
Molecular Characterization and Phylogenetic Analysis of Casein Gene Family in Camelus ferus
 , , ,
, , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
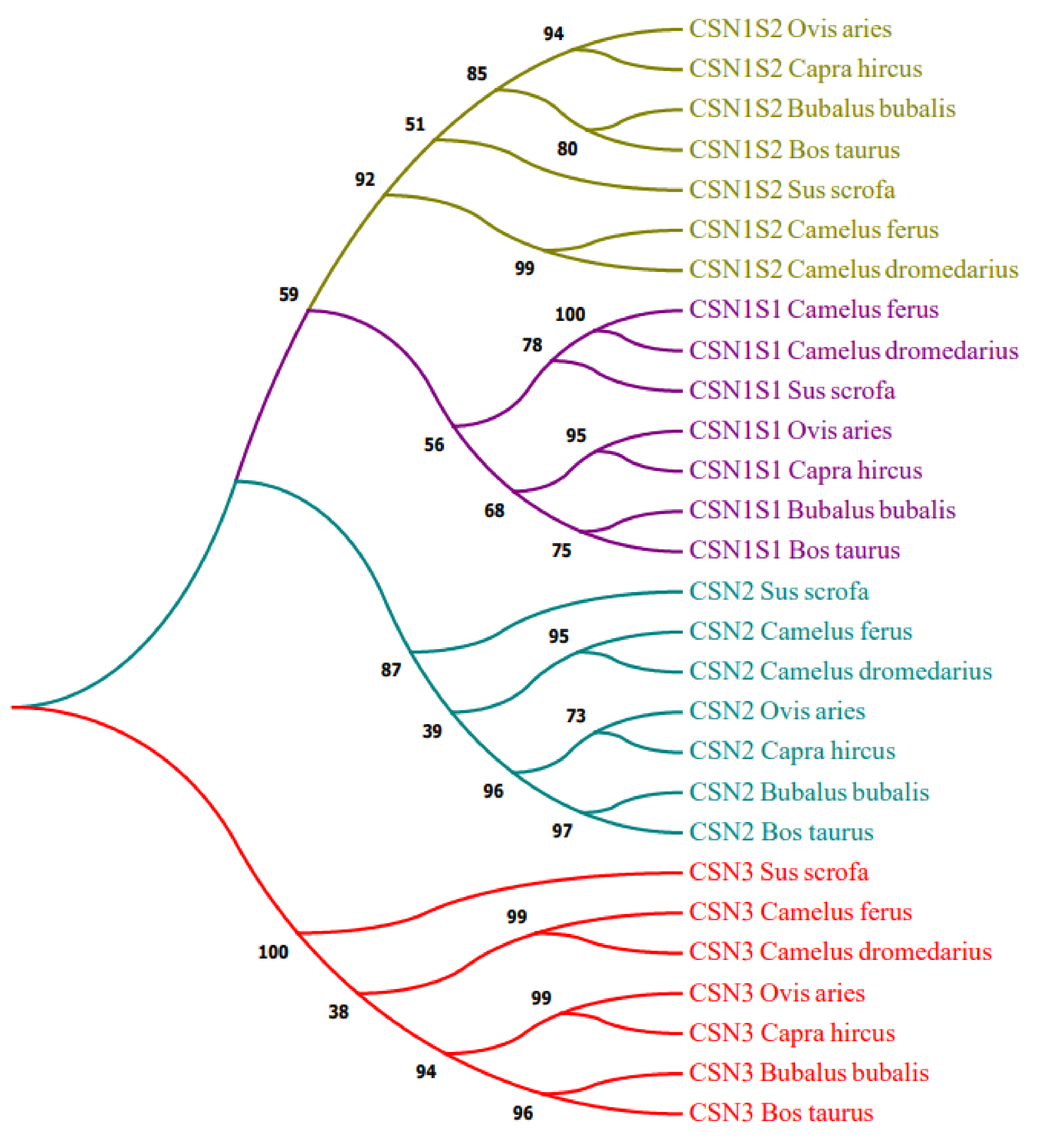
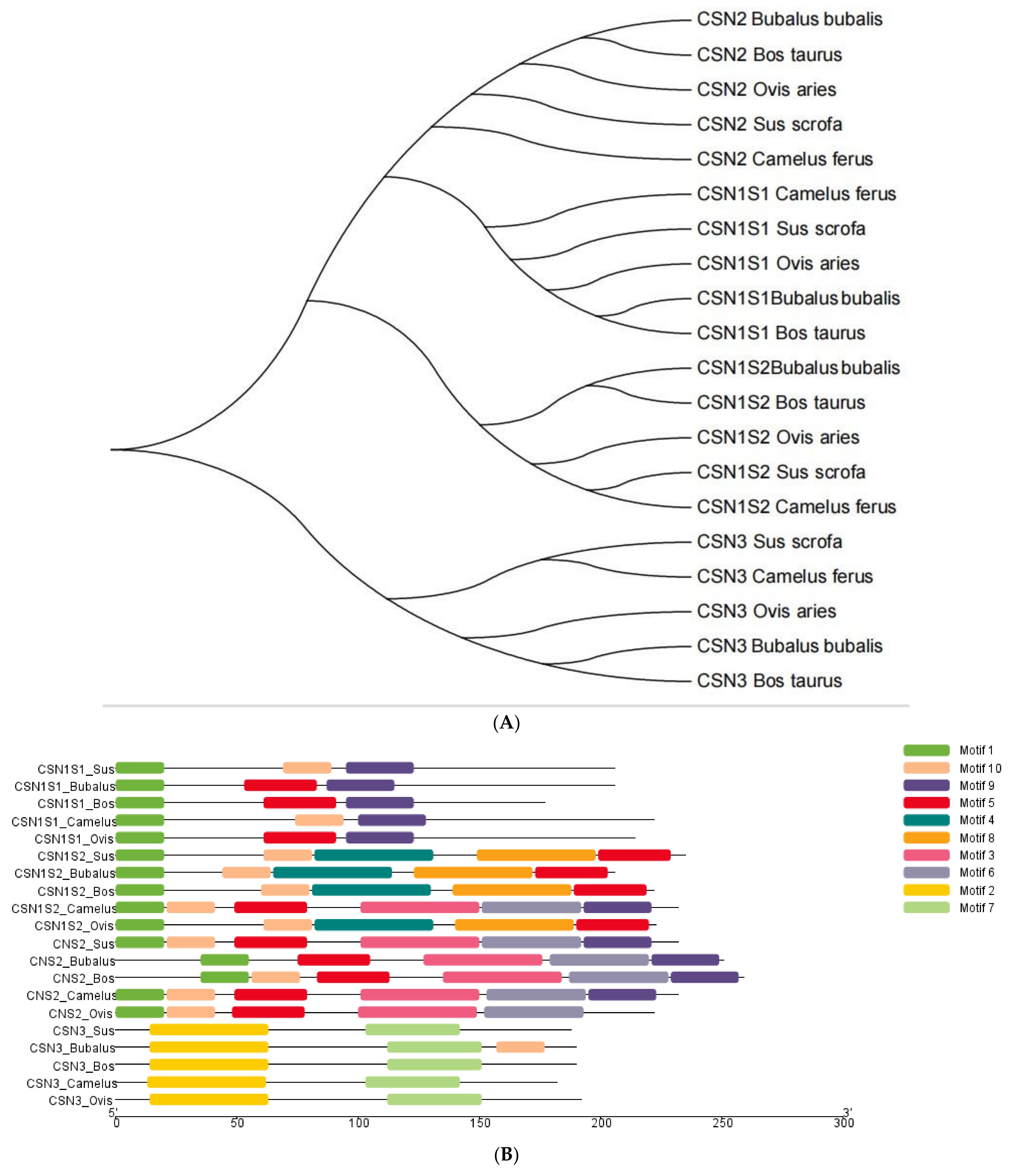
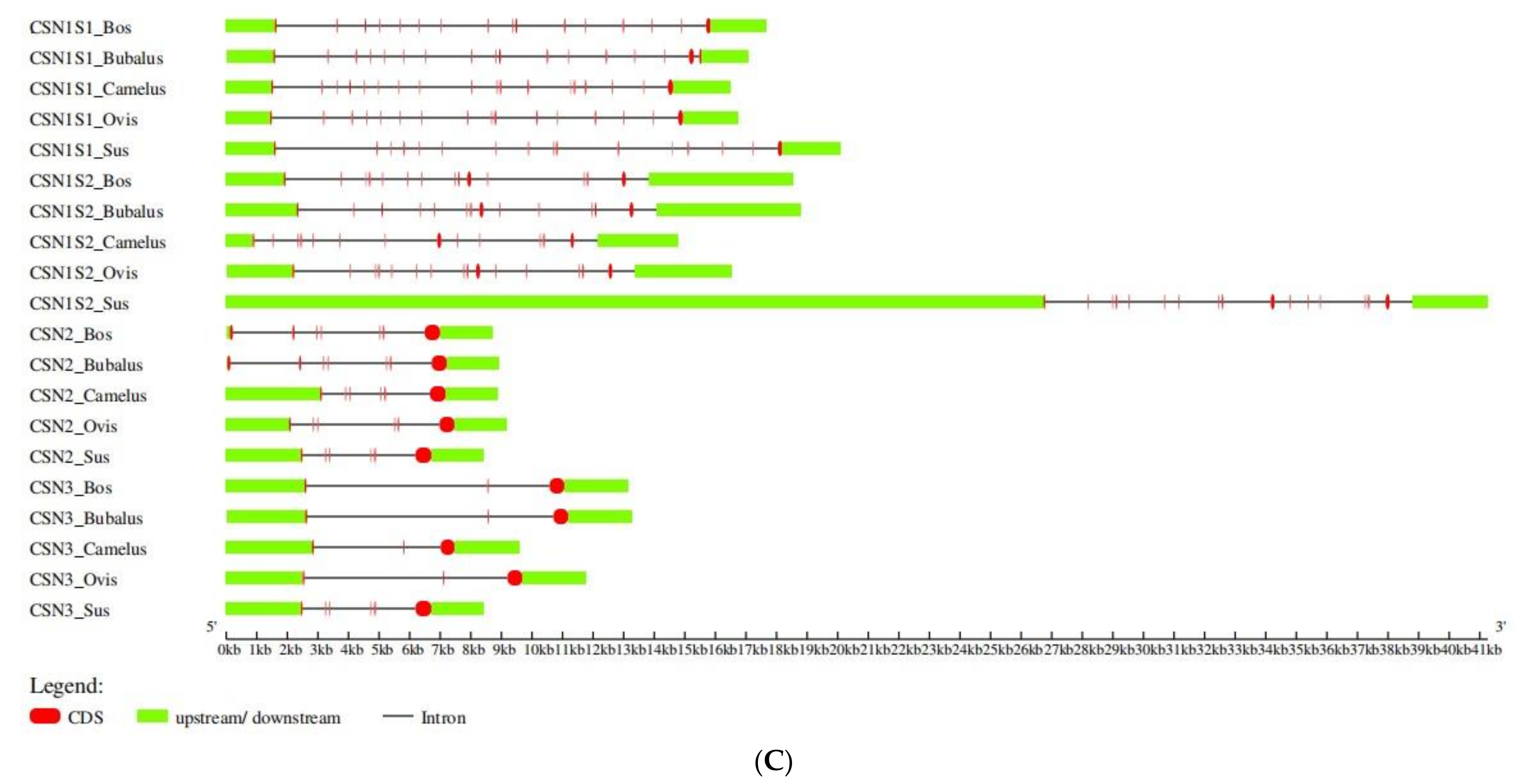
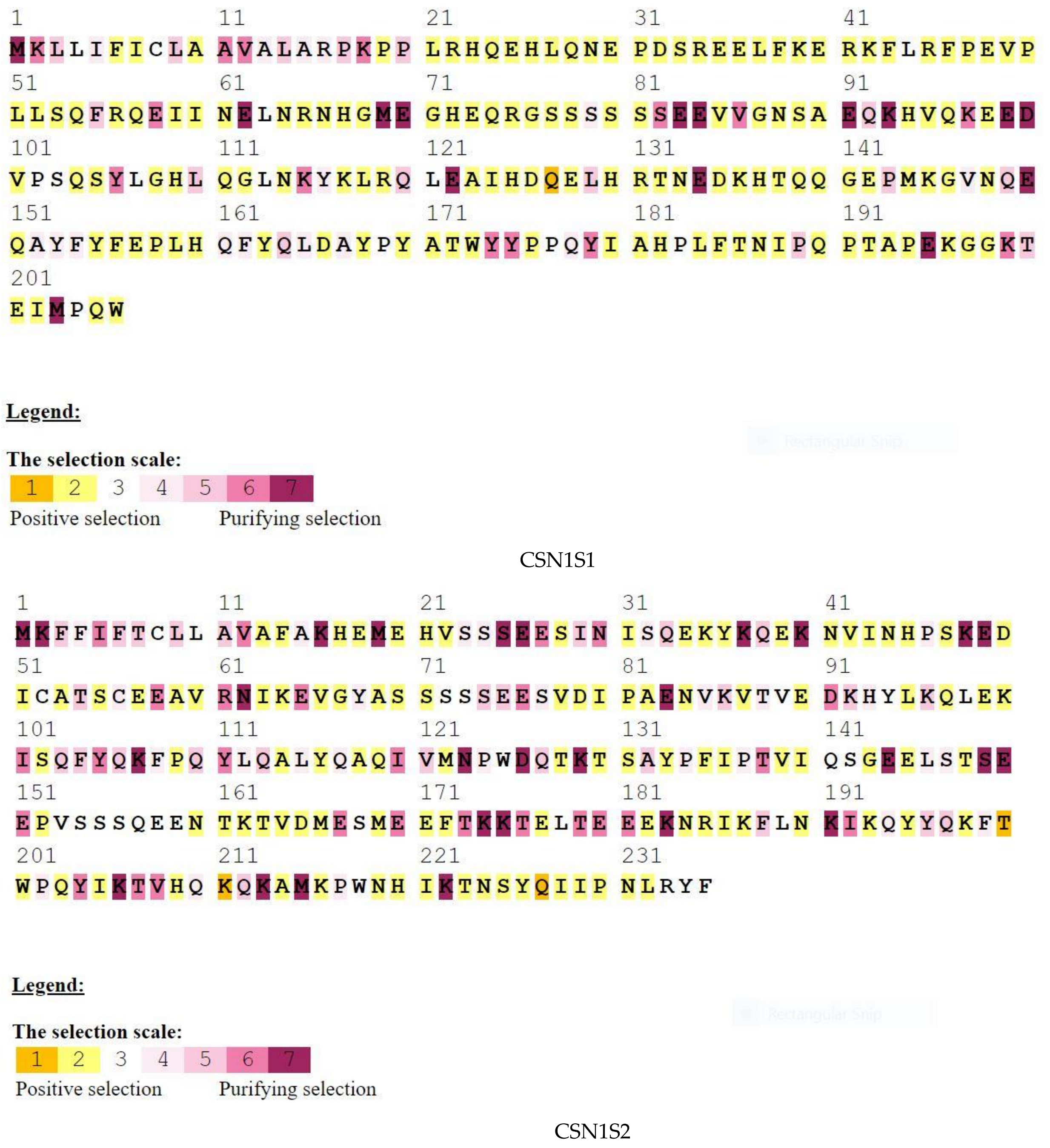
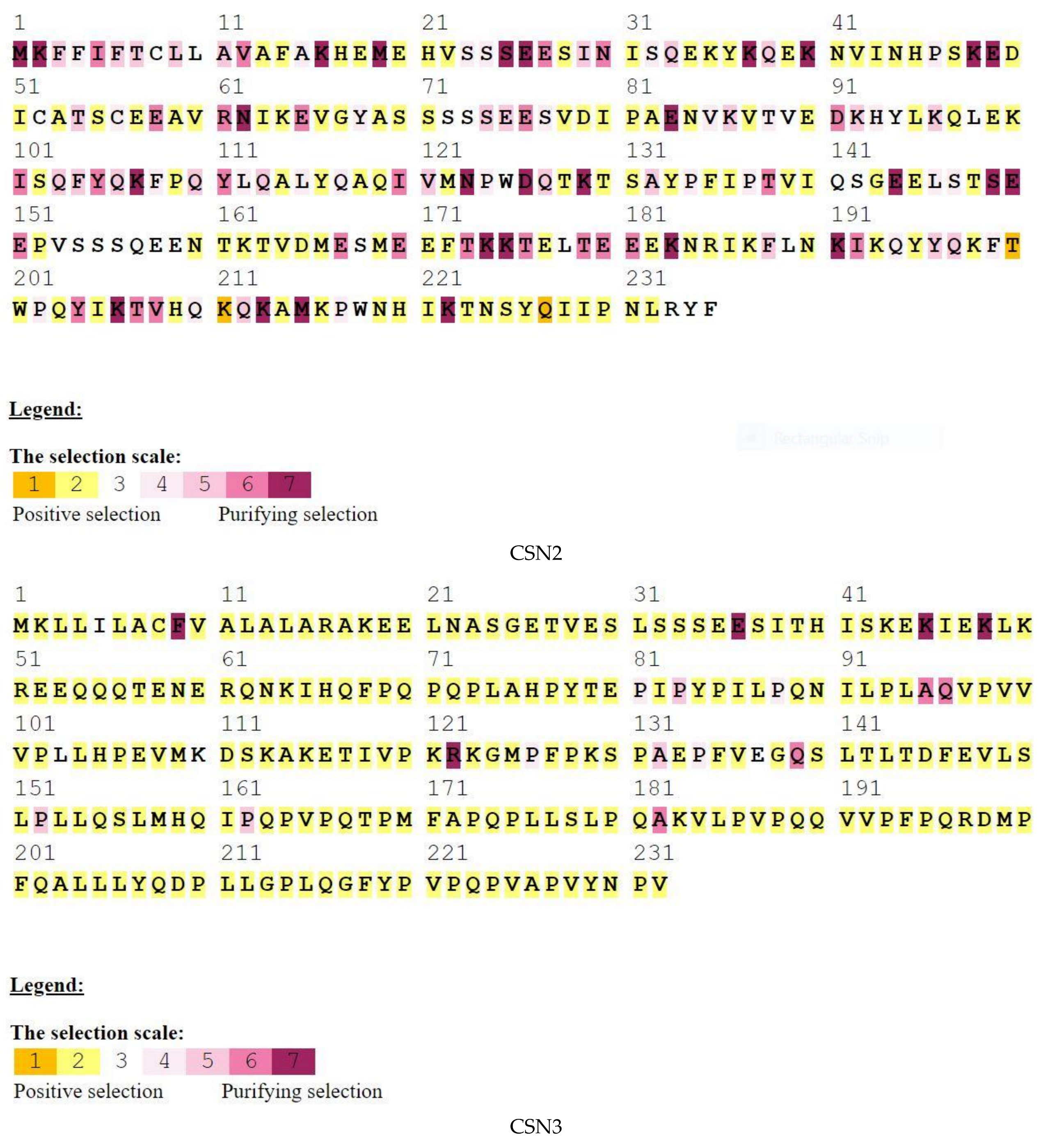
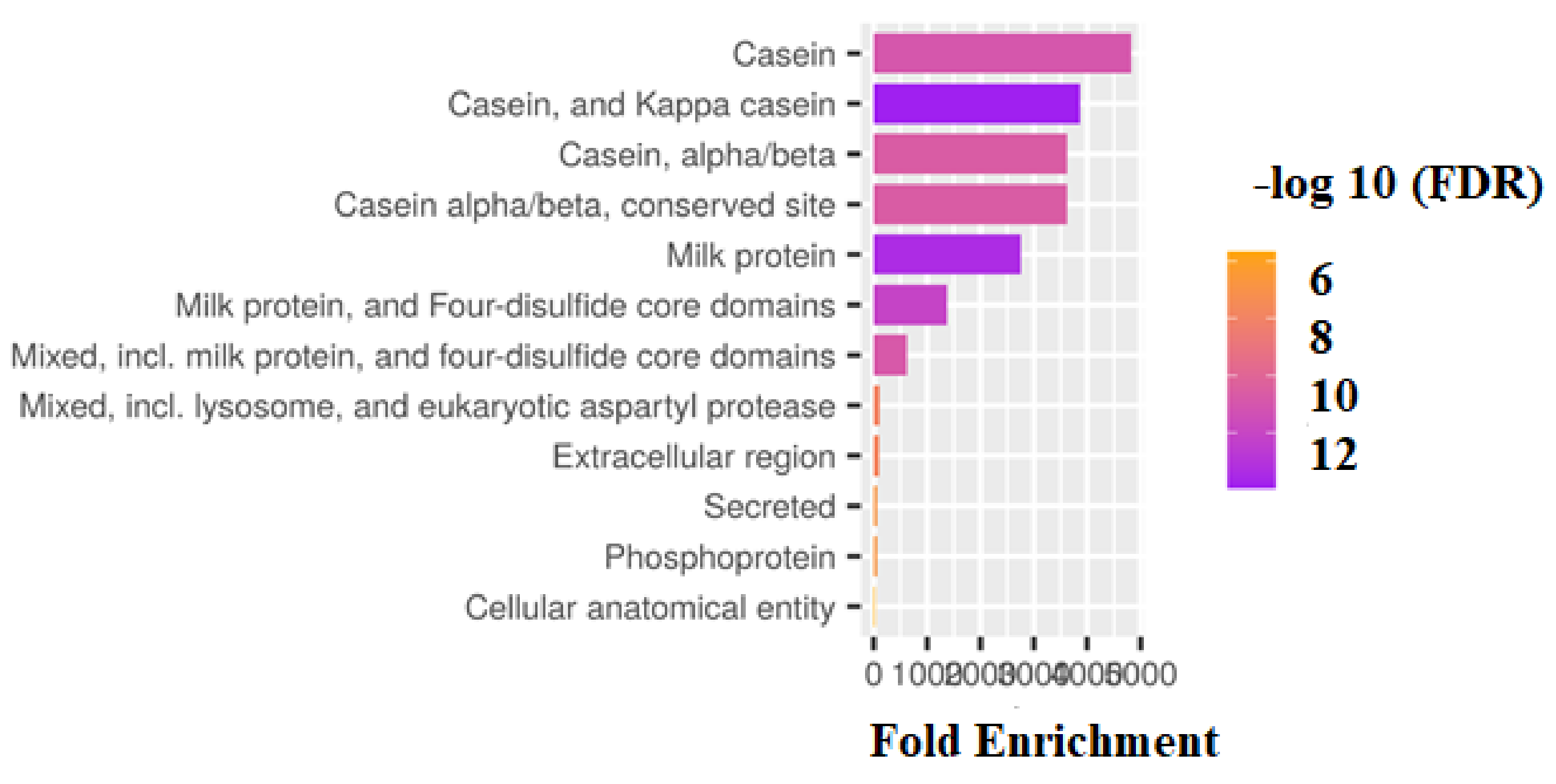
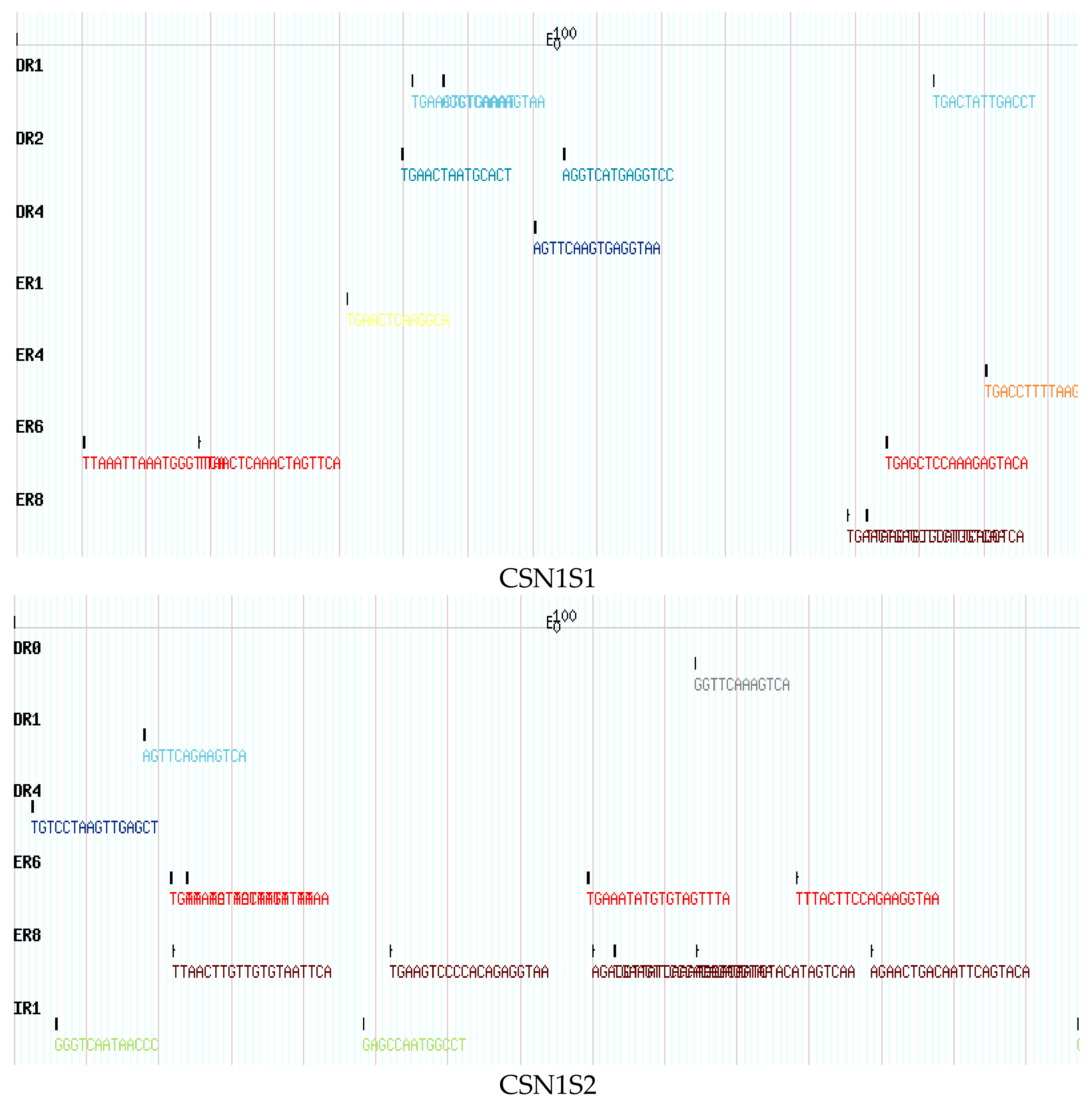
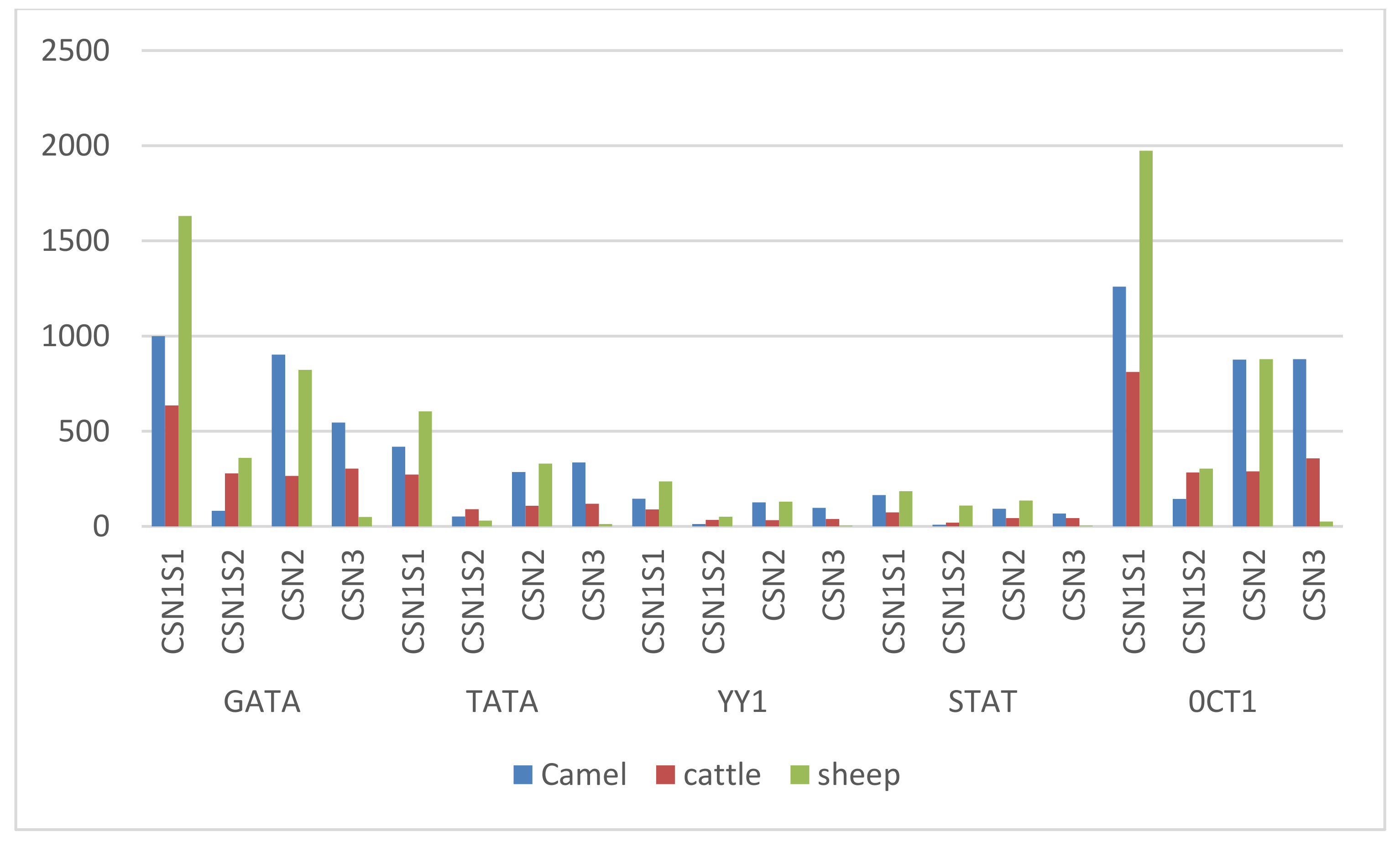
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rijnkels, M.; Elnitski, L.; Miller, W.; Rosen, J.M. Multispecies Comparative Analysis of a Mammalian-Specific Genomic Domain Encoding Secretory Proteins. Genomics 2003, 82, 417–432. [Google Scholar] [CrossRef]
- Wei, L.; Liu, Y.; Dubchak, I.; Shon, J.; Park, J. Comparative Genomics Approaches to Study Organism Similarities and Differences. J. Biomed. Inform. 2002, 35, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauciullo, A.; Shuiep, E.T.; Ogah, M.D.; Cosenza, G.; Di Stasio, L.; Erhardt, G. Casein Gene Cluster in Camelids: Comparative Genome Analysis and New Findings on Haplotype Variability and Physical Mapping. Front. Genet. 2019, 10, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faye, B.; Konuspayeva, G. The Sustainability Challenge to the Dairy Sector–The Growing Importance of Non-Cattle Milk Production Worldwide. Int. Dairy J. 2012, 24, 50–56. [Google Scholar] [CrossRef]
- Habib, H.M.; Ibrahim, W.H.; Schneider-Stock, R.; Hassan, H.M. Camel Milk Lactoferrin Reduces the Proliferation of Colorectal Cancer Cells and Exerts Antioxidant and DNA Damage Inhibitory Activities. Food Chem. 2013, 141, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Mal, G.; Sena, D.S.; Jain, V.K.; Sahani, M.S. Therapeutic Value of Camel Milk as a Nutritional Supplement for Multiple Drug Resistant (MDR) Tuberculosis Patients. Isr. J. Vet. Med. 2006, 61, 88. [Google Scholar]
- Kim, D.-H.; Jeong, D.-K.; Kim, H.; Chon, J.-W.; Lim, H.-W.; Chang, H.-S.; Park, J.-H.; Kim, Y.-J.; Kang, I.-B.; Song, K.-Y.; et al. Manufacture of Functional Koumiss Supplemented with Cichorium Intybus L. (Chicory) Extract-Preliminary Study. J. Dairy Sci. Biotechnol. 2017, 35, 1–7. [Google Scholar] [CrossRef]
- Rao, M.B.; Gupta, R.C.; Dastur, N.N. Camels’ Milk and Milk Products. Indian J. Dairy Sci. 1970, 23, 71–78. [Google Scholar]
- Shalash, M.R. Utilization of Camel Meat and Milk in Human Nourishment. In Proceedings of the Workshop on Camels, Khartoum, Sudan, 18–20 December 1979; pp. 295–306. [Google Scholar]
- Yagil, R.; Etzion, Z. Milk Yield of Camels (Camelus Dromedarius) in Drought Areas. Comp. Biochem. Physiol. Part A Physiol. 1980, 67, 207–209. [Google Scholar] [CrossRef]
- Fan, X.; Gao, S.; Fu, L.; Qiu, L.; Miao, Y. Polymorphism and Molecular Characteristics of the CSN1S2 Gene in River and Swamp Buffalo. Arch. Anim. Breed. 2020, 63, 345–354. [Google Scholar] [CrossRef]
- Pauciullo, A.; Erhardt, G. Molecular Characterization of the Llamas (Lama Glama) Casein Cluster Genes Transcripts (CSN1S1, CSN2, CSN1S2, CSN3) and Regulatory Regions. PLoS ONE 2015, 10, e0124963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elagamy, E.I.; Ruppanner, R.; Ismail, A.; Champagne, C.P.; Assaf, R. Purification and Characterization of Lactoferrin, Lactoperoxidase, Lysozyme and Immunoglobulins from Camel’s Milk. Int. Dairy J. 1996, 6, 129–145. [Google Scholar] [CrossRef]
- El-Agamy, E.S.I. Camel Milk. In Handbook of Milk of Non-Bovine Mammals, 2nd ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2017; pp. 409–480. [Google Scholar] [CrossRef]
- Amandykova, M.; Dossybayev, K.; Mussayeva, A.; Bekmanov, B.; Saitou, N. Comparative Analysis of the Polymorphism of the Casein Genes in Camels Bred in Kazakhstan. Diversity 2022, 14, 285. [Google Scholar] [CrossRef]
- Amigo, L.; Recio, I.; Ramos, M. Genetic Polymorphism of Ovine Milk Proteins: Its Influence on Technological Properties of Milk—A Review. Int. Dairy J. 2000, 10, 135–149. [Google Scholar] [CrossRef]
- Alim, N.; Fondrini, F.; Bonizzi, I.; Feligini, M.; Enne, G. Characterization of Casein Fractions from Algerian Dromedary (Camelus Dromedarius) Milk. Pakistan J. Nutr. 2005, 4, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, D.J.; Winzeler, E.A. Genomics, Gene Expression and DNA Arrays. Nature 2000, 405, 827–836. [Google Scholar] [CrossRef]
- Erhardt, G.; Shuiep, E.T.S.; Lisson, M.; Weimann, C.; Wang, Z.; El Zubeir, I.E.Y.M.; Pauciullo, A. Alpha S1-Casein Polymorphisms in Camel (Camelus Dromedarius) and Descriptions of Biological Active Peptides and Allergenic Epitopes. Trop. Anim. Health Prod. 2016, 48, 879–887. [Google Scholar] [CrossRef]
- Bulyk, M.L. Computational prediction of transcription-factor binding site locations. Genome Biol. 2003, 5, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soyudal, B.; Ardicli, S.; Samli, H.; Dincel, D.; Balci, F. Association of Polymorphisms in the CSN2, CSN3, LGB and LALBA Genes with Milk Production Traits in Holstein Cows Raised in Turkey. J. Hell. Vet. Med. Soc. 2019, 69, 1271–1282. [Google Scholar] [CrossRef] [Green Version]
- Ur Rehman, S.; Feng, T.; Wu, S.; Luo, X.; Lei, A.; Luobu, B.; Hassan, F.U.; Liu, Q. Comparative Genomics, Evolutionary and Gene Regulatory Regions Analysis of Casein Gene Family in Bubalus Bubalis. Front. Genet. 2021, 12, 662609. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Akopyan, G.; Garban, H.; Bonavida, B. Transcription Factor YY1: Structure, Function, and Therapeutic Implications in Cancer Biology. Oncogene 2006, 25, 1125–1142. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kawasaki, K.; Lafont, A.-G.; Sire, J.-Y. The Evolution of Milk Casein Genes from Tooth Genes before the Origin of Mammals. Mol. Biol. Evol. 2011, 28, 2053–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, P.; Kuscu, C.; Dutta, A. Biogenesis and Function of Transfer RNA-Related Fragments (TRFs). Trends Biochem. Sci. 2016, 41, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Adar, S.; Selby, C.P.; Lieb, J.D.; Sancar, A. Genome-Wide Analysis of Human Global and Transcription-Coupled Excision Repair of UV Damage at Single-Nucleotide Resolution. Genes Dev. 2015, 29, 948–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and Analyzing DNA and Protein Sequence Motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Stern, A.; Doron-Faigenboim, A.; Erez, E.; Martz, E.; Bacharach, E.; Pupko, T. Selecton 2007: Advanced Models for Detecting Positive and Purifying Selection Using a Bayesian Inference Approach. Nucleic Acids Res. 2007, 35, W506–W511. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.X.; Jung, D.; Jung, D.; Yao, R. ShinyGO: A Graphical Gene-Set Enrichment Tool for Animals and Plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, S. Promoter2.0: For the Recognition of PolII Promoter Sequences. Bioinformatics 1999, 15, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, T.; Takagi, T. Estimating Transcription Factor Bindability on DNA. Bioinformatics 1999, 15, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.W.; Johnson, P.F.; Hennighausen, L.; Sterneck, E. The C/EBPβ Transcription Factor Regulates Epithelial Cell Proliferation and Differentiation in the Mammary Gland. Genes Dev. 1998, 12, 1907–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennighausen, L.; Robinson, G.W. Think Globally, Act Locally: The Making of a Mouse Mammary Gland. Genes Dev. 1998, 12, 449–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, J.M.; Wyszomierski, S.L.; Hadsell, D. Regulation of Milk Protein Gene Expression. Annu. Rev. Nutr. 1999, 19, 407–436. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, W.C.; Whiting, M.; Wheeler, Q.D.; Carpenter, J.M. The Phylogeny of the Extant Hexapod Orders. Cladistics 2001, 17, 113–169. [Google Scholar] [CrossRef]
- Tomic, S.; Chughtai, N.; Ali, S. SOCS-1,-2,-3: Selective Targets and Functions Downstream of the Prolactin Receptor. Mol. Cell. Endocrinol. 1999, 158, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.S.; Patel, S.J.; Patel, N.R.; Chaudhary, G. V Importance of Camel Milk-An Alternative Dairy Food. J. Livest. Sci. 2016, 7, 19–25. [Google Scholar]
- Luo, X.; Zhou, Y.; Zhang, B.; Zhang, Y.; Wang, X.; Feng, T.; Li, Z.; Cui, K.; Wang, Z.; Luo, C. Understanding Divergent Domestication Traits from the Whole-Genome Sequencing of Swamp-and River-Buffalo Populations. Natl. Sci. Rev. 2020, 7, 686–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijnkels, M. Multispecies Comparison of the Casein Gene Loci and Evolution of Casein Gene Family. J. Mammary Gland. Biol. Neoplasia 2002, 7, 327–345. [Google Scholar] [CrossRef]
- Madende, M.; Osthoff, G. Comparative Genomics of Casein Genes. J. Dairy Res. 2019, 86, 323–330. [Google Scholar] [CrossRef]
- Poth, A.G.; Deeth, H.C.; Alewood, P.F.; Holland, J.W. Analysis of the Human Casein Phosphoproteome by 2-D Electrophoresis and MALDI-TOF/TOF MS Reveals New Phosphoforms. J. Proteome Res. 2008, 7, 5017–5027. [Google Scholar] [CrossRef] [PubMed]
- Madende, M.; Kemp, G.; Stoychev, S.; Osthoff, G. Characterisation of African Elephant Beta Casein and Its Relevance to the Chemistry of Caseins and Casein Micelles. Int. Dairy J. 2018, 85, 112–120. [Google Scholar] [CrossRef]
- Guan, D.; Mármol-Sánchez, E.; Cardoso, T.F.; Such, X.; Landi, V.; Tawari, N.R.; Amills, M. Genomic Analysis of the Origins of Extant Casein Variation in Goats. J. Dairy Sci. 2019, 102, 5230–5241. [Google Scholar] [CrossRef] [PubMed]
- Caroli, A.; Chiatti, F.; Chessa, S.; Rignanese, D.; Bolla, P.; Pagnacco, G. Focusing on the Goat Casein Complex. J. Dairy Sci. 2006, 89, 3178–3187. [Google Scholar] [CrossRef] [Green Version]
- Caroli, A.M.; Chessa, S.; Erhardt, G.J. Invited Review: Milk Protein Polymorphisms in Cattle: Effect on Animal Breeding and Human Nutrition. J. Dairy Sci. 2009, 92, 5335–5352. [Google Scholar] [CrossRef] [Green Version]
- Ramunno, L.; Cosenza, G.; Rando, A.; Pauciullo, A.; Illario, R.; Gallo, D.; Di Berardino, D.; Masina, P. Comparative Analysis of Gene Sequence of Goat CSN1S1 F and N Alleles and Characterization of CSN1S1 Transcript Variants in Mammary Gland. Gene 2005, 345, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Luigi-Sierra, M.G.; Mármol-Sánchez, E.; Amills, M. Comparing the Diversity of the Casein Genes in the Asian Mouflon and Domestic Sheep. Anim. Genet. 2020, 51, 470–475. [Google Scholar] [CrossRef]
- Inostroza, M.G.P.; González, F.J.N.; Landi, V.; Jurado, J.M.L.; Bermejo, J.V.D.; Fernández Álvarez, J.; Martínez Martínez, M.D.A. Bayesian Analysis of the Association between Casein Complex Haplotype Variants and Milk Yield, Composition, and Curve Shape Parameters in Murciano-Granadina Goats. Animals 2020, 10, 1845. [Google Scholar] [CrossRef]
- Kappeler, S.; Farah, Z.; Puhan, Z. Sequence Analysis of Camelus Dromedarius Milk Caseins. J. Dairy Res. 1998, 65, 209–222. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Mal, G.; Kumar, D.; Patil, N.V.; Pathak, K.M.L. Camel Milk: An Important Natural Adjuvant. Agric. Res. 2017, 6, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Pauciullo, A.; Giambra, I.J.; Iannuzzi, L.; Erhardt, G. The β-Casein in Camels: Molecular Characterization of the CSN2 Gene, Promoter Analysis and Genetic Variability. Gene 2014, 547, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Stormo, G.D. Modeling the Specificity of Protein-DNA Interactions. Quant. Biol. 2013, 1, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Guigó, R.; Knudsen, S.; Drake, N.; Smith, T. Prediction of Gene Structure. J. Mol. Biol. 1992, 226, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.I.; Afzal, G.; Sadia, S.; Haider, G.; Ahmed, S.; Saeed, S.; Chen, J. Structural and Evolutionary Adaptations of Nei-Like DNA Glycosylases Proteins Involved in Base Excision Repair of Oxidative DNA Damage in Vertebrates. Oxid. Med. Cell. Longev. 2022, 2022, 1144387. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Al-Shahrour, F.; Díaz-Uriarte, R.; Dopazo, J. FatiGO: A Web Tool for Finding Significant Associations of Gene Ontology Terms with Groups of Genes. Bioinformatics 2004, 20, 578–580. [Google Scholar] [CrossRef] [Green Version]
- Swaisgood, H.E.; Brunner, J.R. The Caseins. Crit. Rev. Food Sci. Nutr. 1973, 3, 375–414. [Google Scholar] [CrossRef]
- Teshima, K.M.; Przeworski, M. Directional Positive Selection on an Allele of Arbitrary Dominance. Genetics 2006, 172, 713–718. [Google Scholar] [CrossRef] [Green Version]
- Holt, C.; Raynes, J.K.; Carver, J.A. Sequence Characteristics Responsible for Protein-Protein Interactions in the Intrinsically Disordered Regions of Caseins, Amelogenins, and Small Heat-Shock Proteins. Biopolymers 2019, 110, e23319. [Google Scholar] [CrossRef] [PubMed]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited Review: Caseins and the Casein Micelle: Their Biological Functions, Structures, and Behavior in Foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef] [Green Version]
- Gallego, M.I.; Binart, N.; Robinson, G.W.; Okagaki, R.; Coschigano, K.T.; Perry, J.; Kopchick, J.J.; Oka, T.; Kelly, P.A.; Hennighausen, L. Prolactin, Growth Hormone, and Epidermal Growth Factor Activate Stat5 in Different Compartments of Mammary Tissue and Exert Different and Overlapping Developmental Effects. Dev. Biol. 2001, 229, 163–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurisaki, K.; Kurisaki, A.; Valcourt, U.; Terentiev, A.A.; Pardali, K.; Ten Dijke, P.; Heldin, C.-H.; Ericsson, J.; Moustakas, A. Nuclear Factor YY1 Inhibits Transforming Growth Factor β-and Bone Morphogenetic Protein-Induced Cell Differentiation. Mol. Cell. Biol. 2003, 23, 4494–4510. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Sequence | Length | Pfam Domain |
|---|---|---|---|
| 1 | MKLLILTCLVAVALARPKEEL | 21 | ---- |
| 2 | TEVFTKKTKLTEEEKNRLNFLKKISQYYQKFAWPQYLKTVYQYQKAMKPW | 50 | Casein |
| 3 | KFPQYLLPLYQGPIVVPPWDQ | 21 | ---- |
| 4 | AGEEEESLSSSSEEIVHISKEQKKIQKED | 29 | ---- |
| 5 | EQLHSMKEGNHAQQKEPMIGVNQELAYFYPELFRQFYQLDAYPSGAWYY | 49 | Casein |
| 6 | EVMGVSKVKETIVPKHKEMPFPKYPVEPFTESQSLTLTDVE | 41 | ----- |
| 7 | LPLLQSWMHQPPQPLPPTPMFPPQSLLSLSQAKVLPVPQKAVP | 43 | Casein |
| 8 | MKSFFLVVTILALTLPFLGAQEQNQEQPIRCEKDERFFNDKIAKYIPIQY | 50 | K-casein |
| 9 | PRNALPFQAIPLKEQPDKEEINGLNTIIG | 29 | ---- |
| 10 | QQQTEDELQDKIHPFPQPQSLVYPYTGPI | 29 | ---- |
| Species | Gene | Chromosome | MW(Da) | AA | pI | AI | II | Gravy |
|---|---|---|---|---|---|---|---|---|
| Camelus ferus | CSN1S1 | 2 | 26,861.4 | 230 | 4.96 | 84.3 | 64.07 | −0.661 |
| Camelus ferus | CSN1S2 | 2 | 22,964.1 | 193 | 6 | 67.62 | 58.11 | −0.661 |
| Camelus ferus | CSN2 | 2 | 26,174.81 | 232 | 5.43 | 101.59 | 97.41 | −0.147 |
| Camelus ferus | CSN3 | 2 | 20,373.53 | 182 | 8.44 | 90.49 | 44.72 | −0.152 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parveen, S.; Zhu, P.; Shafique, L.; Lan, H.; Xu, D.; Ashraf, S.; Ashraf, S.; Sherazi, M.; Liu, Q. Molecular Characterization and Phylogenetic Analysis of Casein Gene Family in Camelus ferus. Genes 2023, 14, 256. https://doi.org/10.3390/genes14020256
Parveen S, Zhu P, Shafique L, Lan H, Xu D, Ashraf S, Ashraf S, Sherazi M, Liu Q. Molecular Characterization and Phylogenetic Analysis of Casein Gene Family in Camelus ferus. Genes. 2023; 14(2):256. https://doi.org/10.3390/genes14020256
Chicago/Turabian StyleParveen, Shakeela, Peng Zhu, Laiba Shafique, Hong Lan, Dingyun Xu, Sana Ashraf, Saba Ashraf, Maryam Sherazi, and Qingyou Liu. 2023. "Molecular Characterization and Phylogenetic Analysis of Casein Gene Family in Camelus ferus" Genes 14, no. 2: 256. https://doi.org/10.3390/genes14020256