

Inferring Causalities of Environmental and Genetic Factors for Differential Somatic Cell Count and Mastitis Pathogens in Dairy Cows Using Structural Equation Modelling

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Farms, Animals and Sampling
2.3. Genome-Wide Associations
2.4. Structural Equation Model
3. Results
3.1. Overall Structural Equation Model Evaluation
3.2. Latent Variable Intramammary Infection
3.3. Latent Variable Production
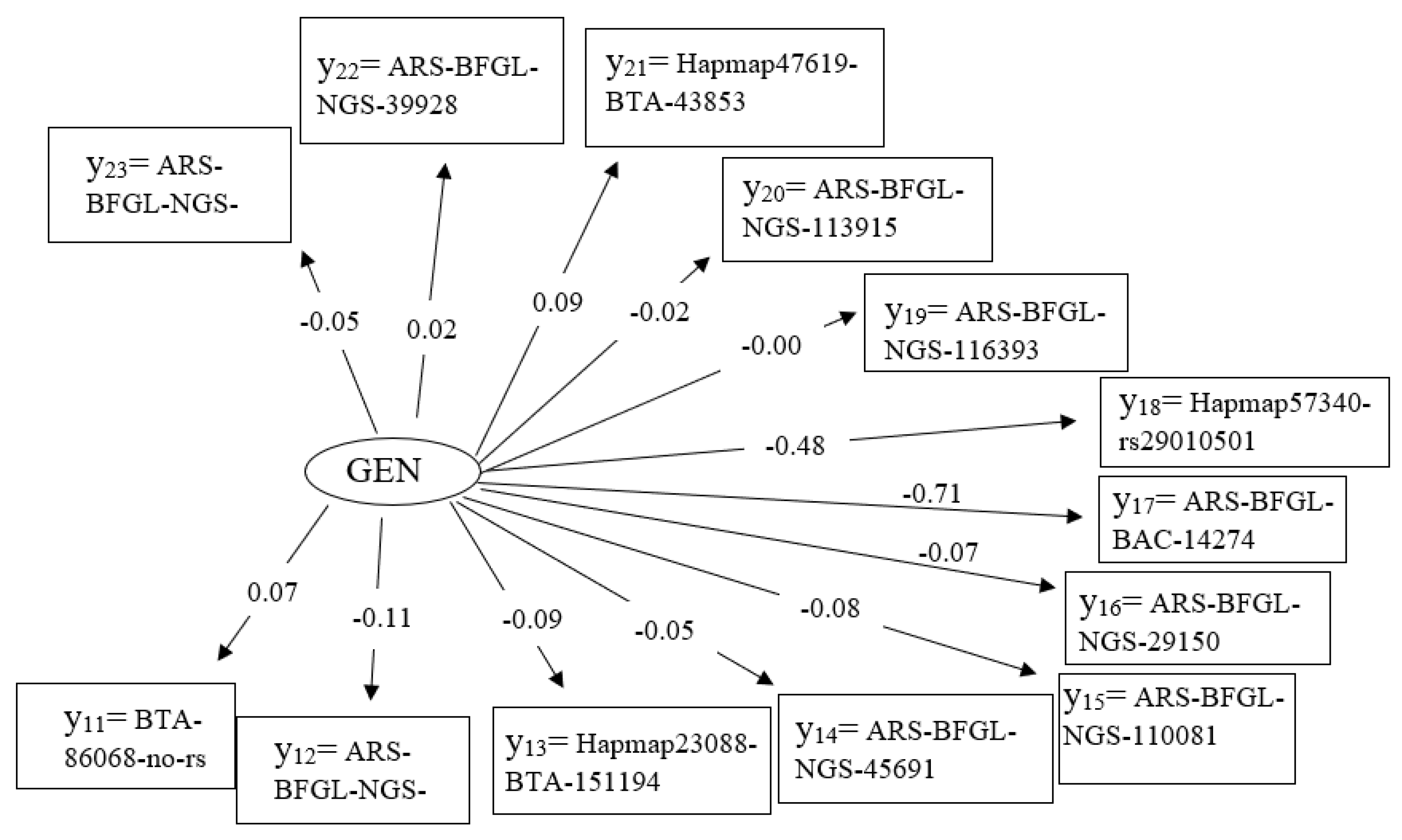
3.4. Latent Variable Genetic
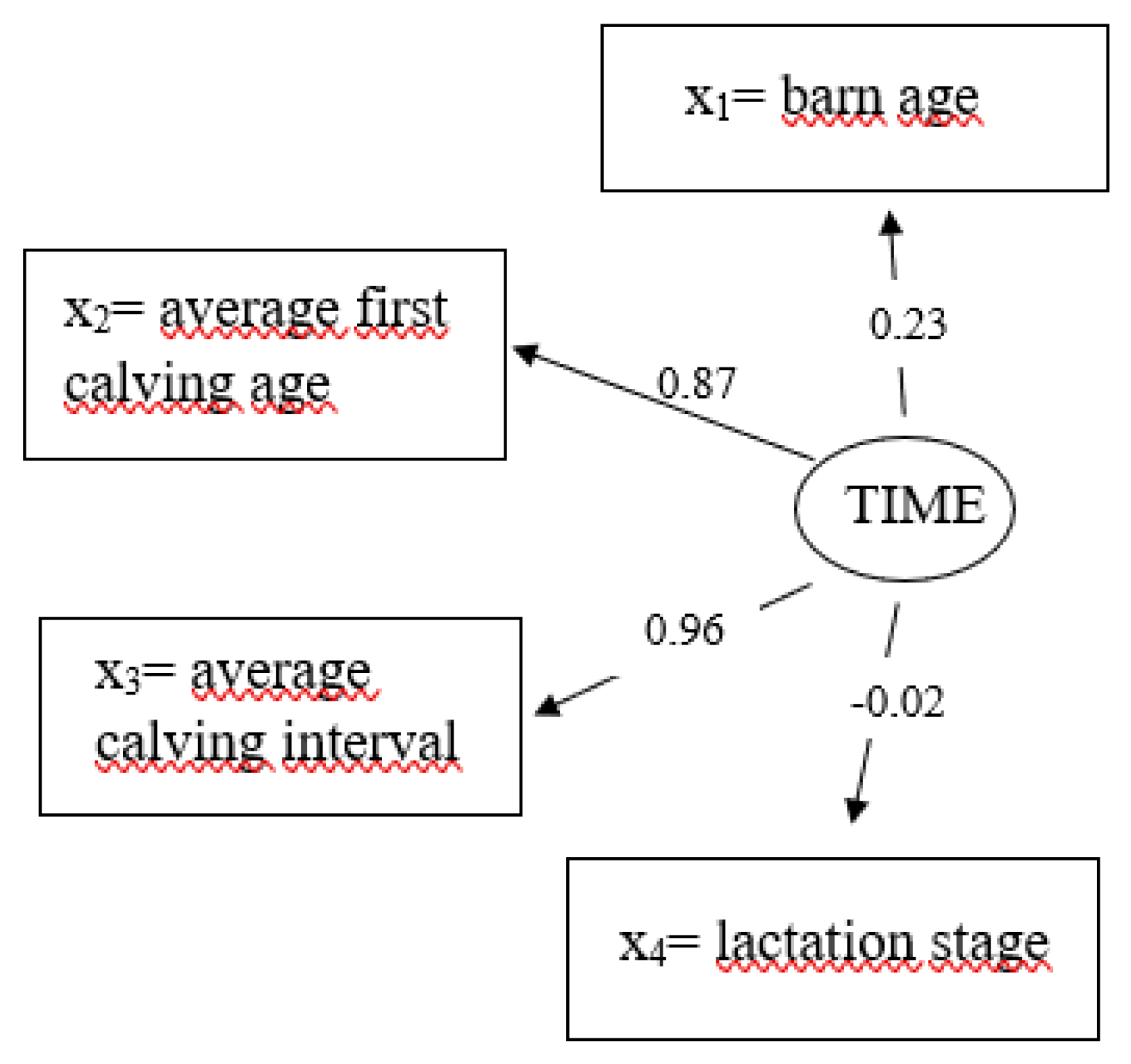
3.5. Latent Variable Time
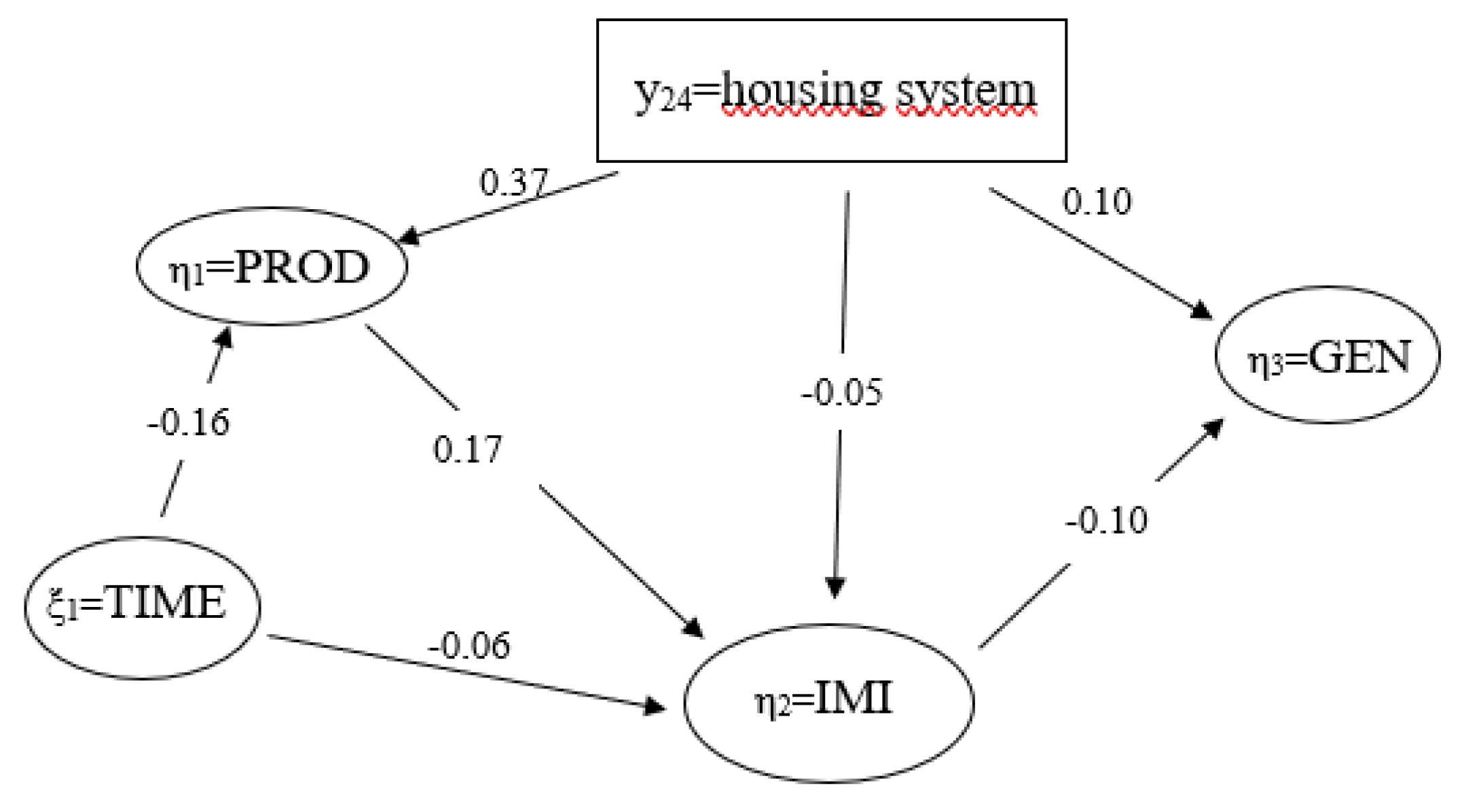
3.6. Relationships among Latent Variables
4. Discussion
4.1. Manifest Variables on Intramammary Infection and Production
4.2. Genetic Influence in the Structural Equation Model
4.3. Overall Structural Equation Model Evaluations, Limitations and Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fourichon, C.; Seegers, H.; Beaudeau, F.; Verfaille, L.; Bareille, N. Health-control costs in dairy farming systems in western France. Livest. Prod. Sci. 2001, 68, 141–156. [Google Scholar] [CrossRef]
- Schukken, Y.H.; González, R.N.; Tikofsky, L.L.; Schulte, H.F.; Santisteban, C.G.; Welcome, F.L.; Bennett, G.J.; Zurakowski, M.J.; Zadoks, R.N. CNS mastitis: Nothing to worry about? Vet. Microbiol. 2009, 134, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Detilleux, J.; Theron, L.; Beduin, J.-M.; Hanzen, C. A structural equation model to evaluate direct and indirect factors associated with a latent measure of mastitis in Belgian dairy herds. Prev. Vet. Med. 2012, 107, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Leso, L.; Barbari, M.; Lopes, M.A.; Damasceno, F.A.; Galama, P.; Taraba, J.L.; Kuipers, A. Invited review: Compost-bedded pack barns for dairy cows. J. Dairy Sci. 2020, 103, 1072–1099. [Google Scholar] [CrossRef] [PubMed]
- Barberg, A.E.; Endres, M.I.; Salfer, J.A.; Reneau, J.K. Performance and welfare of dairy cows in an alternative housing system in Minnesota. J. Dairy Sci. 2007, 90, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.; Yin, T.; Brügemann, K.; Engel, P.; Weimann, C.; Schlez, K.; König, S. Genome-Wide Associations for Microscopic Differential Somatic Cell Count and Specific Mastitis Pathogens in Holstein Cows in Compost-Bedded Pack and Cubicle Farming Systems. Animals 2021, 11, 1839. [Google Scholar] [CrossRef] [PubMed]
- Pighetti, G.M.; Elliott, A.A. Gene polymorphisms: The keys for marker assisted selection and unraveling core regulatory pathways for mastitis resistance. J. Mammary Gland. Biol. Neoplasia 2011, 16, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Malek dos Reis, C.B.; Barreiro, J.R.; Mestieri, L.; Porcionato, M.A.d.F.; dos Santos, M.V. Effect of somatic cell count and mastitis pathogens on milk composition in Gyr cows. BMC Vet. Res. 2013, 9, 67. [Google Scholar] [CrossRef]
- França, M.M.; Del Valle, T.A.; Campana, M.; Veronese, L.P.; Nascimento, G.; Morais, J.P.G. Agentes causadores de mastite e relações entre a CCS com a produção e com a composição do leite em vacas leiteiras. Arch. Zootec. 2017, 66, 45–49. [Google Scholar] [CrossRef]
- Klarmann, M. Methodische Problemfelder der Erfolgsfaktorenforschung: Bestandsaufnahme und Empirische Analysen; Gabler: Wiesbaden, Germany, 2008; ISBN 9783834914071. [Google Scholar]
- Gana, K.; Broc, G. (Eds.) Structural Equation Modeling with Lavaan; Wiley-ISTE: London, UK; Hoboken, NJ, USA, 2019; ISBN 9781119579038. [Google Scholar]
- Hair, J.F., Jr.; Sarstedt, M.; Hopkins, L.; Kuppelwieser, V.G. Partial least squares structural equation modeling (PLS-SEM). Eur. Bus. Rev. 2014, 26, 106–121. [Google Scholar] [CrossRef]
- de los Campos, G.; Gianola, D.; Heringstad, B. A structural equation model for describing relationships between somatic cell score and milk yield in first-lactation dairy cows. J. Dairy Sci. 2006, 89, 4445–4455. [Google Scholar] [CrossRef]
- Wu, X.-L.; Heringstad, B.; Gianola, D. Exploration of lagged relationships between mastitis and milk yield in dairy cows using a Bayesian structural equation Gaussian-threshold model. Genet. Sel. Evol. 2008, 40, 333–357. [Google Scholar] [CrossRef]
- Casal, J.; Learte, P.; Torre, E. A path model of factors influencing bovine leukemia virus transmission between cattle herds. Prev. Vet. Med. 1990, 10, 47–61. [Google Scholar] [CrossRef]
- Detilleux, J.; Theron, L.; Duprez, J.-N.; Reding, E.; Humblet, M.-F.; Planchon, V.; Delfosse, C.; Bertozzi, C.; Mainil, J.; Hanzen, C. Structural equation models to estimate risk of infection and tolerance to bovine mastitis. Genet. Sel. Evol. 2013, 45, 6. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Penedo, I.; Ouweltjes, W.; Ofner-Schröck, E.; Brügemann, K.; Emanuelson, U. Symposium review: Animal welfare in free-walk systems in Europe. J. Dairy Sci. 2020, 103, 5773–5782. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.; Brügemann, K.; Yin, T.; Engel, P.; Weimann, C.; Schlez, K.; König, S. Microscopic differential cell count and specific mastitis pathogens in cow milk from compost-bedded pack barns and cubicle barns. J. Dairy Res. 2021, 88, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Deutsche Veterinärmedizinische Gesellschaft. Leitlinien zur Entnahme von Milchproben unter antiseptischen Bedingungen und Leitlinien zur Isolierung und Identifizierung von Mastitiserregern; Dt. Veterinärmed. Ges., Sachverständigenausschuss Subklinische Mastitis: Gießen, Germany, 2000; ISBN 3930511819. [Google Scholar]
- Ariznabarreta, A.; Gonzalo, C.; San Primitivo, F. Microbiological quality and somatic cell count of ewe milk with special reference to staphylococci. J. Dairy Sci. 2002, 85, 1370–1375. [Google Scholar] [CrossRef] [PubMed]
- Zecconi, A.; Dell’Orco, F.; Vairani, D.; Rizzi, N.; Cipolla, M.; Zanini, L. Differential Somatic Cell Count as a Marker for Changes of Milk Composition in Cows with Very Low Somatic Cell Count. Animals 2020, 10, 604. [Google Scholar] [CrossRef]
- Schwarz, D.; Santschi, D.E.; Durocher, J.; Lefebvre, D.M. Evaluation of the new differential somatic cell count parameter as a rapid and inexpensive supplementary tool for udder health management through regular milk recording. Prev. Vet. Med. 2020, 181, 105079. [Google Scholar] [CrossRef]
- Kirkeby, C.; Toft, N.; Schwarz, D.; Farre, M.; Nielsen, S.S.; Zervens, L.; Hechinger, S.; Halasa, T. Differential somatic cell count as an additional indicator for intramammary infections in dairy cows. J. Dairy Sci. 2020, 103, 1759–1775. [Google Scholar] [CrossRef]
- Sarikaya, H.; Werner-Misof, C.; Atzkern, M.; Bruckmaier, R.M. Distribution of leucocyte populations, and milk composition, in milk fractions of healthy quarters in dairy cows. J. Dairy Res. 2005, 72, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Pappenheim, A. Zur Blutzellfärbung im klinischen Bluttrockenpräparat und zur histologischen Schnittpräparatfärbung der hämatopoetischen Gewebe nach meinen Methoden. Folia Haematologica. 1912, 13, 337–344. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; Bakker, P.I.W.; de Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G.; et al. Ensembl 2018. Nucleic Acids Res. 2018, 46, D754–D761. [Google Scholar] [CrossRef]
- Yang, J.; Benyamin, B.; McEvoy, B.P.; Gordon, S.; Henders, A.K.; Nyholt, D.R.; Madden, P.A.; Heath, A.C.; Martin, N.G.; Montgomery, G.W.; et al. Common SNPs explain a large proportion of the heritability for human height. Nat. Genet. 2010, 42, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Karunarathna, C.B.; Graham, J. 46th European Mathematical Genetics Meeting (EMGM) 2018, Cagliari, Italy, April 18–20, 2018: Abstracts. Hum. Hered. 2018, 83, 1–29. [Google Scholar] [CrossRef]
- Halli, K.; Vanvanhossou, S.F.; Bohlouli, M.; König, S.; Yin, T. Identification of candidate genes on the basis of SNP by time-lagged heat stress interactions for milk production traits in German Holstein cattle. PLoS ONE 2021, 16, e0258216. [Google Scholar] [CrossRef] [PubMed]
- Wald, A. Tests of statistical hypotheses concerning several parameters when the number of observations is large. Trans. Amer. Math. Soc. 1943, 54, 426–482. [Google Scholar] [CrossRef]
- ENSEMBL Genome Browser. Available online: http://www.ensembl.org/ (accessed on 30 April 2023).
- National Center for Biotchnology Information (NCBI). Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 30 April 2023).
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, D199–D205. [Google Scholar] [CrossRef]
- R Core Team (2019). Available online: https://www.r-project.org (accessed on 30 April 2023).
- Rosseel, Y. lavaan: An R Package for Structural Equation Modeling. J. Stat. Soft. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Chin, W.W. The partial least squares approach to structural equation modeling. In Modern Methods for Business Research; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 1998; pp. 295–336. [Google Scholar]
- Beaudeau, F.; Fourichon, C.; Seegers, H.; Bareille, N. Risk of clinical mastitis in dairy herds with a high proportion of low individual milk somatic-cell counts. Prev. Vet. Med. 2002, 53, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Paape, M.J.; Bannerman, D.D.; Zhao, X.; Lee, J.-W. The bovine neutrophil: Structure and function in blood and milk. Vet. Res. 2003, 34, 597–627. [Google Scholar] [CrossRef] [PubMed]
- Riggio, V.; Portolano, B.; Bovenhuis, H.; Bishop, S.C. Genetic parameters for somatic cell score according to udder infection status in Valle del Belice dairy sheep and impact of imperfect diagnosis of infection. Genet. Sel. Evol. 2010, 42, 30. [Google Scholar] [CrossRef] [PubMed]
- Dosogne, H.; Vangroenweghe, F.; Mehrzad, J.; Massart-Leën, A.M.; Burvenich, C. Differential leukocyte count method for bovine low somatic cell count milk. J. Dairy Sci. 2003, 86, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Diesterbeck, U.S.; König, S.; Brügemann, K.; Schlez, K.; Zschöck, M.; Wolter, W.; Czerny, C.-P. Microscopic differential cell counts in milk for the evaluation of inflammatory reactions in clinically healthy and subclinically infected bovine mammary glands. J. Dairy Res. 2011, 78, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Streicher, K.L. Mammary gland immunity and mastitis susceptibility. J. Mammary Gland Biol. Neoplasia 2002, 7, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Tenhagen, B.-A.; Köster, G.; Wallmann, J.; Heuwieser, W. Prevalence of mastitis pathogens and their resistance against anti-microbial agents in dairy cows in Brandenburg, Germany. J. Dairy Sci. 2006, 89, 2542–2551. [Google Scholar] [CrossRef]
- Piessens, V.; van Coillie, E.; Verbist, B.; Supré, K.; Braem, G.; van Nuffel, A.; de Vuyst, L.; Heyndrickx, M.; de Vliegher, S. Distribution of coagulase-negative Staphylococcus species from milk and environment of dairy cows differs between herds. J. Dairy Sci. 2011, 94, 2933–2944. [Google Scholar] [CrossRef]
- Craig, A.-L.; Gordon, A.W.; Hamill, G.; Ferris, C.P. Milk Composition and Production Efficiency within Feed-To-Yield Systems on Commercial Dairy Farms in Northern Ireland. Animals 2022, 12, 1771. [Google Scholar] [CrossRef]
- Sørensen, L.P.; Madsen, P.; Mark, T.; Lund, M.S. Genetic parameters for pathogen-specific mastitis resistance in Danish Holstein Cattle. Animal 2009, 3, 647–656. [Google Scholar] [CrossRef]
- Li, M.; Lu, G.; Hu, J.; Shen, X.; Ju, J.; Gao, Y.; Qu, L.; Xia, Y.; Chen, Y.; Bai, Y. EVA1A/TMEM166 Regulates Embryonic Neu-rogenesis by Autophagy. Stem Cell Rep. 2016, 6, 396–410. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Kan, S.; Liu, Z.; Lu, G.; Zhang, X.; Chen, Y.; Bai, Y. EVA1A inhibits GBM cell proliferation by inducing autophagy and apoptosis. Exp. Cell Res. 2017, 352, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Yang, X.; Yang, H.; Lv, M.; Sun, X.; Zhou, B. Exosomal miR-338-3p suppresses non-small-cell lung cancer cells metastasis by inhibiting CHL1 through the MAPK signaling pathway. Cell Death Dis. 2021, 12, 1030. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu, L.-l.; Zhao, T.; Wu, L.-y.; Wu, K.-w.; Schachner, M.; Xiao, Z.-C.; Fan, M. CHL1 negatively regulates the proliferation and neuronal differentiation of neural progenitor cells through activation of the ERK1/2 MAPK pathway. Mol. Cell. Neurosci. 2011, 46, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.Z.; Khan, A.; Xiao, J.; Ma, J.; Ma, Y.; Chen, T.; Shao, D.; Cao, Z. Overview of Research Development on the Role of NF-κB Signaling in Mastitis. Animals 2020, 10, 1625. [Google Scholar] [CrossRef] [PubMed]
- Ivemeyer, S.; Knierim, U.; Waiblinger, S. Effect of human-animal relationship and management on udder health in Swiss dairy herds. J. Dairy Sci. 2011, 94, 5890–5902. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.M.; El Agawany, A.A.A. Impact of age at first calving on reproduction, lactation, postpartum disorders and longevity in Holsteins under Egyptian circumstances. J. Vet. Med. Res. 2009, 19, 42–52. [Google Scholar] [CrossRef]
- Pirlo, G.; Miglior, F.; Speroni, M. Effect of age at first calving on production traits and on difference between milk yield returns and rearing costs in Italian Holsteins. J. Dairy Sci. 2000, 83, 603–608. [Google Scholar] [CrossRef]
- Eastham, N.T.; Coates, A.; Cripps, P.; Richardson, H.; Smith, R.; Oikonomou, G. Associations between age at first calving and subsequent lactation performance in UK Holstein and Holstein-Friesian dairy cows. PLoS ONE 2018, 13, e0197764. [Google Scholar] [CrossRef]
- Sawa, A.; Siatka, K.; Krężel-Czopek, S. Effect of Age at First Calving on First Lactation Milk Yield, Lifetime Milk Production and Longevity of Cows. Ann. Anim. Sci. 2019, 19, 189–200. [Google Scholar] [CrossRef]
- Drews, J.; Czycholl, I.; Junge, W.; Krieter, J. An evaluation of efficiency in dairy production using structural equation modelling. J. Agric. Sci. 2018, 156, 996–1004. [Google Scholar] [CrossRef]
- Urban, D.; Mayerl, J. SEM-Grundlagen. In Strukturgleichungsmodellierung: Ein Ratgeber Für Die Praxis; Urban, D., Mayerl, J., Eds.; Springer: Wiesbaden, Germany, 2014; pp. 25–81. ISBN 978-3-658-01918-1. [Google Scholar]
- Gernand, E.; König, S.; Kipp, C. Influence of on-farm measurements for heat stress indicators on dairy cow productivity, female fertility, and health. J. Dairy Sci. 2018, 102, 6660–6671. [Google Scholar] [CrossRef]
- König, S.; Wu, X.; Gianola, D.; Heringstad, B.; Simianer, H. Exploration of relationships between claw disorders and milk yield in Holstein cows via recursive linear and threshold Models. J. Dairy Sci. 2008, 81, 395–406. [Google Scholar] [CrossRef]
- Naderi, S.; Bohlouli, M.; Yin, T.; König, S. Genomic breeding values, SNP effects and gene identification for disease traits in cow training sets. Anim. Genet. 2018, 49, 178–192. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Udder Health Traits 1 | Mean | Min | Max | SD |
|---|---|---|---|---|
| Cell fractions (in relation to the total sum of all cell counts) Macrophages | 0.292 | 0.000 | 0.980 | 0.208 |
| Lymphocytes | 0.608 | 0.000 | 1.000 | 0.246 |
| PMN | 0.100 | 0.000 | 0.971 | 0.143 |
| Mastitis pathogens (in prevalences) Negative samples | 0.514 | 0.000 | 1.000 | 0.500 |
| Minor pathogens | 0.407 | 0.000 | 1.000 | 0.491 |
| Major pathogens | 0.030 | 0.000 | 1.000 | 0.171 |
| Trait | SNP | CHR | Position | p-Value SNP | SNP Located in a Gene | Gene Name |
|---|---|---|---|---|---|---|
| MAJOR | BTA-86068-no-rs (y = 11) M | 22 | 26048787 | 0.000000002563736 a | yes | CHL1 |
| BTA-86068-no-rs (y = 11) I | 22 | 26048787 | 0.0000004836461 a | yes | CHL1 | |
| ARS-BFGL-NGS-39928 (y = 22) M | 26 | 38508625 | 0.000002509339 a | - | - | |
| ARS-BFGL-BAC-14274 (y = 17) M | 11 | 44153677 | 0.000001737926 a | yes | EVA1A | |
| Hapmap57340-rs29010501 (y = 18) M | 11 | 44928962 | 0.000003947047 b | - | - | |
| Hapmap23088-BTA-151194 (y = 13) M | 1 | 152612216 | 0.000003844456 b | yes | HACL1 | |
| ARS-BFGL-NGS-60721 (y = 12) M | 1 | 35809354 | 0.000004774763 b | - | - | |
| Hapmap47619-BTA-43853 (y = 21) M | 18 | 4489809 | 0.0000008244985 b | - | - | |
| ARS-BFGL-NGS-110081 (y = 15) M | 4 | 41230144 | 0.000001551728 b | - | - | |
| ARS-BFGL-NGS-45691 (y = 14) M | 2 | 127889562 | 0.00000764746 b | - | - | |
| ARS-BFGL-NGS-29150 (y = 16) M | 5 | 108921269 | 0.000008442227 b | - | - | |
| ARS-BFGL-NGS-116393 (y = 19) M | 11 | 104186003 | 0.000002801936 b | yes | ABO | |
| ARS-BFGL-NGS-113915 (y = 20) M | 17 | 32550404 | 0.0000008220506 b | - | - | |
| MINOR | ARS-BFGL-NGS-112964 (y = 23) M | 14 | 68578807 | 0.000002708568 b | - | - |
| Latent Variables | Indicator Variables (Manifest Variables) | Range (Min–Max) or Groups | Mean | SD |
|---|---|---|---|---|
| η1 = Production (PROD) | y1 = fat content [in %] | 2.43–7.6 | 4.88 | 0.73 |
| y2 = protein content [in %] | 2.7–4.93 | 3.56 | 0.37 | |
| y3 = lactose content [in %] | 3.4–5.28 | 4.88 | 0.19 | |
| y4 = milk yield [in kg] | <25, 25.1–30, 30.1–35, 35.1–40, >40 | - | - | |
| η2 = Intramammary infection (IMI) | y5 = average somatic cell count of the herd | 5.05–5.58 | 5.30 | 0.14 |
| y6 = lymphocyte content [in %] | 0–100 | 61.00 | 0.25 | |
| y7 = PMN content [in %] | 0–97 | 9.80 | 0.14 | |
| y8 = somatic cell count of test day (the exact test day of our sampling) | 2.64–11.16 | 2.35 | 2.09 | |
| y9 = MAJOR | 0, 1 | - | - | |
| y10 = MINOR | 0, 1 | - | - | |
| η3 = Genetic (GEN) | y11 − 23 = significant SNP from GWAS | |||
| ξ1 = Time (TIME) | x1 = barn age [in years] | 1, 2, 3 | - | - |
| x2 = average first calving age [in days] | 750–760, 760.1–775, 775.1–780, 780.1–800, >800 | - | - | |
| x3 = average calving interval [in days] | 382–432 | 407 | 0.17 | |
| x4 = lactation stage [in days] | 0–100, 100.1–200, 200.1–300, >300 | - | - | |
| formative model | y24 = housing system | compost, cubicle | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, P.; Brügemann, K.; Yin, T.; Engel, P.; König, S. Inferring Causalities of Environmental and Genetic Factors for Differential Somatic Cell Count and Mastitis Pathogens in Dairy Cows Using Structural Equation Modelling. Genes 2023, 14, 2102. https://doi.org/10.3390/genes14112102
Wagner P, Brügemann K, Yin T, Engel P, König S. Inferring Causalities of Environmental and Genetic Factors for Differential Somatic Cell Count and Mastitis Pathogens in Dairy Cows Using Structural Equation Modelling. Genes. 2023; 14(11):2102. https://doi.org/10.3390/genes14112102
Chicago/Turabian StyleWagner, Patricia, Kerstin Brügemann, Tong Yin, Petra Engel, and Sven König. 2023. "Inferring Causalities of Environmental and Genetic Factors for Differential Somatic Cell Count and Mastitis Pathogens in Dairy Cows Using Structural Equation Modelling" Genes 14, no. 11: 2102. https://doi.org/10.3390/genes14112102