
A Redesigned Method for CNP-Synchronized In Vitro Maturation Inhibits Oxidative Stress and Apoptosis in Cumulus-Oocyte Complexes and Improves the Developmental Potential of Porcine Oocytes
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Collection of Cumulus-Oocyte-Complexes (COCs), Denuded Oocytes (DOs), Cumulus Cells (CCs) and Mural Granulosa Cells (MGCs)
2.2. In Vitro Maturation of Porcine COCs
2.3. Assessment of Nuclear Maturation
2.4. Parthenogenetic Activation (PA)
2.5. Immunofluorescence Staining (IF)
2.6. Detection of Intracellular Reactive Oxygen Species (ROS) in the Oocytes
2.7. Detection of Early Apoptosis by Annexin-V Staining in the Oocytes
2.8. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick-End Labeling (TUNEL) Assay
2.9. RNA Extraction and Real-Time Reverse Transcription Quantitative PCR (RT-qPCR) Analysis
2.10. Western Blot Analysis
2.11. Statistical Analysis
3. Results
3.1. The Expression Pattern of CNP and NPR2 in Porcine Ovarian Follicles
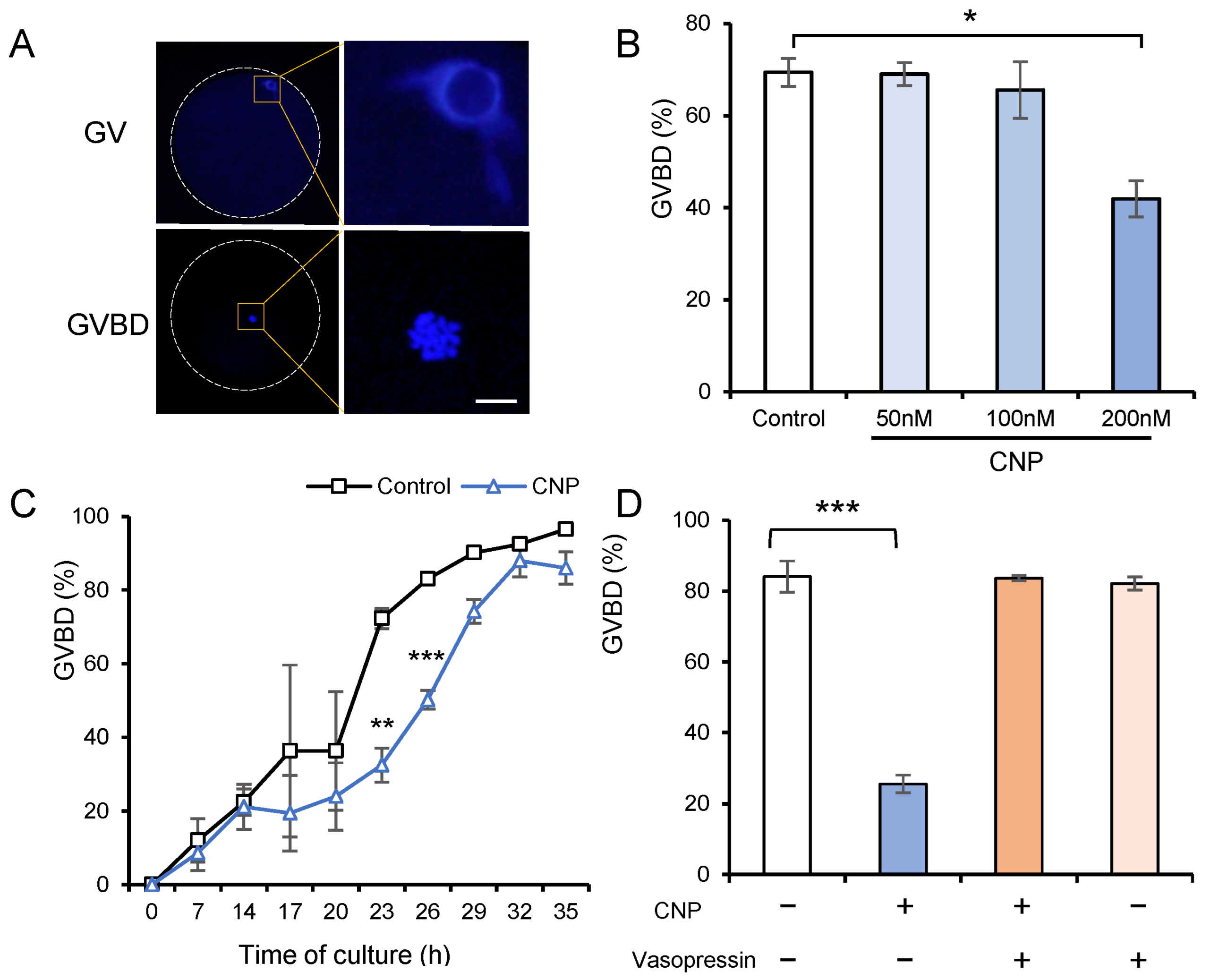
3.2. CNP Exposure of In Vitro Cultured COCs Maintains the Meiotic Arrest of Porcine Oocytes
3.3. CNP-Based Biphasic IVM System Improves the Developmental Potential of In Vitro Matured Porcine Oocytes
3.4. CNP Alleviates Oxidative Stress and Early Apoptosis in Porcine Oocytes
3.5. CNP Reduces DNA Damage and Apoptosis in Cumulus Cells
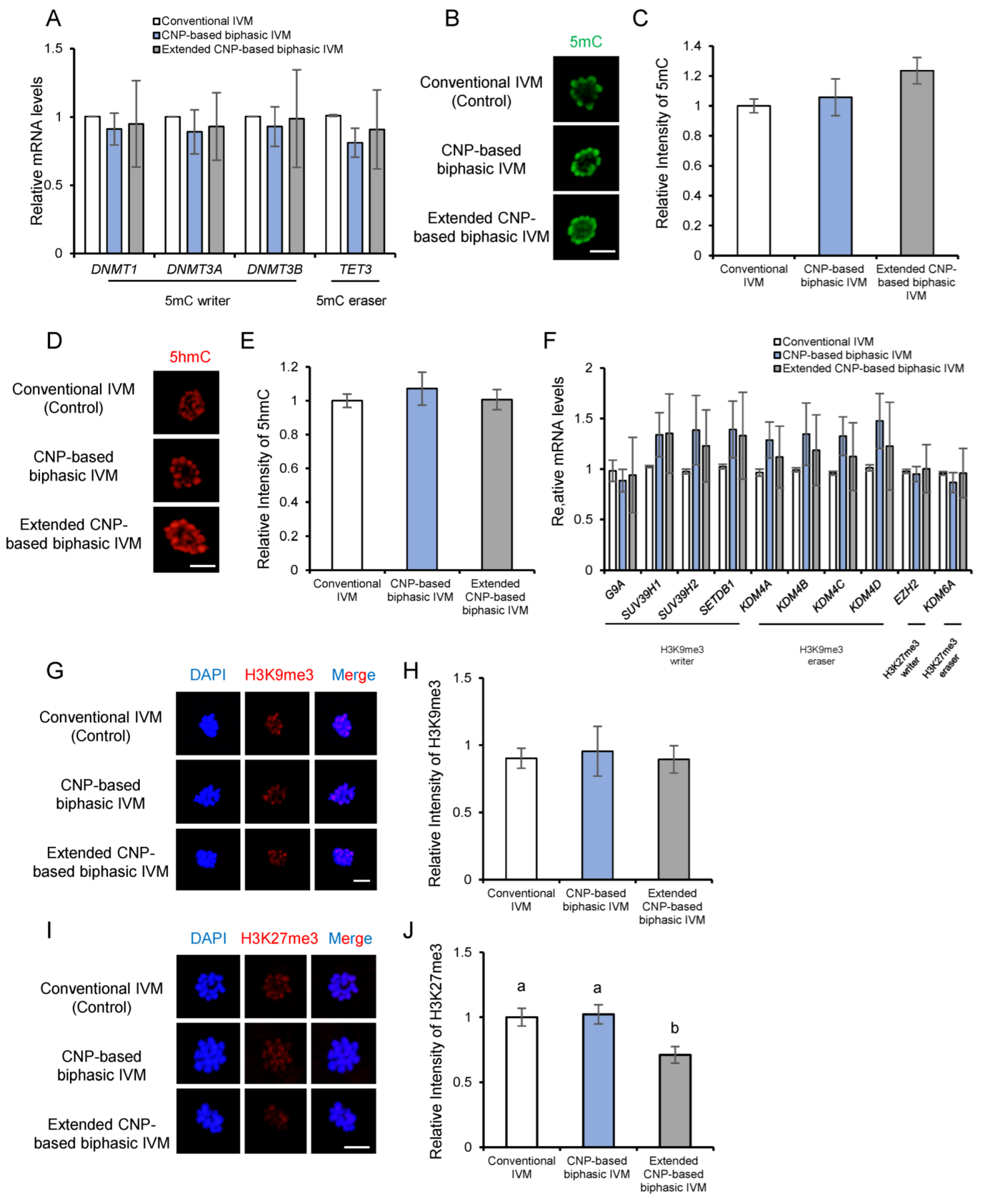
3.6. CNP Does Not Affect Epigenetic Modifications of In Vitro Matured Porcine Oocytes
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eppig, J.J. Coordination of nuclear and cytoplasmic oocyte maturation in eutherian mammals. Reprod. Fertil. Dev. 1996, 8, 485–489. [Google Scholar] [CrossRef] [PubMed]
- Josefsberg, L.B.; Galiani, D.; Lazar, S.; Kaufman, O.; Seger, R.; Dekel, N. MPF governs MAPK activation and interphase suppression during meiosis of rat oocytes. Biol. Reprod. 2003, 68, 1282–1290. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.P.; Ratzan, W.J.; Freudzon, M.; Mehlmann, L.M.; Krall, J.; Movsesian, M.A.; Wang, H.; Ke, H.; Nikolaev, V.O.; Jaffe, L.A. Cyclic GMP from the surrounding somatic cells regulates cyclic AMP and meiosis in the mouse oocyte. Development 2009, 136, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.G. Maturation in vitro of mouse, sheep, cow, pig, rhesus monkey and human ovarian oocytes. Nature 1965, 208, 349–351. [Google Scholar] [CrossRef]
- Schultz, R.M.; Montgomery, R.R.; Belanoff, J.R. Regulation of mouse oocyte meiotic maturation: Implication of a decrease in oocyte cAMP and protein dephosphorylation in commitment to resume meiosis. Dev. Biol. 1983, 97, 264–273. [Google Scholar] [CrossRef]
- Gilchrist, R.B.; Luciano, A.M.; Richani, D.; Zeng, H.T.; Wang, X.; De Vos, M.; Sugimura, S.; Smitz, J.; Richard, F.J.; Thompson, J.G. Oocyte maturation and quality: Role of cyclic nucleotides. Reproduction 2016, 152, R143–R157. [Google Scholar] [CrossRef]
- Thomas, R.E.; Thompson, J.G.; Armstrong, D.T.; Gilchrist, R.B. Effect of specific phosphodiesterase isoenzyme inhibitors during in vitro maturation of bovine oocytes on meiotic and developmental capacity. Biol. Reprod. 2004, 71, 1142–1149. [Google Scholar] [CrossRef]
- Shu, Y.; Zeng, H.; Ren, Z.; Zhuang, G.; Liang, X.; Shen, H.; Yao, S.; Ke, P.; Wang, N. Effects of cilostamide and forskolin on the meiotic resumption and embryonic development of immature human oocytes. Hum. Reprod. 2008, 23, 504–513. [Google Scholar] [CrossRef]
- Albuz, F.K.; Sasseville, M.; Lane, M.; Armstrong, D.T.; Thompson, J.G.; Gilchrist, R.B. Simulated physiological oocyte maturation (SPOM): A novel in vitro maturation system that substantially improves embryo yield and pregnancy outcomes. Hum. Reprod. 2010, 25, 2999–3011. [Google Scholar] [CrossRef]
- Leal, G.R.; Monteiro, C.A.S.; Souza-Fabjan, J.M.G.; de Paula Vasconcelos, C.O.; Nogueira, L.A.G.; Ferreira, A.M.R.; Serapião, R.V. Role of cAMP modulator supplementations during oocyte in vitro maturation in domestic animals. Anim. Reprod. Sci. 2018, 199, 1–14. [Google Scholar] [CrossRef]
- Xi, G.; An, L.; Jia, Z.; Tan, K.; Zhang, J.; Wang, Z.; Zhang, C.; Miao, K.; Wu, Z.; Tian, J. Natriuretic peptide receptor 2 (NPR2) localized in bovine oocyte underlies a unique mechanism for C-type natriuretic peptide (CNP)-induced meiotic arrest. Theriogenology 2018, 106, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Su, Y.; Sugiura, K.; Xia, G.; Eppig, J.J. Granulosa cell ligand NPPC and its receptor NPR2 maintain meiotic arrest in mouse oocytes. Science 2010, 330, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Hiradate, Y.; Hoshino, Y.; Tanemura, K.; Sato, E. C-type natriuretic peptide inhibits porcine oocyte meiotic resumption. Zygote 2014, 22, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, F.; Coticchio, G.; Lodde, V.; Tessaro, I.; Modina, S.C.; Fadini, R.; Dal Canto, M.; Renzini, M.M.; Albertini, D.F.; Luciano, A.M. Natriuretic peptide precursor C delays meiotic resumption and sustains gap junction-mediated communication in bovine cumulus-enclosed oocytes. Biol. Reprod. 2014, 91, 61. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, Q.; Cai, J.; Zhao, X.; Ma, B. Effect of C-type natriuretic peptide on maturation and developmental competence of goat oocytes matured in vitro. PLoS ONE 2015, 10, e132318. [Google Scholar] [CrossRef]
- Robinson, J.W.; Zhang, M.; Shuhaibar, L.C.; Norris, R.P.; Geerts, A.; Wunder, F.; Eppig, J.J.; Potter, L.R.; Jaffe, L.A. Luteinizing hormone reduces the activity of the NPR2 guanylyl cyclase in mouse ovarian follicles, contributing to the cyclic GMP decrease that promotes resumption of meiosis in oocytes. Dev. Biol. 2012, 366, 308–316. [Google Scholar] [CrossRef]
- Wei, Q.; Zhou, C.; Yuan, M.; Miao, Y.; Zhao, X.; Ma, B. Effect of C-type natriuretic peptide on maturation and developmental competence of immature mouse oocytes in vitro. Reprod. Fertil. Dev. 2017, 29, 319–324. [Google Scholar] [CrossRef]
- Soto-Heras, S.; Paramio, M.; Thompson, J.G. Effect of pre-maturation with C-type natriuretic peptide and 3-isobutyl-1-methylxanthine on cumulus-oocyte communication and oocyte developmental competence in cattle. Anim. Reprod. Sci. 2019, 202, 49–57. [Google Scholar] [CrossRef]
- Zhang, T.; Fan, X.; Li, R.; Zhang, C.; Zhang, J. Effects of pre-incubation with C-type natriuretic peptide on nuclear maturation, mitochondrial behavior, and developmental competence of sheep oocytes. Biochem. Biophys. Res. Commun. 2018, 497, 200–206. [Google Scholar] [CrossRef]
- Soto-Heras, S.; Menéndez-Blanco, I.; Catalá, M.; Izquierdo, D.; Thompson, J.G.; Paramio, M. Biphasic in vitro maturation with C-type natriuretic peptide enhances the developmental competence of juvenile-goat oocytes. PLoS ONE 2019, 14, e221663. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Liu, W.; Yang, Y.; Wang, X.; Zhang, Z.; Guo, Q.; Wang, C.; Xia, G. Natriuretic peptides improve the developmental competence of in vitro cultured porcine oocytes. Reprod. Biol. Endocrinol. 2017, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Grupen, C.G. The evolution of porcine embryo in vitro production. Theriogenology 2014, 81, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Tatemoto, H.; Sakurai, N.; Muto, N. Protection of porcine oocytes against apoptotic cell death caused by oxidative stress during in vitro maturation: Role of cumulus cells. Biol. Reprod. 2000, 63, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Sutton-Mcdowall, M.L.; Wang, X.; Sugimura, S.; Thompson, J.G.; Gilchrist, R.B. Extending prematuration with cAMP modulators enhances the cumulus contribution to oocyte antioxidant defence and oocyte quality via gap junctions. Hum. Reprod. 2016, 31, 810–821. [Google Scholar] [CrossRef]
- Yuan, Y.Q.; Van Soom, A.; Leroy, J.; Dewulf, J.; Van Zeveren, A.; de Kruif, A.; Peelman, L.J. Apoptosis in cumulus cells, but not in oocytes, may influence bovine embryonic developmental competence. Theriogenology 2005, 63, 2147–2163. [Google Scholar] [CrossRef]
- Corn, C.M.; Hauser-Kronberger, C.; Moser, M.; Tews, G.; Ebner, T. Predictive value of cumulus cell apoptosis with regard to blastocyst development of corresponding gametes. Fertil. Steril. 2005, 84, 627–633. [Google Scholar] [CrossRef]
- Gu, L.; Wang, Q.; Sun, Q. Histone modifications during mammalian oocyte maturation: Dynamics, regulation and functions. Cell Cycle 2010, 9, 1942–1950. [Google Scholar] [CrossRef]
- Racedo, S.E.; Wrenzycki, C.; Lepikhov, K.; Salamone, D.; Walter, J.; Niemann, H. Epigenetic modifications and related mRNA expression during bovine oocyte in vitro maturation. Reprod. Fertil. Dev. 2009, 21, 738–748. [Google Scholar] [CrossRef]
- Andreu-Vieyra, C.V.; Chen, R.; Agno, J.E.; Glaser, S.; Anastassiadis, K.; Stewart, A.F.; Matzuk, M.M. MLL2 is required in oocytes for bulk histone 3 lysine 4 trimethylation and transcriptional silencing. PLoS Biol. 2010, 8, e1000453. [Google Scholar] [CrossRef]
- Xiong, X.; Zhang, X.; Yang, M.; Zhu, Y.; Yu, H.; Fei, X.; Mastuda, F.; Lan, D.; Xiong, Y.; Fu, W. Oocyte-Specific Knockout of Histone Lysine Demethylase KDM2a Compromises Fertility by Blocking the Development of Follicles and Oocytes. Int. J. Mol. Sci. 2022, 23, 12008. [Google Scholar] [CrossRef]
- Abbey, S.E.; Potter, L.R. Vasopressin-dependent inhibition of the C-type natriuretic peptide receptor, NPR-B/GC-B, requires elevated intracellular calcium concentrations. J. Biol. Chem. 2002, 277, 42423–42430. [Google Scholar] [CrossRef] [PubMed]
- Combelles, C.M.; Gupta, S.; Agarwal, A. Could oxidative stress influence the in-vitro maturation of oocytes? Reprod. Biomed. Online 2009, 18, 864–880. [Google Scholar] [CrossRef] [PubMed]
- Beaujean, N. Epigenetics, embryo quality and developmental potential. Reprod. Fertil. Dev. 2015, 27, 53–62. [Google Scholar] [CrossRef]
- Liu, H.; Aoki, F. Transcriptional activity associated with meiotic competence in fully grown mouse GV oocytes. Zygote 2002, 10, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Fair, T.; Hyttel, P.; Motlik, J.; Boland, M.; Lonergan, P. Maintenance of meiotic arrest in bovine oocytes in vitro using butyrolactone I: Effects on oocyte ultrastructure and nucleolus function. Mol. Reprod. Dev. Inc. Gamete Res. 2002, 62, 375–386. [Google Scholar] [CrossRef]
- Lonergan, P.; Faerge, I.; Hyttel, P.M.; Boland, M.; Fair, T. Ultrastructural modifications in bovine oocytes maintained in meiotic arrest in vitro using roscovitine or butyrolactone. Mol. Reprod. Dev. 2003, 64, 369–378. [Google Scholar] [CrossRef]
- Alexander, B.; Coppola, G.; Di Berardino, D.; Rho, G.J.; St John, E.; Betts, D.H.; King, W.A. The effect of 6-dimethylaminopurine (6-DMAP) and cycloheximide (CHX) on the development and chromosomal complement of sheep parthenogenetic and nuclear transfer embryos. Mol. Reprod. Dev. Inc. Gamete Res. 2006, 73, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Jia, Z.; Zhang, J.; An, L.; Wu, Z. Method for In Vitro Oocyte Maturation. Google Patents US10011818B2, 3 July 2018. [Google Scholar]
- Scheuerer, B. Factors and methods of pig oocyte and embryo quality improvement and their application in reproductive biotechnology. Reprod. Biol. 2009, 9, 97–112. [Google Scholar] [CrossRef]
- Kiyosu, C.; Tsuji, T.; Yamada, K.; Kajita, S.; Kunieda, T. NPPC/NPR2 signaling is essential for oocyte meiotic arrest and cumulus oophorus formation during follicular development in the mouse ovary. Reproduction 2012, 144, 187. [Google Scholar] [CrossRef]
- Mcgee, E.; Spears, N.; Minami, S.; Hsu, S.; Chun, S.; Billig, H.; Hsueh, A.J. Preantral ovarian follicles in serum-free culture: Suppression of apoptosis after activation of the cyclic guanosine 3′, 5′-monophosphate pathway and stimulation of growth and differentiation by follicle-stimulating hormone. Endocrinology 1997, 138, 2417–2424. [Google Scholar] [CrossRef]
- De La Fuente, R. Chromatin modifications in the germinal vesicle (GV) of mammalian oocytes. Dev. Biol. 2006, 292, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, S.; Liu, H.; Kaneko, N.; Ooga, M.; Nagata, M.; Aoki, F. Alterations in epigenetic modifications during oocyte growth in mice. Reproduction 2007, 133, 85–94. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primers | Reverse Primers |
|---|---|---|
| GAPDH | TCGGAGTGAACGGATTTG | CCTGGAAGATGGTGATGG |
| RPL19 | GGAAGGGTACTGCCAATGCT | GTGCTCCATGAGAATCCGCT |
| CNP | CCGAAGGTCCCTCGAACTC | GGAGTCTTGTCGCCCTTCTT |
| NPR2 | AATGGAGTCTAACGGTCAAG | GGAAGAAGTAGGGTTTATAGGA |
| DNMT1 | GCGTCTTGCAGGCTGGTCAGTA | CTTCTTATCATCGACCACGACGCT |
| DNMT3A | ATCAGTACGACGATGACGGC | CACCAAGAGATCCACGCACT |
| DNMT3B | ACCTGTCATCCGACACCTCT | CTCGGCATGAACCCACGTTA |
| TET3 | TCTTCCGTCGTTCAGCTACTACAG | GTGGAGGTCTGGCTTCTTCTCAAA |
| G9A | GGAGGAGCTGGGGTTTGAC | CAGAGGTGGCTGCTGAGTTG |
| SUV39H1 | GAATCAGCTCCAGGACCTGTGC | CAGGTGCTCTCTGAGTCTGGGTAC |
| SUV39H2 | GCAGGACGAACTCAACAGAA | CAACCAAAGGTGGCTTCATT |
| SETDB1 | CATTGGTTTGGATGCAGCAGC | GATGCATCATCAAAGAGCTGGTC |
| KDM4A | CTGAAACCTTGAACCCCAGTGC | GATATCGTCATAGGATGCCCGTG |
| KDM4B | CTGGCCAACAGCGAGAAGTACTG | GATGTTCCACTGGGCCACGTC |
| KDM4C | TGTGAAAAGCCAGGAGAAGCAAAG | CAGGTTTGGTCAGCCTCGGT |
| KDM4D | AAGGATGCAGTGTGTGTTGC | CCTGTTCGCGGATCTTTTTA |
| EZH2 | TGCAACACCCAATACTTACAAGC | ACTCTTTTGCTCCCTCCAAGT |
| KDM6A | GCAGGCTCAGTTGTGTAACC | GGTTTACATGCCTGCTGTGC |
| BAX | TGCCTCAGGATGCATCTACC | AAGTAGAAAAGCGCGACCAC |
| BCL2 | AATGACCACCTAGAGCCTTG | GGTCATTTCCGACTGAAGAG |
| CASPASE-3 | CCGAGGCACAGAATTGGACT | TCGCCAGGAATAGTAACCAGG |
| C-MYC | GATAGTGGAAAACCCGGCTGC | CAGATATCCTCGCTGGGTGC |
| P53 | TTTCACCCTCCAGATCCGTG | TTTATGGCGGGAGGGAGACT |
| SOD1 | TCCATGTCCATCAGTTTGGA | AGTCACATTGCCCAGGTCTC |
| SOD2 | AAGCCATCAAACGCGACTTT | CCTTGTTGAAACCGAGCCAA |
| CAT | ACATGGTCTGGGACTTCTGG | TCATGTGCCTGTGTCCATCT |
| GPX4 | ATTCTCAGCCAAGGACATCG | CCTCATTGAGAGGCCACATT |
| NRF2 | CATAGCAGAGCCCAGTACCA | CACGGTGGTCTTGGTTGAAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, J.; Guo, M.; Wang, X.; Wang, R.; Xi, G.; An, L.; Tian, J.; Chu, M. A Redesigned Method for CNP-Synchronized In Vitro Maturation Inhibits Oxidative Stress and Apoptosis in Cumulus-Oocyte Complexes and Improves the Developmental Potential of Porcine Oocytes. Genes 2023, 14, 1885. https://doi.org/10.3390/genes14101885
Lu J, Guo M, Wang X, Wang R, Xi G, An L, Tian J, Chu M. A Redesigned Method for CNP-Synchronized In Vitro Maturation Inhibits Oxidative Stress and Apoptosis in Cumulus-Oocyte Complexes and Improves the Developmental Potential of Porcine Oocytes. Genes. 2023; 14(10):1885. https://doi.org/10.3390/genes14101885
Chicago/Turabian StyleLu, Jinlun, Min Guo, Xiaodong Wang, Rui Wang, Guangyin Xi, Lei An, Jianhui Tian, and Meiqiang Chu. 2023. "A Redesigned Method for CNP-Synchronized In Vitro Maturation Inhibits Oxidative Stress and Apoptosis in Cumulus-Oocyte Complexes and Improves the Developmental Potential of Porcine Oocytes" Genes 14, no. 10: 1885. https://doi.org/10.3390/genes14101885