
ZZ/ZW Sex Chromosomes in the Madagascar Girdled Lizard, Zonosaurus madagascariensis (Squamata: Gerrhosauridae)

{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Species Verification
2.2. Chromosome Preparation and Staining
2.3. Fluorescence In Situ Hybridization with Probes for rDNA Loci and Telomeric Repeats
2.4. Comparative Genomic Hybridization
2.5. Microscopy and Image Analyses
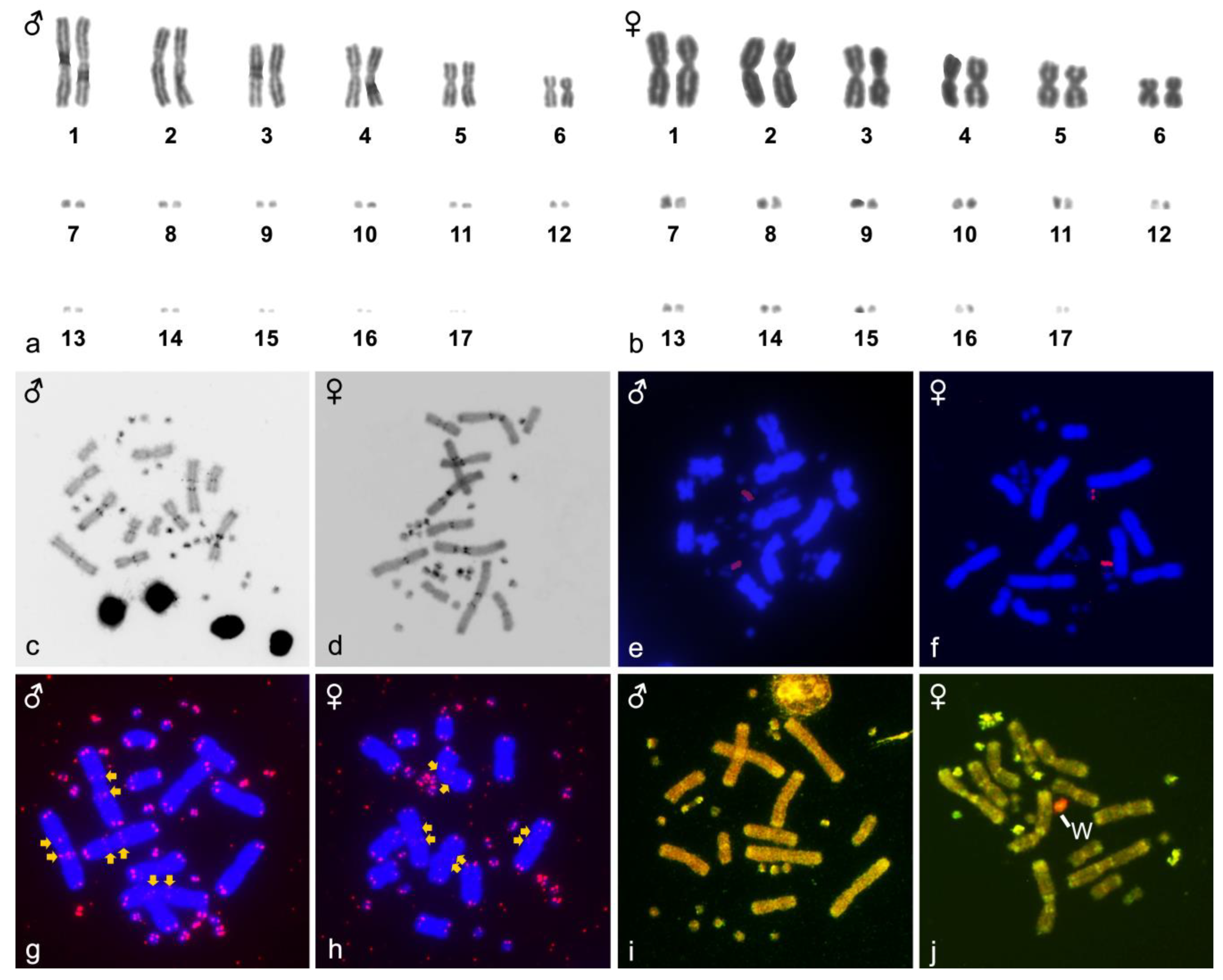
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bull, J.J. Evolution of Sex Determining Mechanisms; Benjamin/Cummings Publishing Company: Menlo Park, CA, USA, 1983. [Google Scholar]
- Abbott, J.K.; Nordén, A.K.; Hansson, B. Sex chromosome evolution: Historical insights and future perspectives. Proc. R. Soc. Lond. B Biol. Sci. 2017, 284, 20162806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capel, B. Vertebrate sex determination: Evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 2017, 18, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Stöck, M.; Rovatsos, M.; Bullejos, M.; Herpin, A.; Jeffries, D.L.; Peichel, C.L.; Perrin, N.; Valenzuela, N.; Johnson Pokorná, M. Expanding the classical paradigm: What we have learnt from vertebrates about sex chromosome evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200097. [Google Scholar] [CrossRef] [PubMed]
- Johnson Pokorná, M.; Kratochvíl, L. What was the ancestral sex-determining mechanism in amniote vertebrates? Biol. Rev. Camb. Philos. Soc. 2016, 91, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Gamble, T.; Rovatsos, M. Sex chromosome evolution among amniotes: Is the origin of sex chromosomes non-random? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200108. [Google Scholar] [CrossRef]
- Cortez, D.; Marin, R.; Toledo-Flores, D.; Froidevaux, L.; Liechti, A.; Waters, P.D.; Grützner, F.; Kaessmann, H. Origins and functional evolution of Y chromosomes across mammals. Nature 2014, 508, 488–493. [Google Scholar] [CrossRef]
- Rovatsos, M.; Pokorná, M.; Altmanová, M.; Kratochvíl, L. Cretaceous park of sex determination: Sex chromosomes are conserved across iguanas. Biol. Lett. 2014, 10, 20131093. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukić, J.; Lymberakis, P.; Kratochvíl, L. Evolutionary stability of sex chromosomes in snakes. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20151992. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.; Vukić, J.; Altmanová, M.; Johnson Pokorná, M.; Moravec, J.; Kratochvíl, L. Conservation of sex chromosomes in lacertid lizards. Mol. Ecol. 2016, 25, 3120–3126. [Google Scholar] [CrossRef]
- Rovatsos, M.; Rehák, I.; Velenský, P.; Kratochvíl, L. Shared ancient sex chromosomes in varanids, beaded lizards, and alligator lizards. Mol. Biol. Evol. 2019, 36, 1113–1120. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, J.; Bachtrog, D.; An, N.; Huang, Q.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1246338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augstenová, B.; Pensabene, E.; Veselý, M.; Kratochvíl, L.; Rovatsos, M. Are geckos special in sex determination? Independently evolved differentiated ZZ/ZW sex chromosomes in carphodactylid geckos. Genome Biol. Evol. 2021, 13, evab119. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, B.J.; Keating, S.E.; Nielsen, S.V.; Scantlebury, D.P.; Daza, J.D.; Gamble, T. Chromosome-level genome assembly reveals dynamic sex chromosomes in neotropical leaf-litter geckos (Sphaerodactylidae: Sphaerodactylus). J. Hered. 2022, 113, 272–287. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. (Eds.) The Reptile Database. 2022. Available online: http://www.reptile-database.org (accessed on 11 April 2022).
- Bates, M.F.; Tolley, K.A.; Edwards, S.; Davids, Z.; Da Silva, J.M.; Branch, W.R. A molecular phylogeny of the African plated lizards, genus Gerrhosaurus Wiegmann, 1828 (Squamata: Gerrhosauridae), with the description of two new genera. Zootaxa 2013, 3750, 465. [Google Scholar] [CrossRef] [Green Version]
- Bezy, R.L. Karyotypic variation and evolution of the lizards in the family Xantusiidae. Los Angeles Cty. Mus. Contr. Sci. 1972, 227, 1–29. [Google Scholar] [CrossRef]
- Hardy, G.S. The karyotypes of two scincid lizards, and their bearing on relationships in genus Leiolopisma and its relatives (Scincidae: Lygosominae). N. Z. J. Zool. 1979, 6, 609–612. [Google Scholar] [CrossRef]
- Odierna, G.; Olmo, E. Chromosomal evolution and DNA of cordylid lizards. Herpetologica 1980, 36, 311–316. [Google Scholar]
- De Smet, W. Description of the orcein stained karyotypes of 36 lizard species (Lacertilia, Reptilia) belonging to the families Teiidae, Scincidae, Lacertidae, Cordylidae and Varanidae (Autarchoglossa). Acta Zool. Pathol. Antverp. 1981, 76, 73–118. [Google Scholar]
- Hass, C.A.; Hedges, S.B. Karyotype of the Cuban lizard Cricosaura typica and its implications for xantusiid phylogeny. Copeia 1992, 1992, 563. [Google Scholar] [CrossRef]
- Caputo, V.; Odierna, G.; Aprea, G. A chromosomal study of Eumeces and Scincus, primitive members of the Scincidae (Reptilia, Squamata). Bolletino Zool. 1994, 61, 155–162. [Google Scholar] [CrossRef]
- Odierna, G.; Canapa, A.; Andreone, F.; Aprea, G.; Barucca, M.; Capriglione, T.; Olmo, E. A phylogenetic analysis of Cordyliformes (Reptilia: Squamata): Comparison of molecular and karyological data. Mol. Phylogenet. Evol. 2002, 23, 37–42. [Google Scholar] [CrossRef]
- Giovannotti, M.; Caputo, V.; O’Brien, P.; Lovell, F.; Trifonov, V.; Cerioni, P.N.; Olmo, E.; Ferguson-Smith, M.; Rens, W. Skinks (Reptilia: Scincidae) have highly conserved karyotypes as revealed by chromosome painting. Cytogenet. Genome Res. 2009, 127, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Kostmann, A.; Augstenová, B.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetically elusive sex chromosomes in scincoidean lizards. Int. J. Mol. Sci. 2021, 22, 8670. [Google Scholar] [CrossRef] [PubMed]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. Proc. R. Soc. Lond. B Biol. Sci. 2021, 288, 20202139. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Guarino, F.M.; Odierna, G. Lizards as model organisms of sex chromosome evolution: What we really know from a systematic distribution of available data? Genes 2021, 12, 1341. [Google Scholar] [CrossRef]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. First report of sex chromosomes in plated lizards (Squamata: Gerrhosauridae). Sex. Dev. 2020, 14, 60–65. [Google Scholar] [CrossRef]
- Castiglia, R.; Bezerra, A.; Flores-Villela, O.; Annesi, F.; Muñoz, A.; Gornung, E. Comparative cytogenetics of two species of ground skinks: Scincella assata and S. cherriei (Squamata: Scincidae: Lygosominae) from Chiapas, Mexico. Acta Herpetol. 2013, 8, 69–73. [Google Scholar]
- Patawang, I.; Chuaynkern, Y.; Supanuam, P.; Maneechot, N.; Pinthong, K.; Tanomtong, A. Cytogenetics of the skinks (Reptilia, Scincidae) from Thailand; IV: Newly investigated karyotypic features of Lygosoma quadrupes and Scincella melanosticta. Caryologia 2018, 71, 29–34. [Google Scholar] [CrossRef]
- Cornejo-Páramo, P.; Dissanayake, D.; Lira-Noriega, A.; Martínez-Pacheco, M.L.; Acosta, A.; Ramírez-Suástegui, C.; Méndez-De-La-Cruz, F.R.; Székely, T.; Urrutia, A.O.; Georges, A.; et al. Viviparous reptile regarded to have temperature-dependent sex determination has old XY chromosomes. Genome Biol. Evol. 2020, 12, 924–930. [Google Scholar] [CrossRef]
- Nielsen, S.V.; Pinto, B.J.; Guzmán-Méndez, I.A.; Gamble, T. First report of sex chromosomes in night lizards (Scincoidea: Xantusiidae). J. Hered. 2020, 111, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Kratochvíl, L.; Vukić, J.; Červenka, J.; Kubička, L.; Johnson Pokorná, M.; Kukačková, D.; Rovatsos, M.; Piálek, L. Mixed-sex offspring produced via cryptic parthenogenesis in a lizard. Mol. Ecol. 2020, 29, 4118–4127. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Endow, S. Polytenization of the ribosomal genes on the X and Y chromosomes of Drosophila melanogaster. Genetics 1982, 100, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.T.; Marchal, J.A.; Romero-Fernández, I.; Fernandez, F.J.; Giagia-Athanosopoulou, E.B.; Sánchez, A. Rapid, independent, and extensive amplification of telomeric repeats in pericentromeric regions in karyotypes of arvicoline rodents. Chromosome Res. 2011, 19, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Alföldi, J.; Di Palma, F.; Grabherr, M.; Williams, C.; Kong, L.; Mauceli, E.; Russell, P.; Lowe, C.B.; Glor, R.E.; Jaffe, J.D.; et al. The genome of the green anole lizard and a comparative analysis with birds and mammals. Nature 2011, 477, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Capanna, E.; Castiglia, R. Chromosomes and speciation in Mus musculus domesticus. Cytogenet. Genome Res. 2004, 105, 375–384. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Velenský, P.; Sánchez Baca, A.; Kratochvíl, L. Evolution of karyotypes in chameleons. Genes 2017, 8, 382. [Google Scholar] [CrossRef] [Green Version]
- Augstenová, B.; Pensabene, E.; Kratochvíl, L.; Rovatsos, M. Cytogenetic evidence for sex chromosomes and karyotype evolution in anguimorphan lizards. Cells 2021, 10, 1612. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kostmann, A.; Altmanová, M.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetic analysis did not reveal differentiated sex chromosomes in ten species of boas and pythons (Reptilia: Serpentes). Genes 2019, 10, 934. [Google Scholar] [CrossRef] [Green Version]
- Slotkin, R.K.; Martienssen, R. Transposable elements and the epigenetic regulation of the genome. Nat. Rev. Genet. 2007, 8, 272–285. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.G.; Jones, B.C.; Jiang, N.; Chang, C.; Hosier, S.; Wickremesinghe, P.; Garcia, M.; Hartnett, D.A.; Burhenn, L.; Neretti, N.; et al. Chromatin-modifying genetic interventions suppress age-associated transposable element activation and extend life span in Drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 11277–11282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allshire, R.C.; Madhani, H.D. Ten principles of heterochromatin formation and function. Nat. Rev. Mol. 2018, 19, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Kratochvíl, L.; Altmanová, M.; Johnson Pokorná, M. Interstitial telomeric motifs in squamate reptiles: When the exceptions outnumber the rule. PLoS ONE 2015, 10, e0134985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente, L.; Mazzoleni, S.; Pensabene Bellavia, E.; Augstenová, B.; Auer, M.; Praschag, P.; Protiva, T.; Velenský, P.; Wagner, P.; Fritz, U.; et al. Interstitial telomeric repeats are rare in turtles. Genes 2020, 11, 657. [Google Scholar] [CrossRef]
- Park, V.M.; Gustashaw, K.M.; Wathen, T.M. The presence of interstitial telomeric sequences in constitutional chromosome abnormalities. Am. J. Hum. Genet. 1992, 50, 914–923. [Google Scholar]
- Melek, M.; Shippen, D.E. Chromosome healing: Spontaneous and programmed de novo telomere formation by telomerase. BioEssays 1996, 18, 301–308. [Google Scholar] [CrossRef]
- Vermeesch, J.R.; Petit, P.; Speleman, F.; Devriendt, K.; Fryns, J.P.; Marynen, P. Interstitial telomeric sequences at the junction site of a jumping translocation. Hum. Genet. 1997, 99, 735–737. [Google Scholar] [CrossRef]
- Shay, J.R.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Porter, C.A.; Hamilton, M.J.; Sites, J.W., Jr.; Baker, R.J. Location of ribosomal DNA in chromosomes of squamate reptiles: Systematic and evolutionary implications. Herpetologica 1991, 47, 271–280. [Google Scholar]
- Porter, C.A.; Haiduk, M.W.; de Queiroz, K. Evolution and phylogenetic significance of ribosomal gene location in chromosomes of squamate reptiles. Copeia 1994, 1994, 302–313. [Google Scholar] [CrossRef]
- Matsubara, K.; Kumazawa, Y.; Ota, H.; Nishida, C.; Matsuda, Y. Karyotype analysis of four blind snake species (Reptilia: Squamata: Scolecophidia) and karyotypic changes in serpentes. Cytogenet. Genome Res. 2019, 157, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Clemente, L.; Mazzoleni, S.; Pensabene, E.; Protiva, T.; Wagner, P.; Fritz, U.; Kratochvíl, L.; Rovatsos, M. Cytogenetic analysis of the Asian box turtles of the genus Cuora (Testudines, Geoemydidae). Genes 2021, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Sex is determined by XX/XY sex chromosomes in Australasian side-necked turtles (Testudines: Chelidae). Sci. Rep. 2020, 10, 4276. [Google Scholar] [CrossRef] [Green Version]
- Badenhorst, D.; Stanyon, R.; Engstrom, T.; Valenzuela, N. A ZZ/ZW microchromosome system in the spiny softshell turtle, Apalone spinifera, reveals an intriguing sex chromosome conservation in Trionychidae. Chromosome Res. 2013, 21, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Montiel, E.E.; Badenhorst, D.; Lee, L.; Valenzuela, N. Evolution and dosage compensation of nucleolar organizing regions (NORs) mediated by mobile elements in turtles with female (ZZ/ZW) but not with male (XX/XY) heterogamety. J. Evol. Biol. 2022, 35, 1709–1720. [Google Scholar] [CrossRef]
- Traut, W.; Sahara, K.; Otto, T.D.; Marec, F. Molecular differentiation of sex chromosomes probed by comparative genomic hybridization. Chromosoma 1999, 108, 173–180. [Google Scholar] [CrossRef]
- Traut, W.; Eickhoff, U.; Schorch, J.-C. Identification and analysis of sex chromosomes by comparative genomic hybridization (CGH). Methods Cell Sci. 2001, 23, 155–161. [Google Scholar] [CrossRef]
- Matsubara, K.; Knopp, T.; Sarre, S.D.; Georges, A.; Ezaz, T. Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed worm-lizard (Aprasia parapulchella, Pygopodidae, Squamata). Mol. Cytogenet. 2013, 6, 60. [Google Scholar] [CrossRef] [Green Version]
- Pokorná, M.; Rens, W.; Rovatsos, M.; Kratochvíl, L. A ZZ/ZW sex chromosome system in the thick-tailed gecko (Underwoodisaurus milii; Squamata: Gekkota: Carphodactylidae), a member of the ancient gecko lineage. Cytogenet. Genome Res. 2014, 142, 190–196. [Google Scholar] [CrossRef]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Mixed-up sex chromosomes: Identification of sex chromosomes in the X1X1X2X2/X1X2Y system of the legless lizards of the genus Lialis (Squamata: Gekkota: Pygopodidae). Cytogenet. Genome Res. 2016, 149, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Augstenová, B.; Kratochvíl, L. Cytogenetics of the Javan file snake (Acrochordus javanicus) and the evolution of snake sex chromosomes. J. Zool. Syst. Evol. Res. 2017, 56, 117–125. [Google Scholar] [CrossRef]
- Viana, P.F.; Ezaz, T.; de Bello Cioffi, M.; Jackson Almeida, B.; Feldberg, E. Evolutionary insights of the ZW sex chromosomes in snakes: A new chapter added by the Amazonian puffing snakes of the genus Spilotes. Genes 2019, 10, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Female heterogamety in Madagascar chameleons (Squamata: Chamaeleonidae: Furcifer): Differentiation of sex and neo-sex chromosomes. Sci. Rep. 2015, 5, 13196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Wang, C.; Tang, X.; Xin, Y.; Yue, F.; Yan, X.; Liu, B.; An, B.; Wang, X.; Chen, Q. Identification of sex chromosomes by means of comparative genomic hybridization in a lizard, Eremias multiocellata. Zool. Sci. 2015, 32, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Sarre, S.D.; Georges, A.; Matsuda, Y.; Graves, J.A.M.; Ezaz, T. Highly differentiated ZW sex microchromosomes in the Australian Varanus species evolved through rapid amplification of repetitive sequences. PLoS ONE 2014, 9, e95226. [Google Scholar] [CrossRef]
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Graves, J. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- Shine, R.; Elphick, M.J.; Donnellan, S. Co-occurrence of multiple, supposedly incompatible modes of sex determination in a lizard population. Ecol. Lett. 2002, 5, 486–489. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostmann, A.; Kratochvíl, L.; Rovatsos, M. ZZ/ZW Sex Chromosomes in the Madagascar Girdled Lizard, Zonosaurus madagascariensis (Squamata: Gerrhosauridae). Genes 2023, 14, 99. https://doi.org/10.3390/genes14010099
Kostmann A, Kratochvíl L, Rovatsos M. ZZ/ZW Sex Chromosomes in the Madagascar Girdled Lizard, Zonosaurus madagascariensis (Squamata: Gerrhosauridae). Genes. 2023; 14(1):99. https://doi.org/10.3390/genes14010099
Chicago/Turabian StyleKostmann, Alexander, Lukáš Kratochvíl, and Michail Rovatsos. 2023. "ZZ/ZW Sex Chromosomes in the Madagascar Girdled Lizard, Zonosaurus madagascariensis (Squamata: Gerrhosauridae)" Genes 14, no. 1: 99. https://doi.org/10.3390/genes14010099