504Cys) | NLS | No deviations detected | 0.0003240 (4) | 0.65 | VUSx10 | BP1, BS3_sup, PM2_sup | VUS | LB | ||
| c.1567T > G p.(Leu523Val) | No deviations detected | - (1) | 0.61 | VUSx2 | BP1, BS3_sup, PM2_sup | VUS | LB | |||
| c.1723G > A p.(Glu575Lys) | No deviations detected | 0.0009700 (5) | 0.62 | VUSx6 | BP1, BS3_sup, PM2_sup | VUS | LB | |||
| c.1829G > C p.(Arg610Thr) | NLS | No deviations detected | - (0) | 0.74 | VUSx1 | BP1, BS3_sup, PP3, PM2_mod | VUS | VUS | ||
| c.1834A > G p.(Arg612Gly) | NLS | No deviations detected | - (0) | 0.60 | VUSx6, LBx1, Bx1 (ENIGMA) | BS3_sup, BP6, PM2_mod | VUS | LB | ||
| c.2668G > A p.(Gly890Arg) | Reduced protein stability | - (0) | 0.39 | Possible effect | VUSx3, LBx5 | PS3_sup, PM2_mod | VUS | VUS | ||
| c.2773A > G p.(Ile925Val) | No deviations detected | 0.0003240 (2) | 0.19 | VUSx4, LBx3 | BP1, BP4, BS3_sup, PM2_sup | LB, VUS | LB | |||
| c.2798G > A p.(Gly933Asp) | Neutral in cisplatin, olaparib and DR-GFP HRR assays [12] | No deviations detected | - (0) | 0.29 | Possible effect | VUSx5, LBx1 | BS3_strong, BS4_sup, PM2_mod | LB, VUS | LB | |
| c.3454G > A p.(Asp1152Asn) | Neutral in cisplatin, olaparib and DR-GFP HRR assays [12] | Reduced protein expression and increased proteasomal degradation | 0.004516 (10) | 0.37 | VUSx10, LBx6, Bx1 | BP1, BP4 | LB | VUS • | ||
| c.3767C > T p.(Thr1256Ile) | No deviations detected | - (0) | 0.42 | VUSx1 | BP1, BS3_sup, PM2_mod | VUS | LB | |||
| c.4315C > T p.(Leu1439Phe) | Neutral in cisplatin and DR-GFP HRR assays, not clear in olaparib assay [12] | Reduced protein expression and stability | 0.0007760(3) | 0.25 | VUSx3, LBx1 | BP1, BP4, PM2_sup | LB, VUS | VUS • |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hovland, H.N.; Mchaina, E.K.; Høberg-Vetti, H.; Ariansen, S.L.; Sjursen, W.; Van Ghelue, M.; Haukanes, B.I.; Knappskog, P.M.; Aukrust, I.; Ognedal, E. Functional Analyses of Rare Germline Missense BRCA1 Variants Located within and outside Protein Domains with Known Functions. Genes 2023, 14, 262. https://doi.org/10.3390/genes14020262
Hovland HN, Mchaina EK, Høberg-Vetti H, Ariansen SL, Sjursen W, Van Ghelue M, Haukanes BI, Knappskog PM, Aukrust I, Ognedal E. Functional Analyses of Rare Germline Missense BRCA1 Variants Located within and outside Protein Domains with Known Functions. Genes. 2023; 14(2):262. https://doi.org/10.3390/genes14020262
Chicago/Turabian StyleHovland, Henrikke Nilsen, Eunice Kabanyana Mchaina, Hildegunn Høberg-Vetti, Sarah Louise Ariansen, Wenche Sjursen, Marijke Van Ghelue, Bjørn Ivar Haukanes, Per Morten Knappskog, Ingvild Aukrust, and Elisabet Ognedal. 2023. "Functional Analyses of Rare Germline Missense BRCA1 Variants Located within and outside Protein Domains with Known Functions" Genes 14, no. 2: 262. https://doi.org/10.3390/genes14020262