
The Complete Mitochondrial Genome and Gene Arrangement of the Enigmatic Scaphopod Pictodentalium vernedei
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Isolation
2.2. Mitochondrial Genome Sequencing and Assembling
2.3. Annotation and Characterization of Scaphopod Mitochondrial Genome
2.4. Phylogenetic Placement
3. Results and Discussion
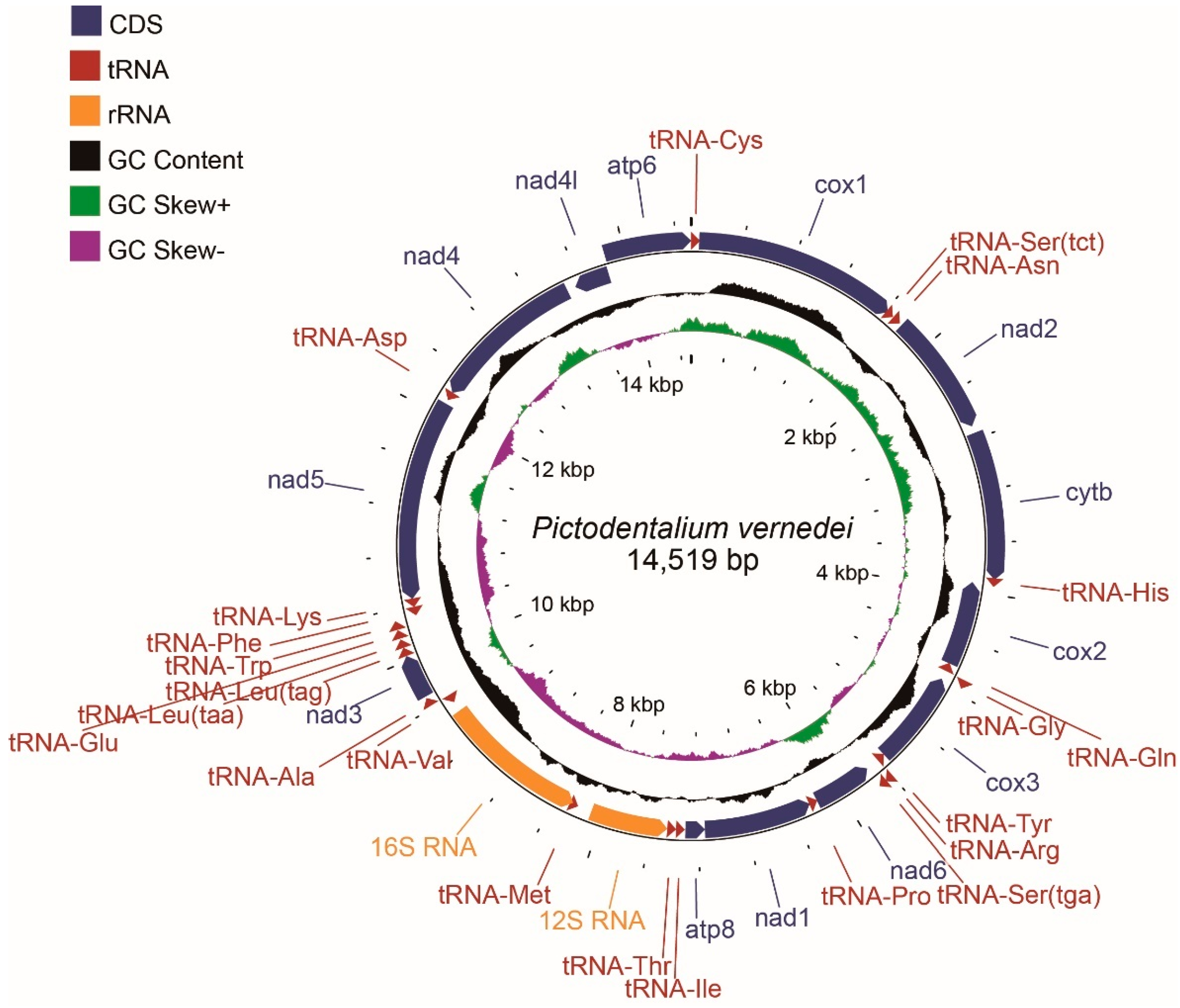
3.1. General Features of Mitochondrial Genome
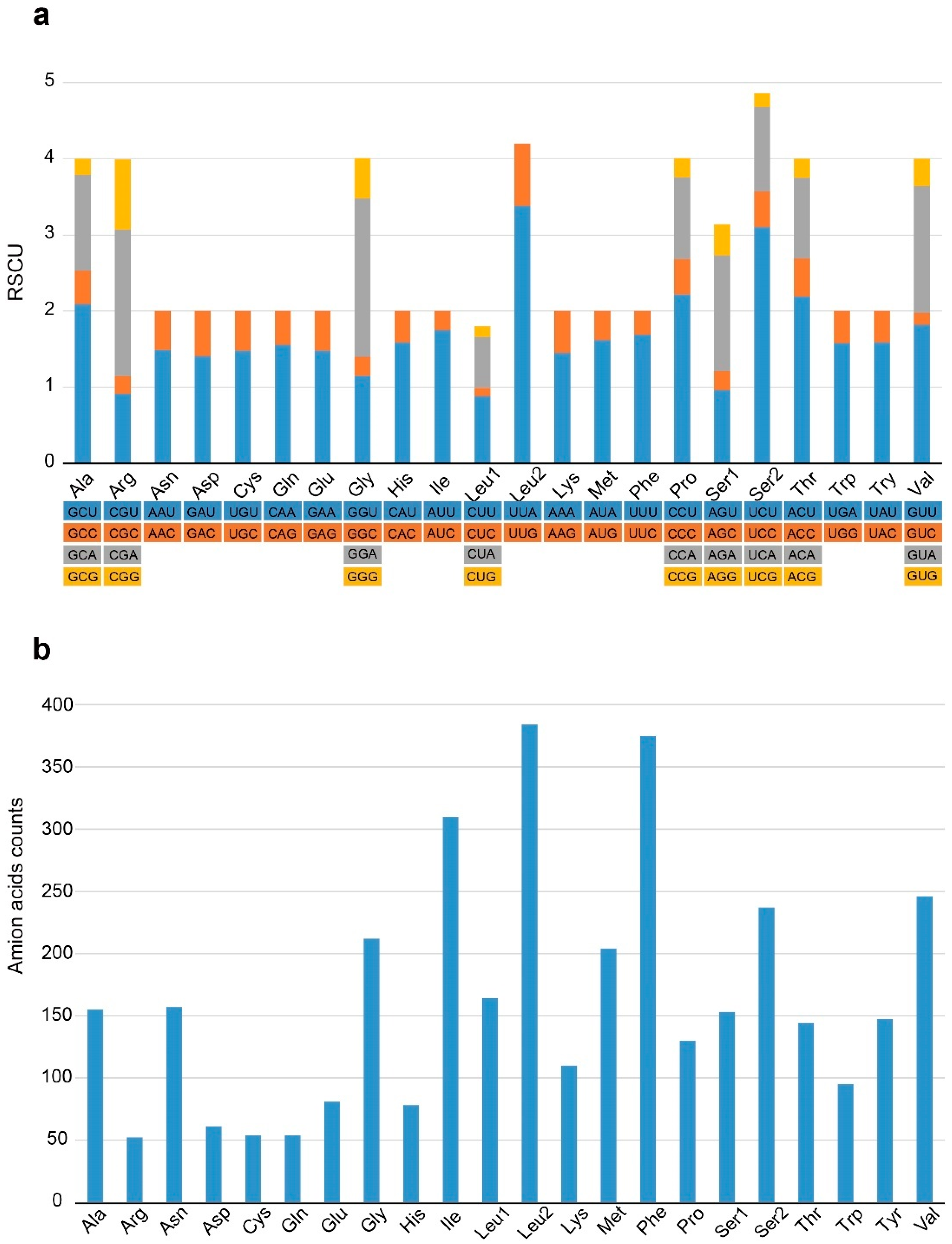
3.2. Protein-Coding Genes (PCGs)
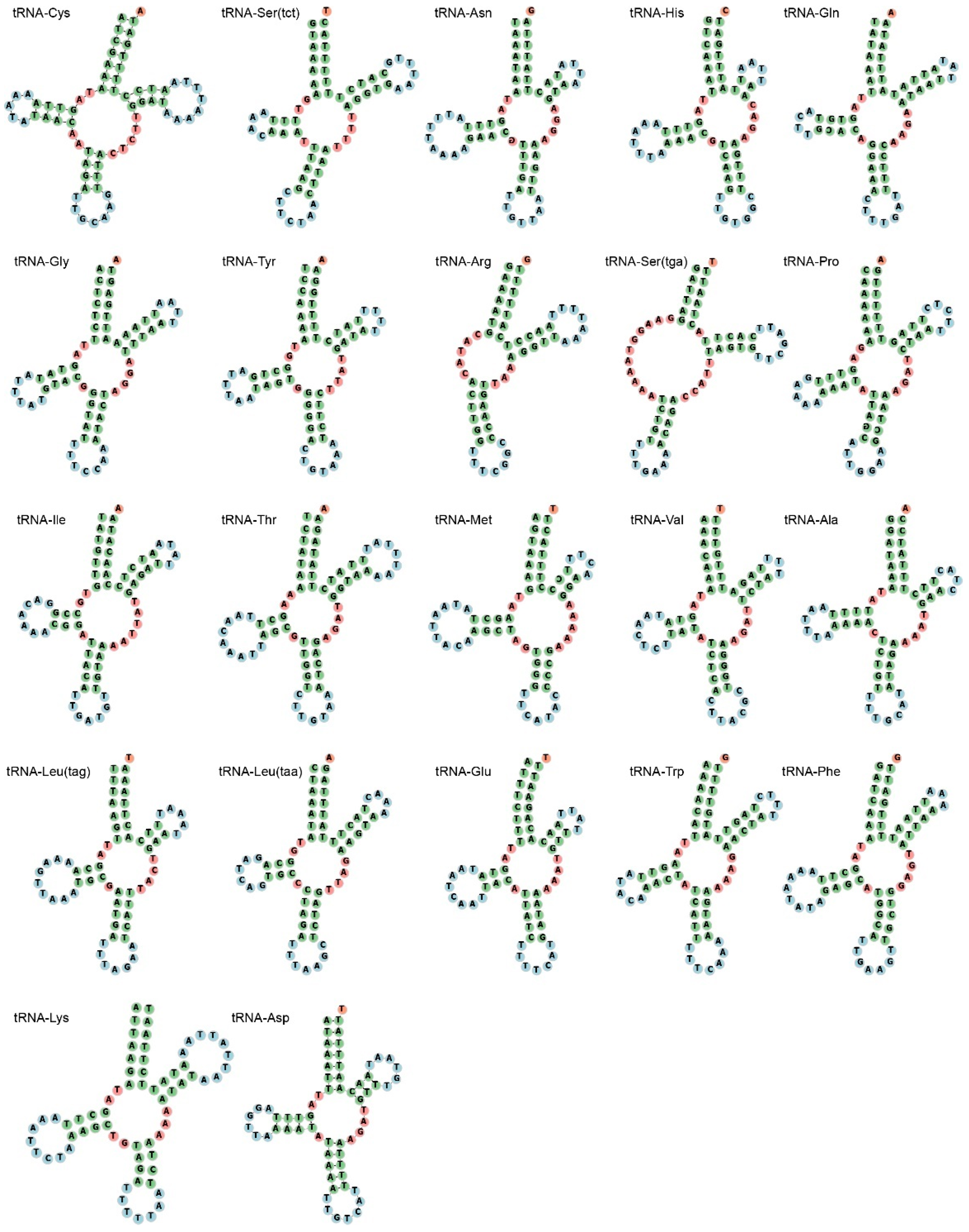
3.3. Ribosomal RNAs and Transfer RNAs
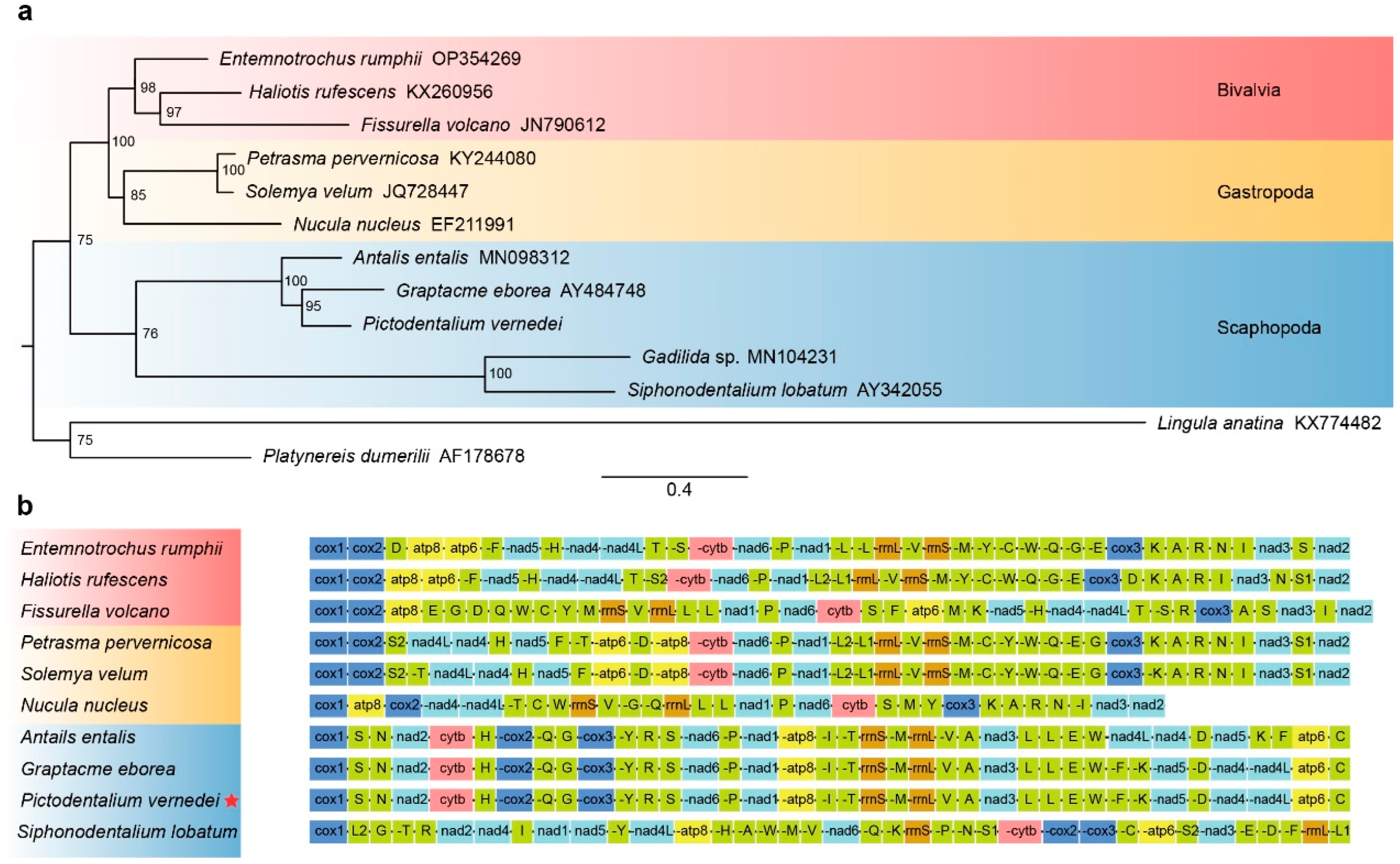
3.4. Phylogenetic Placement
3.5. Mitochondrial Gene Arrangement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BS | Bootstrap |
| DHU | Di-hydroxy uridine |
| DUI | doubly uniparental inheritance |
| ML | maximum likelihood |
| PCG | protein coding gene |
| RSCU | relative synonymous codon usage |
References
- Ghiselli, F.; Gomes-dos-Santos, A.; Adema, C.M.; Lopes-Lima, M.; Sharbrough, J.; Boore, J.L. Molluscan mitochondrial genomes break the rules. Philos. Trans. R. Soc. B 2021, 376, 20200159. [Google Scholar] [CrossRef] [PubMed]
- Ponder, W.F.; Lindberg, D.R.; Ponder, J.M. Biology and Evolution of the Mollusca, 1st ed.; CRC Press: Boca Raton, FL, USA, 2020; Volume 1, p. 924. [Google Scholar]
- Anderson, M.P.; Fenberg, P.B.; Griffiths, H.J.; Linse, K. Macrobenthic mollusca of the Prince Gustav Channel, eastern Antarctic Peninsula: An area undergoing colonisation. Front. Mar. Sci. 2021, 8, 771369. [Google Scholar] [CrossRef]
- Kocot, K.M.; Cannon, J.T.; Todt, C.; Citarella, M.R.; Kohn, A.B.; Meyer, A.; Santos, S.R.; Schander, C.; Moroz, L.L.; Lieb, B. Phylogenomics reveals deep molluscan relationships. Nature 2011, 477, 452–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocot, K.M. Recent advances and unanswered questions in deep molluscan phylogenetics. Am. Malacol. Bull. 2013, 31, 195–208. [Google Scholar] [CrossRef]
- Smith, S.A.; Wilson, N.G.; Goetz, F.E.; Feehery, C.; Andrade, S.; Rouse, G.W.; Giribet, G.; Dunn, C.W. Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature 2011, 480, 364–367. [Google Scholar] [CrossRef]
- Reynolds, P.D.; Okusu, A. Phylogenetic relationships among families of the Scaphopoda (Mollusca). Zool. J. Linn. Soc. 1999, 126, 131–154. [Google Scholar] [CrossRef]
- Reynolds, P.D. The phylogeny and classification of Scaphopoda (Mollusca): An assessment of current resolution and cladistic reanalysis. Zool. Scr. 1997, 26, 13–21. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M.J. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Hatzoglou, E.; Rodakis, G.C.; Lecanidou, R. Complete sequence and gene organization of the mitochondrial genome of the land snail Albinaria coerulea. Genetics 1995, 140, 1353–1366. [Google Scholar] [CrossRef]
- Kurabayashi, A.; Ueshima, R. Evolution, Complete sequence of the mitochondrial DNA of the primitive opisthobranch gastropod Pupa strigosa: Systematic implication of the genome organization. Mol. Biol. Evol. 2000, 17, 266–277. [Google Scholar] [CrossRef]
- White, T.R.; Conrad, M.M.; Tseng, R.; Balayan, S.; Golding, R.; de Frias Martins, A.M.; Dayrat, B.A. Ten new complete mitochondrial genomes of pulmonates (Mollusca: Gastropoda) and their impact on phylogenetic relationships. BMC Evol. Biol. 2011, 11, 295. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Li, Y.; Kocot, K.M.; Yang, Y.; Qi, L.; Li, Q.; Halanych, K.M. Mitogenomics reveals phylogenetic relationships of Arcoida (Mollusca, Bivalvia) and multiple independent expansions and contractions in mitochondrial genome size. Mol. Phylogenet. Evol. 2020, 150, 106857. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, P.; Zhang, Z.; Li, C.; Liu, Y.; Chen, Y.; Wang, J.; Wang, H.; Song, H. The Complete Mitochondrial Genome of Entemnotrochus rumphii, a Living Fossil for Vetigastropoda (Mollusca: Gastropoda). Genes 2022, 13, 2061. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, H.; Steiner, G. Evolution, The complete sequence and gene organization of the mitochondrial genome of the gadilid scaphopod Siphonondentalium lobatum (Mollusca). Mol. Phylogenet. Evol. 2004, 31, 605–617. [Google Scholar] [CrossRef] [PubMed]
- Garey, J.R.; Wolstenholme, D.R. Platyhelminth mitochondrial DNA: Evidence for early evolutionary origin of a tRNAserAGN that contains a dihydrouridine arm replacement loop, and of serine-specifying AGA and AGG codons. J. Mol. Evol. 1989, 28, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Runnegar, B.; Pojeta, J., Jr. Molluscan Phylogeny: The Paleontological Viewpoint: The early Paleozoic fossil record shows how living and extinct molluscan classes originated and diversified. Science 1974, 186, 311–317. [Google Scholar] [CrossRef]
- Pojeta, J., Jr. The paleontology of rostroconch mollusks and the early history of the Phylum Mollusca. US Geol. Surv. Prof. Pap. 1976, 968, 1–54. [Google Scholar]
- De Lacaze Duthiers, H. Histoire de L’organisation, du Développement, des Moeurs et des Rapports Zoologiques du Dentale; Librairie de Victor Masson: Paris, France, 1858. [Google Scholar]
- Waller, T.R.; Johnston, P.; Haggart, J. Origin of the molluscan class Bivalvia and a phylogeny of major groups. Bivalves Eon Evol. 1998, 1, 5. [Google Scholar]
- Hatschek, B. Lehrbuch der Zoologie, eine morphologische Ubersicht des Thierreiches zur Einfuhrung in das Studium dieser Wissenschaft; Gustav Fischer Verlag: Jena, Germany, 1888. [Google Scholar]
- Steiner, G.; Dreyer, H. Molecular phylogeny of Scaphopoda (Mollusca) inferred from 18S rDNA sequences: Support for a Scaphopoda–Cephalopoda clade. Zool. Scr. 2003, 32, 343–356. [Google Scholar] [CrossRef]
- Plate, L. Über den Bau und die Verwandtschaftsbeziehungen der Solenoconchen. Zool. Jahrbücher Der Anat. 1892, 5, 301–386. [Google Scholar]
- Bronn, H.G. Dr. HG Bronn’s Klassen und Ordnungen des Thier-Reichs, wissenschaftlich dargestellt in Wort und Bild. Dritter Band. Mollusca (Weichthiere); CF Winter’sche-Verlagshandlung: Leipzig, Germany, 1894. [Google Scholar]
- Grande, C.; Templado, J.; Zardoya, R. Evolution of gastropod mitochondrial genome arrangements. BMC Ecol. Evol. 2008, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Gissi, C.; Iannelli, F.; Pesole, G. Evolution of the mitochondrial genome of Metazoa as exemplified by comparison of congeneric species. Heredity 2008, 101, 301–320. [Google Scholar] [CrossRef] [Green Version]
- Guerra, D.; Bouvet, K.; Breton, S. Mitochondrial gene order evolution in Mollusca: Inference of the ancestral state from the mtDNA of Chaetopleura apiculata (Polyplacophora, Chaetopleuridae). Mol. Phylogenet. Evol. 2018, 120, 233–239. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Subclass | Order | Species | Accession Number |
|---|---|---|---|---|---|
| Mollusca | Gastropoda | Vetigastropoda | Pleurotomariida | Entemnotrochus rumphii | OP354269 |
| Lepetellida | Haliotis rufescens | KX260956 | |||
| Fissurella volcano | JN790612 | ||||
| Bivalvia | Protobranchia | Solemyida | Petrasma pervernicosa | KY244080 | |
| Solemya velum | JQ728447 | ||||
| Nuculida | Nucula nucleus | EF211991 | |||
| Scaphopoda | Dentaliida | Antalis entalis | MN098312 | ||
| G. eborea | AY484748 | ||||
| P. vernedei | This study | ||||
| Gadilida | Siphonodentalium lobatum | AY342055 | |||
| Gadilida sp. | MN104231 | ||||
| Annelida | Polychaeta | Errantia | Phyllodocida | Platynereis dumerilii | AF178678 |
| Brachiopoda | Lingulata | Lingulida | Lingula anatina | KX774482 |
| Gene Name | Location | Length (bp) | Codon | Anticodon | Intergenic Region | Strand | ||
|---|---|---|---|---|---|---|---|---|
| Start | End | Start | Stop | |||||
| tRNA-Cys | 1 | 68 | 68 | GCA | 0 | H | ||
| cox1 | 63 | 1619 | 1557 | TTG | TAA | −6 | H | |
| tRNA-Ser(tct) | 1606 | 1670 | 65 | TCT | −14 | H | ||
| tRNA-Asn | 1675 | 1740 | 66 | GTT | 4 | H | ||
| nad2 | 1761 | 2700 | 940 | ATG | T | 20 | H | |
| cytb | 2755 | 3888 | 1134 | ATA | TAG | 54 | H | |
| tRNA-His | 3879 | 3940 | 62 | GTG | −10 | H | ||
| cox2 | 3927 | 4616 | 690 | ATG | TAA | −14 | L | |
| tRNA-Gln | 4617 | 4680 | 64 | TTG | 0 | L | ||
| tRNA-Gly | 4687 | 4753 | 67 | TCC | 6 | H | ||
| cox3 | 4756 | 5535 | 780 | ATG | TAA | 2 | L | |
| tRNA-Tyr | 5536 | 5600 | 65 | GTA | 0 | L | ||
| tRNA-Arg | 5599 | 5657 | 59 | TCG | −2 | H | ||
| tRNA-Ser(tga) | 5657 | 5720 | 64 | TGA | −1 | H | ||
| nad6 | 5714 | 6199 | 486 | ATG | TAG | −7 | L | |
| tRNA-Pro | 6200 | 6264 | 65 | TGG | 0 | L | ||
| nad1 | 6265 | 7147 | 883 | ATG | T | 0 | L | |
| atp8 | 7148 | 7306 | 159 | ATG | TAG | 0 | L | |
| tRNA-Ile | 7314 | 7381 | 68 | GAT | 7 | L | ||
| tRNA-Thr | 7384 | 7454 | 71 | TGT | 2 | L | ||
| 12S RNA | 7456 | 8091 | 636 | 1 | L | |||
| tRNA-Met | 8211 | 8276 | 66 | CAT | 119 | L | ||
| 16S RNA | 8259 | 9458 | 1200 | −18 | L | |||
| tRNA-Val | 9549 | 9611 | 63 | TAC | 90 | L | ||
| tRNA-Ala | 9621 | 9683 | 63 | TGC | 9 | H | ||
| nad3 | 9699 | 10,037 | 339 | TTG | TAA | 15 | H | |
| tRNA-Leu(tag) | 10,039 | 10,103 | 65 | TAG | 1 | H | ||
| tRNA-Leu(taa) | 10,105 | 10,169 | 65 | TAA | 1 | H | ||
| tRNA-Glu | 10,179 | 10,243 | 65 | TTC | 9 | H | ||
| tRNA-Trp | 10,248 | 10,313 | 66 | TCA | 4 | H | ||
| tRNA-Phe | 10,322 | 10,389 | 68 | GAA | 8 | L | ||
| tRNA-Lys | 10,390 | 10,458 | 69 | TTT | 0 | L | ||
| nad5 | 10,458 | 12,104 | 1647 | ATT | TAA | −1 | L | |
| tRNA-Asp | 12,143 | 12,207 | 65 | GTC | 38 | L | ||
| nad4 | 12,209 | 13,477 | 1269 | ATT | TAA | 1 | L | |
| nad4l | 13,549 | 13,839 | 291 | ATT | TAA | 71 | L | |
| atp6 | 13,853 | 14,519 | 667 | ATT | T | 13 | H | |
| Species Selected | Size (bp) | A% | T% | G% | C% | A-T Skew | G-C Skew | AT Proportion |
|---|---|---|---|---|---|---|---|---|
| E. rumphii | 15,795 | 35.21 | 29.98 | 14.43 | 20.39 | 0.08023 | −0.17112 | 65.19% |
| H. rufescens | 16,646 | 35.39 | 24.93 | 13.75 | 25.93 | 0.17339 | −0.30719 | 60.32% |
| F. volcano | 17,575 | 25.72 | 35.83 | 26.69 | 11.76 | −0.16431 | 0.38819 | 61.55% |
| P. pervernicosa | 16,554 | 34.45 | 31.84 | 12.64 | 21.07 | 0.03946 | −0.24996 | 66.29% |
| S. velum | 15,660 | 35.53 | 32.58 | 13.14 | 18.74 | 0.04341 | −0.17565 | 68.11% |
| N. nucleus | 13,671 | 32.13 | 31.78 | 15.66 | 20.42 | 0.00549 | −0.13197 | 63.92% |
| A. entalis | 14,869 | 35.30 | 40.41 | 12.39 | 11.90 | −0.06742 | 0.01993 | 75.71% |
| G. eborea | 14,492 | 36.98 | 37.14 | 12.67 | 13.21 | −0.00214 | −0.02081 | 74.12% |
| P. vernedei | 14,519 | 36.46 | 35.45 | 13.24 | 14.85 | 0.01408 | −0.05738 | 71.91% |
| S. lobatum | 13,932 | 32.08 | 36.20 | 18.92 | 12.81 | −0.06036 | 0.19258 | 68.27% |
| Gadilida sp. | 13,790 | 32.08 | 32.82 | 14.35 | 20.74 | −0.01134 | −0.18214 | 64.91% |
| P. dumerilii | 15,619 | 31.22 | 32.92 | 15.41 | 20.45 | −0.02646 | −0.14071 | 64.14% |
| L. anatina | 24,876 | 26.28 | 36.53 | 21.29 | 15.90 | −0.16308 | 0.14483 | 62.81% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Wang, Y.; Song, H. The Complete Mitochondrial Genome and Gene Arrangement of the Enigmatic Scaphopod Pictodentalium vernedei. Genes 2023, 14, 210. https://doi.org/10.3390/genes14010210
Zhang T, Wang Y, Song H. The Complete Mitochondrial Genome and Gene Arrangement of the Enigmatic Scaphopod Pictodentalium vernedei. Genes. 2023; 14(1):210. https://doi.org/10.3390/genes14010210
Chicago/Turabian StyleZhang, Tianzhe, Yunan Wang, and Hao Song. 2023. "The Complete Mitochondrial Genome and Gene Arrangement of the Enigmatic Scaphopod Pictodentalium vernedei" Genes 14, no. 1: 210. https://doi.org/10.3390/genes14010210