
The Role of lncRNAs in Pig Muscle in Response to Cold Exposure

Abstract
:1. Introduction
2. Materials and Methods
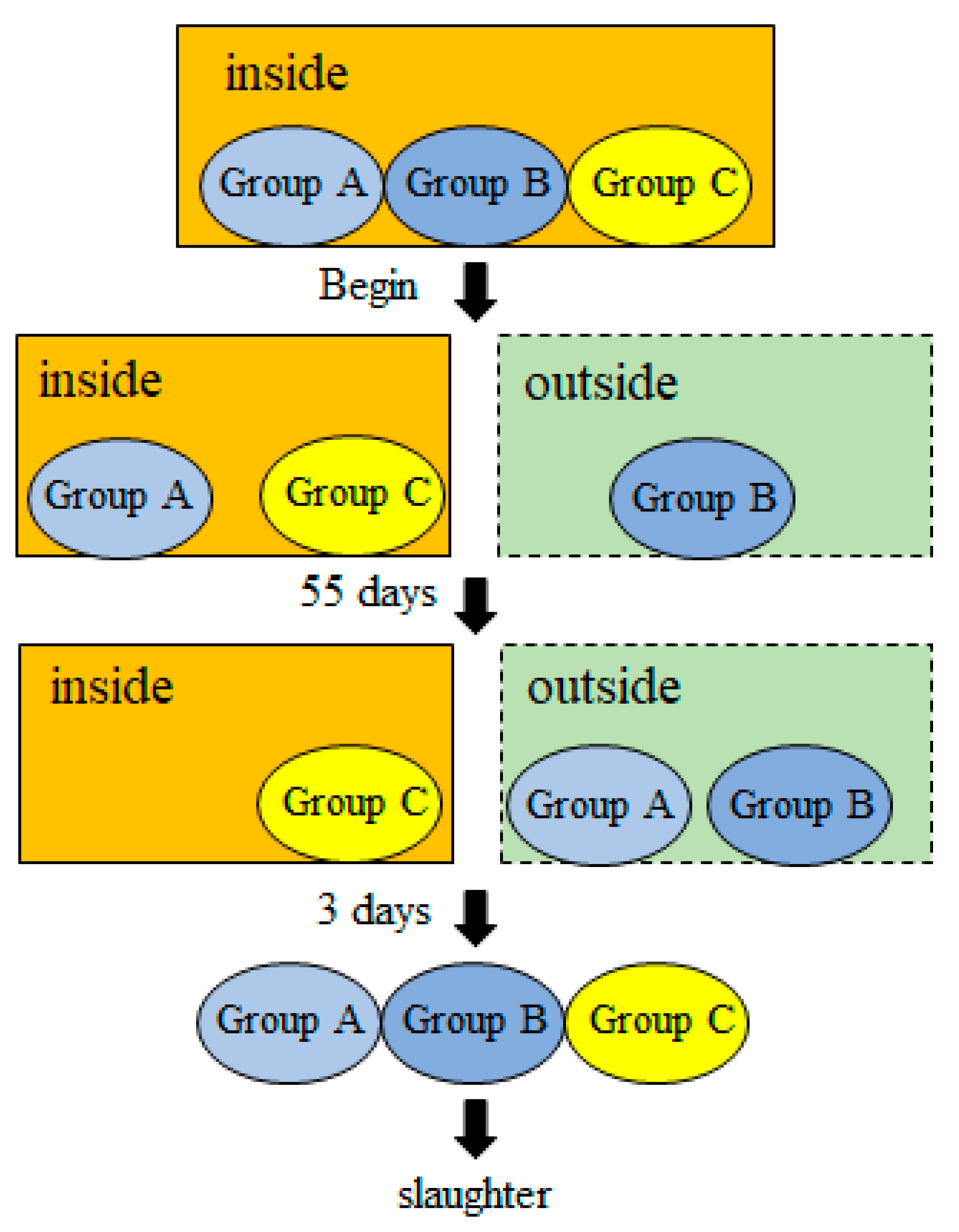
2.1. Experimental Design and Sample Collection
2.2. cDNA Library Construction and Sequencing
2.3. Identification of lncRNAs
2.4. Analysis of DElncRNAs
2.5. lncRNA-mRNA Network Construction
3. Results
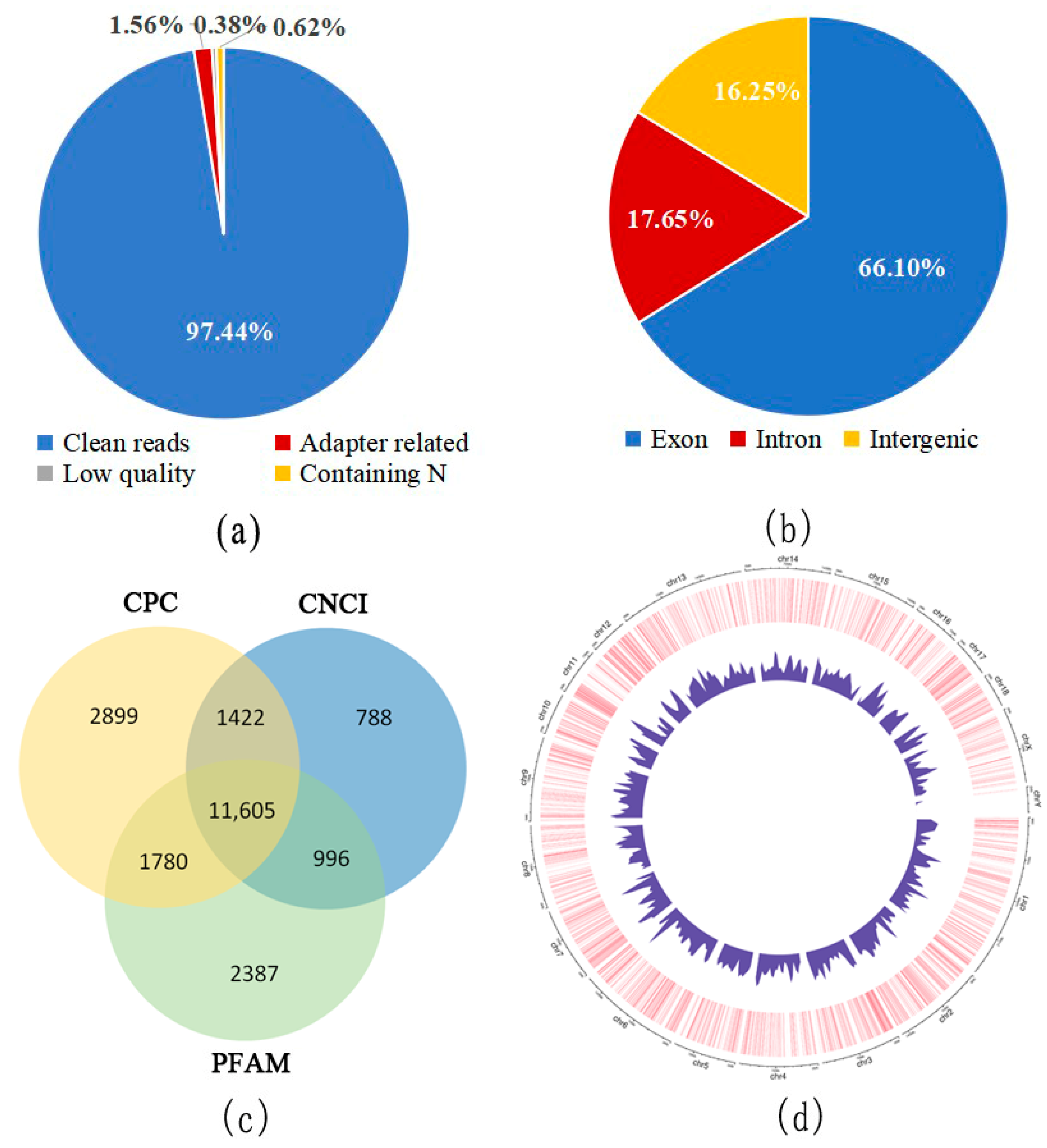
3.1. Identification of lncRNAs in the Dorsal Muscle of Pigs
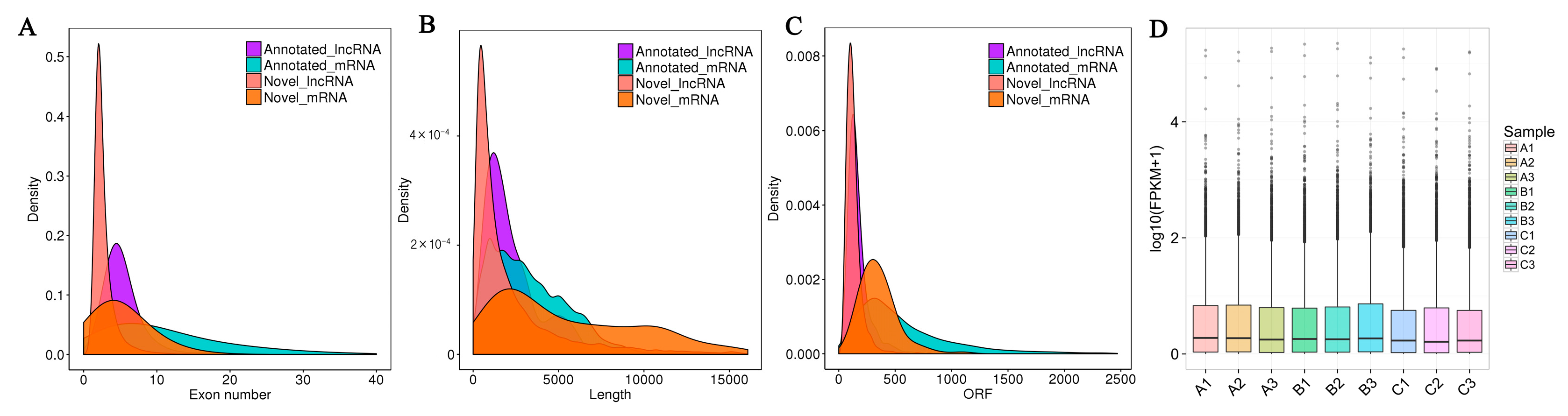
3.2. Characterization of lncRNAs in the Dorsal Muscle
3.3. Differential Expression (DE) of Cold Stress-Related lncRNAs
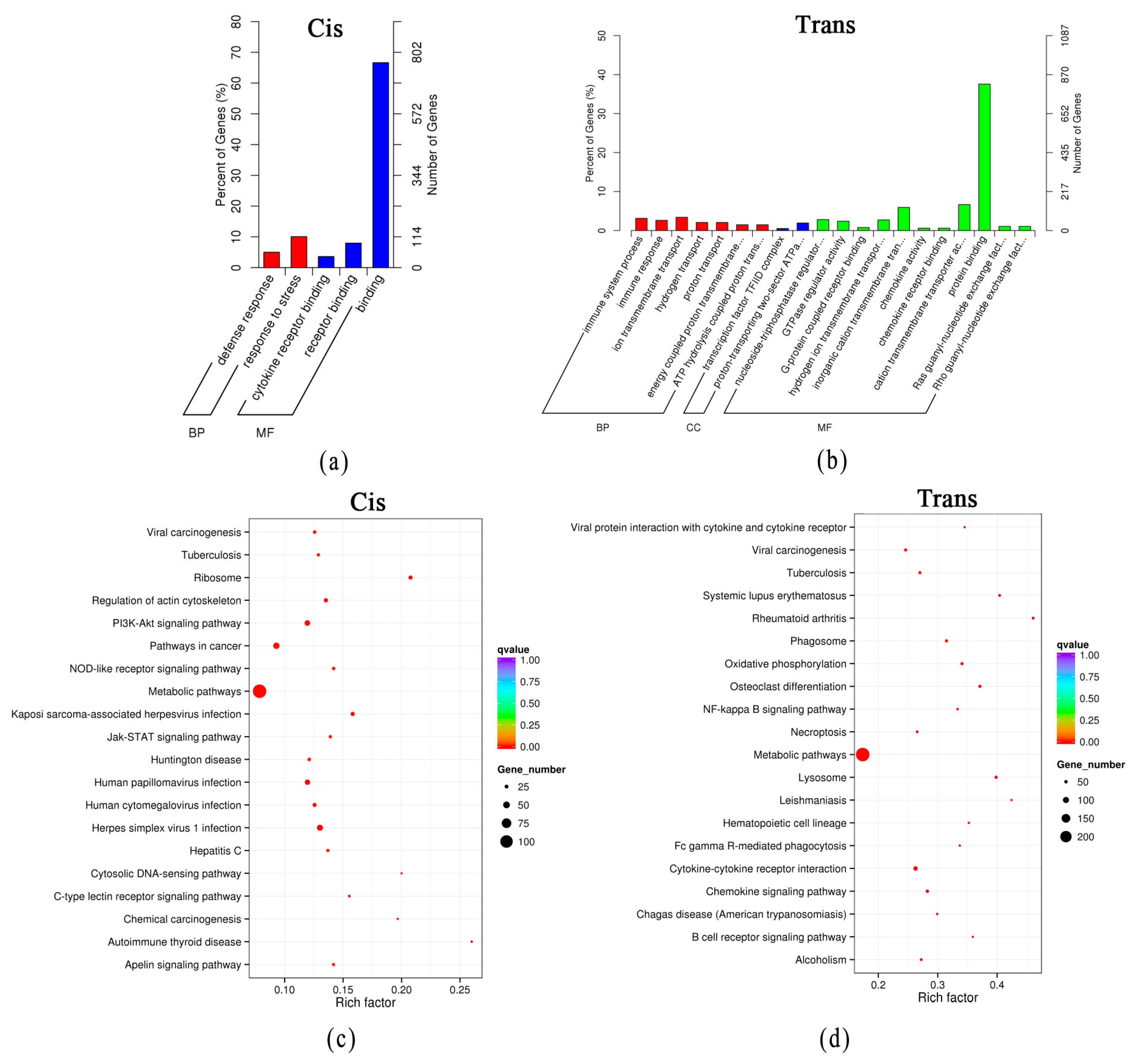
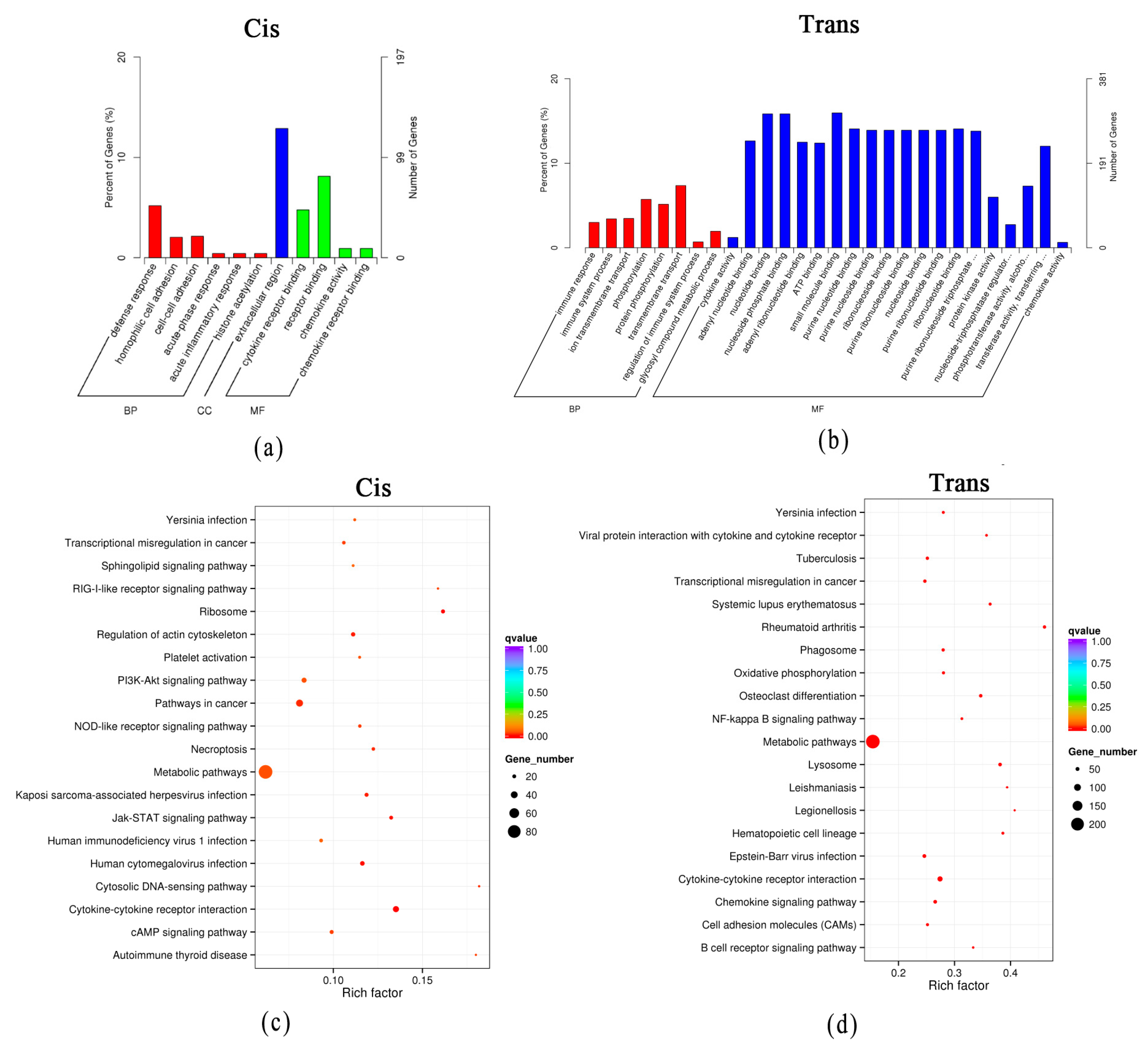
3.4. Functional Prediction of DElncRNAs through Their Target Genes
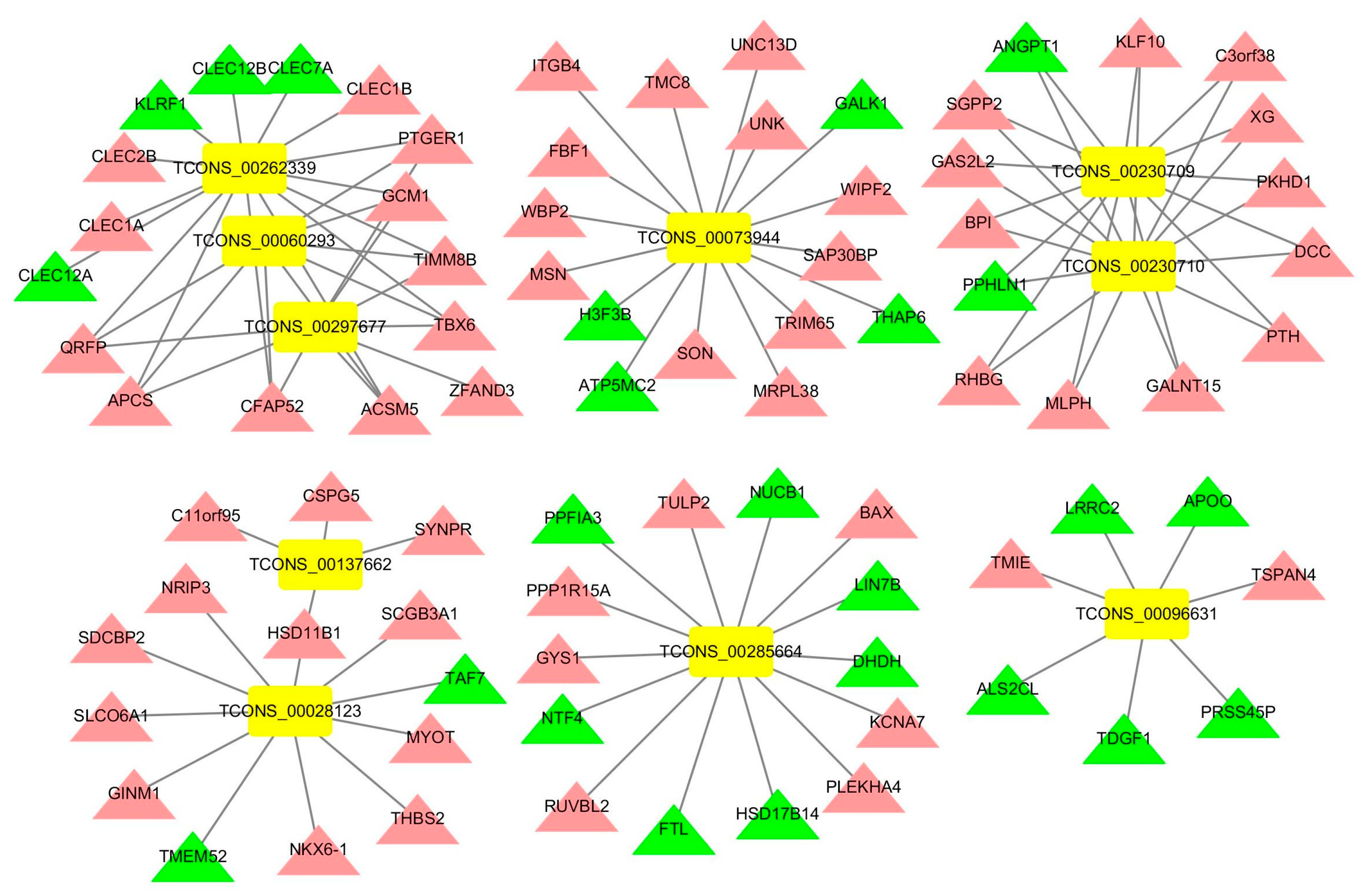
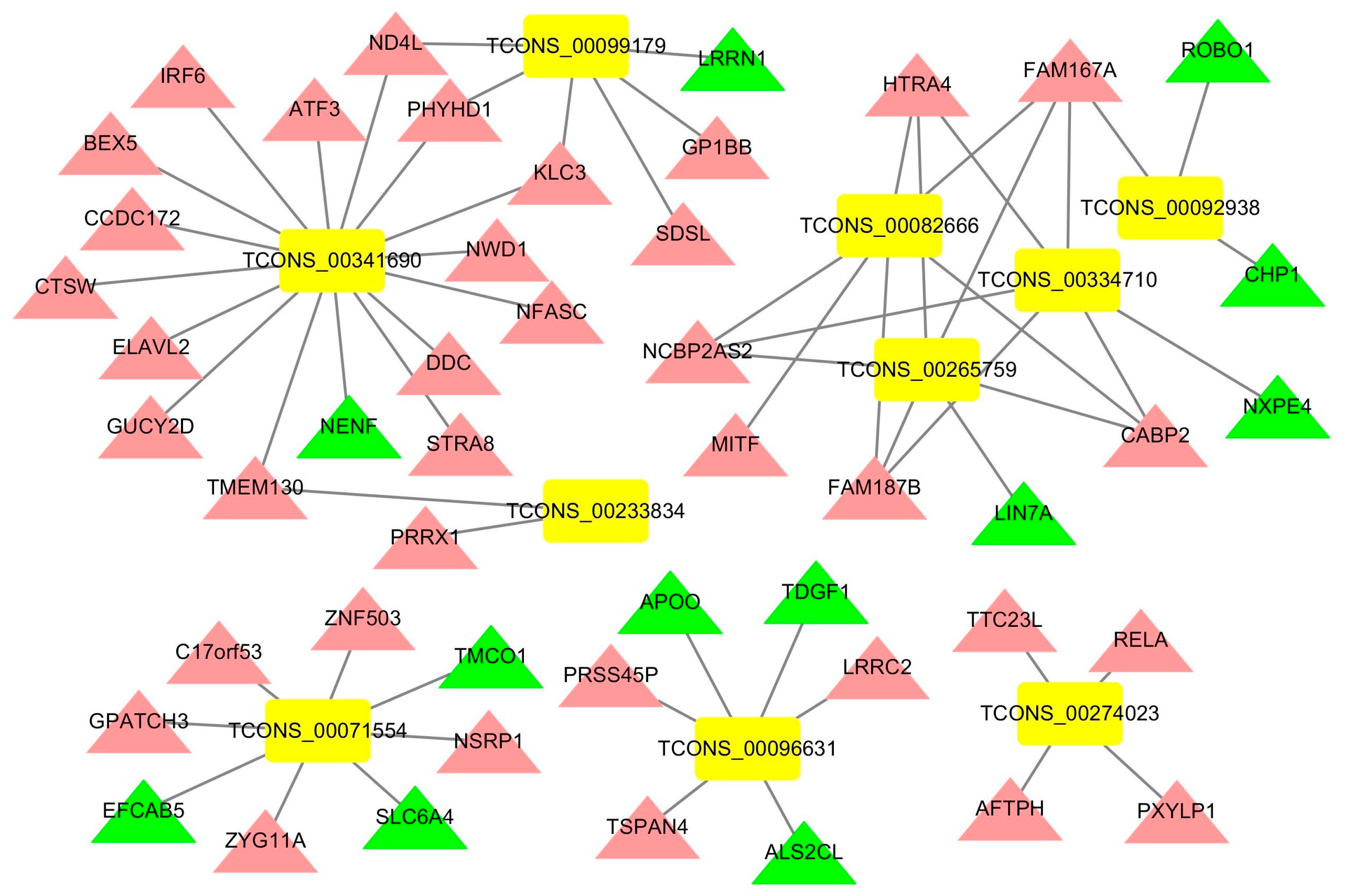
3.5. Construction of an lncRNA-mRNA Network for the Top 10 Upregulated DElncRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramirez, B.C.; Hayes, M.D.; Condotta, I.C.; Leonard, S.M. Impact of Housing Environment and Management on Pre-/Post-Weaning Piglet Productivity. J. Anim. Sci. 2022, 100, skac142. [Google Scholar] [CrossRef]
- Li, M.; Tian, S.; Jin, L.; Zhou, G.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.; et al. Genomic Analyses Identify Distinct Patterns of Selection in Domesticated Pigs and Tibetan Wild Boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef]
- Ratky, J.; Egerszegi, I.; Toth, P.; Keonuchan, S.; Nagai, T.; Kikuchi, K.; Manabe, N.; Brussow, K.P. Saving Genetic Resources of Native Pigs in Occidental and Oriental Countries-Practical Examples of the Characterization and Utilization of Native Pigs in Hungary and Laos. J. Reprod. Dev. 2013, 59, 437–441. [Google Scholar] [CrossRef]
- Thutwa, K.; Chabo, R.; Nsoso, S.J.; Mareko, M.; Kgwatalala, P.M.; Owusu-Sekyere, E. Indigenous Tswana Pig Production Characteristics and Management Practices in Southern Districts of Botswana. Trop. Anim. Health. Prod. 2020, 52, 517–524. [Google Scholar] [CrossRef]
- Frigault, J.J.; Morin, M.D.; Morin, P.J. Differential Expression and Emerging Functions of Non-Coding RNAs in Cold Adaptation. J. Comp. Physiol. B 2017, 187, 19–28. [Google Scholar] [CrossRef]
- Chen, W.; Xu, Z.; You, W.; Zhou, Y.; Wang, L.; Huang, Y.; Shan, T. Cold Exposure Alters Lipid Metabolism of Skeletal Muscle through Hif-1alpha-Induced Mitophagy. BMC Biol. 2023, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Sarver, D.C.; Xu, C.; Rodriguez, S.; Aja, S.; Jaffe, A.E.; Gao, F.J.; Delannoy, M.; Periasamy, M.; Kazuki, Y.; Oshimura, M.; et al. Hypermetabolism in Mice Carrying a near-Complete Human Chromosome 21. eLife 2023, 12, e86023. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, Y.; Zha, X.; Cao, H.; Huang, S.; Cao, D.; Zhang, K.; Xie, T.; Xu, X.; Liang, Z.; et al. Cold-Sensitive Ventromedial Hypothalamic Neurons Control Homeostatic Thermogenesis and Social Interaction-Associated Hyperthermia. Cell Metab. 2022, 34, 888–901.e5. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Li, L.; Li, M.; Lam, S.M.; Wang, G.; Wu, Y.; Zhang, H.; Niu, C.; Zhang, X.; Liu, X.; et al. Microbiota Depletion Impairs Thermogenesis of Brown Adipose Tissue and Browning of White Adipose Tissue. Cell Rep. 2019, 26, 2720–2737.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, S.; Cao, C.; Chen, C.; Liu, J.; Wang, Y.; Liu, J.; Zhao, J.; Tao, C.; Wang, Y. Functional and Genetic Characterization of Porcine Beige Adipocytes. Cells 2022, 11, 751. [Google Scholar] [CrossRef] [PubMed]
- Grigg, G.; Nowack, J.; Bicudo, J.; Bal, N.C.; Woodward, H.N.; Seymour, R.S. Whole-Body Endothermy: Ancient, Homologous and Widespread among the Ancestors of Mammals, Birds and Crocodylians. Biol. Rev. Camb. Philos. Soc. 2022, 97, 766–801. [Google Scholar] [CrossRef] [PubMed]
- Nowack, J.; Giroud, S.; Arnold, W.; Ruf, T. Muscle Non-Shivering Thermogenesis and Its Role in the Evolution of Endothermy. Front. Physiol. 2017, 8, 889. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, S.; Cheviron, Z.A.; Storz, J.F.; McClelland, G.B.; Scott, G.R. Chronic Cold Exposure Induces Mitochondrial Plasticity in Deer Mice Native to High Altitudes. J. Physiol. 2020, 598, 5411–5426. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, X.Y.; Wang, C.Z.; Wang, D.H. Recruitment of Muscle Genes as an Effect of Brown Adipose Tissue Ablation in Cold-Acclimated Brandt’s Voles (Lasiopodomys Brandtii). Int. J. Mol. Sci. 2022, 24, 342. [Google Scholar] [CrossRef]
- Ezzatpanah, S.; Eriksen, M.B.; Moe, A.G.; Haugen, F. Diminished Cold Avoidance Behaviours after Chronic Cold Exposure-Potential Involvement of Trpm8. Neuroscience 2021, 469, 17–30. [Google Scholar] [CrossRef]
- Zhan, H.; Xiong, Y.; Wang, Z.; Dong, W.; Zhou, Q.; Xie, S.; Li, X.; Zhao, S.; Ma, Y. Integrative Analysis of Transcriptomic and Metabolomic Profiles Reveal the Complex Molecular Regulatory Network of Meat Quality in Enshi Black Pigs. Meat Sci. 2022, 183, 108642. [Google Scholar] [CrossRef]
- Zhang, D.; Ma, S.Z.; Wang, L.; Ma, H.; Wang, W.; Xia, J.Q.; Liu, D. Min Pig Skeletal Muscle Response to Cold Stress. PLoS ONE 2022, 17, e0274184. [Google Scholar] [CrossRef]
- Power, C.; Lamarre, S.G.; Dion-Cote, A.M. Transcriptional and Metabolomic Investigation of the Stress Response in Snow Crab During Simulated Transport Condition (Chionoecetes Opilio). Comp. Biochem. Physiol. Part D Genom. Proteom. 2023, 46, 101079. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Singh, S.; Ganguly, I.; Bhatia, A.K.; Dixit, S.P. Deciphering Local Adaptation of Native Indian Cattle (Bos Indicus) Breeds Using Landscape Genomics and in-Silico Prediction of Deleterious Snp Effects on Protein Structure and Function. 3 Biotech 2023, 13, 86. [Google Scholar] [CrossRef]
- Ji, K.; Jiao, D.; Yang, G.; Degen, A.A.; Zhou, J.; Liu, H.; Wang, W.; Cong, H. Transcriptome Analysis Revealed Potential Genes Involved in Thermogenesis in Muscle Tissue in Cold-Exposed Lambs. Front. Genet. 2022, 13, 1017458. [Google Scholar] [CrossRef]
- Zhu, M.; Dong, Q.S.; Bing, J.; Songbuerbatu; Zheng, L.; Dorjee, T.; Liu, Q.; Zhou, Y.J.; Gao, F. Combined LncRNA and mRNA Expression Profiles Identified the LncRNA-MiRNA-mRNA Modules Regulating the Cold Stress Response in Ammopiptanthus Nanus. Int. J. Mol. Sci. 2023, 24, 6502. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Niu, C.; Zhan, X.; Xu, J.; Lian, S.; Xu, B.; Guo, J.; Zhen, L.; Yang, H.; Li, S.; et al. Identification, Functional Prediction, and Key LncRNA Verification of Cold Stress-Related LncRNAs in Rats Liver. Sci. Rep. 2020, 10, 521. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Lin, Y.; Li, H.; Chen, Y.; Wei, W.; Li, P.; Chen, L. Transcriptome Sequencing Reveals the Differentially Expressed LncRNAs and mRNAs in Response to Cold Acclimation and Cold Stress in Pomacea Canaliculata. BMC Genom. 2022, 23, 382. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, J.; Liang, Q.; Zhang, Y.; Kang, K.; Wang, W.; Feng, Y.; Wu, S.; Yang, C.; Li, Y. Genome-Wide Analysis of Long Noncoding RNAs Affecting Floral Bud Dormancy in Pears in Response to Cold Stress. Tree Physiol. 2021, 41, 771–790. [Google Scholar] [CrossRef]
- Liu, Y.; Cai, Y.; Li, Y.; Zhang, X.; Shi, N.; Zhao, J.; Yang, H. Dynamic Changes in the Transcriptome Landscape of Arabidopsis Thaliana in Response to Cold Stress. Front. Plant Sci. 2022, 13, 983460. [Google Scholar] [CrossRef]
- Liu, X.; Huang, C.; Jiang, T.; Sun, X.; Zhan, S.; Zhong, T.; Guo, J.; Dai, D.; Wang, Y.; Li, L.; et al. Lncdgat2 Is a Novel Positive Regulator of the Goat Adipocyte Thermogenic Gene Program. Int. J. Biol. Macromol. 2023, 245, 125465. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, R.; Liang, J.; Izaz, A.; Shu, Y.; Pan, T.; Wu, X. Molecular Mechanism of Chinese Alligator (Alligator Sinensis) Adapting to Hibernation. J. Exp. Zool. B Mol. Dev. Evol. 2021, 336, 32–49. [Google Scholar] [CrossRef]
- Liu, Y.; Shi, H.; Hu, Y.; Yao, R.; Liu, P.; Yang, Y.; Li, S. RNA Binding Motif Protein 3 (Rbm3) Promotes Protein Kinase B (Akt) Activation to Enhance Glucose Metabolism and Reduce Apoptosis in Skeletal Muscle of Mice under Acute Cold Exposure. Cell Stress Chaperones 2022, 27, 603–618. [Google Scholar] [CrossRef]
- Yang, C.; Cao, C.; Liu, J.; Zhao, Y.; Pan, J.; Tao, C.; Wang, Y. Distinct Transcriptional Responses of Skeletal Muscle to Short-Term Cold Exposure in Tibetan Pigs and Bama Pigs. Int. J. Mol. Sci. 2023, 24, 7431. [Google Scholar] [CrossRef]
- Zhang, X.; Niu, Y.; Zhang, H.; Xu, T.; Zeng, Q.; Storey, K.B.; Chen, Q. The Effect of Long-Term Cold Acclimation on Redox State and Antioxidant Defense in the High-Altitude Frog, Nanorana Pleskei. J. Therm. Biol. 2021, 99, 103008. [Google Scholar] [CrossRef]
- Gou, N.; Ji, H.; Wu, W.; Zhong, M.; Zhang, B. Transcriptional Response to Cold and Fasting Acclimation in Onychostoma Macrolepis During the Overwintering Stage. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100901. [Google Scholar] [CrossRef]
- Garcia, C.F.; Oviedo-Diego, M.; Laino, A.; Peterson, G.; Mattoni, C.I.; Peretti, A.V.; Ojanguren-Affilastro, A.A. Low Temperatures Induce Physiological Changes in Lipids, Fatty Acids and Hydrocarbons, in Two Rare Winter Scorpions of Genus Urophonius (Scorpiones, Bothriuridae). J. Therm. Biol. 2021, 96, 102841. [Google Scholar] [CrossRef] [PubMed]
- Blondin, D.P.; Daoud, A.; Taylor, T.; Tingelstad, H.C.; Bezaire, V.; Richard, D.; Carpentier, A.C.; Taylor, A.W.; Harper, M.E.; Aguer, C.; et al. Four-Week Cold Acclimation in Adult Humans Shifts Uncoupling Thermogenesis from Skeletal Muscles to Brown Adipose Tissue. J. Physiol. 2017, 595, 2099–2113. [Google Scholar] [CrossRef] [PubMed]
- Shaftoe, J.B.; Manchester, E.A.; Gillis, T.E. Cardiac Remodeling Caused by Cold Acclimation Is Reversible with Rewarming in Zebrafish (Danio Rerio). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2023, 283, 111466. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.; Jara-Oseguera, A.; Aldrich, R.W. Implications of a Temperature-Dependent Heat Capacity for Temperature-Gated Ion Channels. Proc. Natl. Acad. Sci. USA 2023, 120, e2301528120. [Google Scholar] [CrossRef] [PubMed]
- Samarina, L.; Wang, S.; Malyukova, L.; Bobrovskikh, A.; Doroshkov, A.; Koninskaya, N.; Shkhalakhova, R.; Matskiv, A.; Fedorina, J.; Fizikova, A.; et al. Long-Term Cold, Freezing and Drought: Overlapping and Specific Regulatory Mechanisms and Signal Transduction in Tea Plant (Camellia Sinensis (L.) Kuntze). Front. Plant Sci. 2023, 14, 1145793. [Google Scholar] [CrossRef] [PubMed]
- Gusain, S.; Joshi, S.; Joshi, R. Sensing, Signalling, and Regulatory Mechanism of Cold-Stress Tolerance in Plants. Plant Physiol. Biochem. 2023, 197, 107646. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, H.; Wu, S.; Fu, D.; Li, M.; Gong, Z.; Yang, S. Cpk28-Nlp7 Module Integrates Cold-Induced Ca(2+) Signal and Transcriptional Reprogramming in Arabidopsis. Sci. Adv. 2022, 8, eabn7901. [Google Scholar] [CrossRef]
- He, M.; Yang, X.; Liu, T.; Zhang, X.; Ji, X.; Wei, Y.; Zhang, Q. Hybrid Histidine Kinase Hisk2301 Modulates Carotenoid Production to Counteract Cold-Induced Oxidative Stress in Rhodosporidium Kratochvilovae Ym25235 under Low Temperature. Antonie Van Leeuwenhoek 2022, 115, 1393–1404. [Google Scholar] [CrossRef]
- Xu, Z.; Regenstein, J.M.; Xie, D.; Lu, W.; Ren, X.; Yuan, J.; Mao, L. The Oxidative Stress and Antioxidant Responses of Litopenaeus Vannamei to Low Temperature and Air Exposure. Fish Shellfish Immunol. 2018, 72, 564–571. [Google Scholar] [CrossRef]
- Lu, D.L.; Ma, Q.; Sun, S.X.; Zhang, H.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. Reduced Oxidative Stress Increases Acute Cold Stress Tolerance in Zebrafish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 235, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.M.; Li, Z.; Storey, K.B. Hypoxia Inducible Factor-1alpha Responds to Freezing, Anoxia and Dehydration Stresses in a Freeze-Tolerant Frog. Cryobiology 2023, 110, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, E.; Tranberg, H.K.; Sollid, J.; Nilsson, G.E.; Nikinmaa, M. Temperature Regulates Hypoxia-Inducible Factor-1 (Hif-1) in a Poikilothermic Vertebrate, Crucian Carp (Carassius Carassius). J. Exp. Biol. 2006, 209, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Morin, P., Jr.; McMullen, D.C.; Storey, K.B. Hif-1alpha Involvement in Low Temperature and Anoxia Survival by a Freeze Tolerant Insect. Mol. Cell Biochem. 2005, 280, 99–106. [Google Scholar] [CrossRef]
- Jankovic, A.; Korac, A.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Vucetic, M.; Markelic, M.; Velickovic, K.; Golic, I.; Korac, B. Endocrine and Metabolic Signaling in Retroperitoneal White Adipose Tissue Remodeling During Cold Acclimation. J. Obes. 2013, 2013, 937572. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, W.; Romanelli, L.; Atkins, M.; Hillen, H.; Bravo Gonzalez-Blas, C.; Jacobs, J.; Xie, J.; Soheily, S.; Verboven, E.; Moya, I.M.; et al. Hippo Signaling Instructs Ectopic but Not Normal Organ Growth. Science 2022, 378, eabg3679. [Google Scholar] [CrossRef]
- Zeybek, N.D.; Baysal, E.; Bozdemir, O.; Buber, E. Hippo Signaling: A Stress Response Pathway in Stem Cells. Curr. Stem Cell Res. Ther. 2021, 16, 824–839. [Google Scholar] [CrossRef]
- Tirpe, A.A.; Gulei, D.; Ciortea, S.M.; Crivii, C.; Berindan-Neagoe, I. Hypoxia: Overview on Hypoxia-Mediated Mechanisms with a Focus on the Role of Hif Genes. Int. J. Mol. Sci. 2019, 20, 6140. [Google Scholar] [CrossRef]
- Song, Y.; Bi, Z.; Liu, Y.; Qin, F.; Wei, Y.; Wei, X. Targeting Ras-Raf-Mek-Erk Signaling Pathway in Human Cancer: Current Status in Clinical Trials. Genes Dis. 2023, 10, 76–88. [Google Scholar] [CrossRef]
- Lopez-Soriano, F.J.; Alemany, M. Effect of Cold-Temperature Exposure and Acclimation on Amino Acid Pool Changes and Enzyme Activities of Rat Brown Adipose Tissue. Biochim. Biophys. Acta 1987, 925, 265–271. [Google Scholar] [CrossRef]
- Dassarma, S.; Capes, M.D.; Karan, R.; Dassarma, P. Amino Acid Substitutions in Cold-Adapted Proteins from Halorubrum Lacusprofundi, an Extremely Halophilic Microbe from Antarctica. PLoS ONE 2013, 8, e58587. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, Y.; Zheng, Z.; Deng, Y.; Jiao, Y.; Du, X. Adaptive Response of Pearl Oyster Pinctada Fucata Martensii to Low Water Temperature Stress. Fish Shellfish Immunol. 2018, 78, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Orzechowski, S.; Sitnicka, D.; Grabowska, A.; Compart, J.; Fettke, J.; Zdunek-Zastocka, E. Effect of Short-Term Cold Treatment on Carbohydrate Metabolism in Potato Leaves. Int. J. Mol. Sci. 2021, 22, 7203. [Google Scholar] [CrossRef]
- Ouyang, L.; Leus, L.; De Keyser, E.; Van Labeke, M.C. Seasonal Changes in Cold Hardiness and Carbohydrate Metabolism in Four Garden Rose Cultivars. J. Plant Physiol. 2019, 232, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Furtauer, L.; Weiszmann, J.; Weckwerth, W.; Nagele, T. Dynamics of Plant Metabolism During Cold Acclimation. Int. J. Mol. Sci. 2019, 20, 5411. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A vs. C Group | B vs. C Group | ||||||
|---|---|---|---|---|---|---|---|
| lncRNA | FDR | |log2FC| | Regulated | lncRNA | FDR | |log2FC| | Regulated |
| Linc1620 | 0.0006 | 14.09 | up | Linc862 | 2.83 × 10−6 | 13.11 | down |
| Linc361 | 0.0118 | 11.96 | up | TCONS_00340242 | 0.0065 | 12.55 | down |
| TCONS_00349979 | 0.0127 | 11.87 | down | TCONS_00344988 | 0.0001 | 11.86 | down |
| TCONS_00000583 | 0.0014 | 10.80 | down | TCONS_00233834 | 0.0015 | 11.12 | up |
| TCONS_00073944 | 0.0021 | 10.38 | up | TCONS_00000583 | 0.0031 | 10.78 | down |
| Linc1918 | 0.0001 | 10.01 | down | Linc1207 | 0.0037 | 10.71 | up |
| Linc3782 | 0.0083 | 9.98 | down | Linc938 | 0.0042 | 10.53 | down |
| TCONS_00284293 | 0.0098 | 9.62 | up | Linc2552 | 0.0053 | 10.41 | down |
| TCONS_00244588 | 0.0102 | 9.49 | up | TCONS_00275432 | 0.0080 | 10.07 | down |
| TCONS_00050025 | 0.0194 | 9.14 | up | Linc1680 | 0.0123 | 9.96 | up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, D.; Wang, L.; Wang, W.; Liu, D. The Role of lncRNAs in Pig Muscle in Response to Cold Exposure. Genes 2023, 14, 1901. https://doi.org/10.3390/genes14101901
Zhang D, Wang L, Wang W, Liu D. The Role of lncRNAs in Pig Muscle in Response to Cold Exposure. Genes. 2023; 14(10):1901. https://doi.org/10.3390/genes14101901
Chicago/Turabian StyleZhang, Dongjie, Liang Wang, Wentao Wang, and Di Liu. 2023. "The Role of lncRNAs in Pig Muscle in Response to Cold Exposure" Genes 14, no. 10: 1901. https://doi.org/10.3390/genes14101901