
Identification of Differentially Expressed Genes in the Longissimus Dorsi Muscle of Luchuan and Duroc Pigs by Transcriptome Sequencing

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Collection of Tissue Samples
2.2. RNA Extraction, Library Construction and Illumina Sequencing
2.3. Gene Ontology and Pathway Enrichment Analysis
2.4. Real–Time Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.5. Vector Construction and Transient Transfection
2.6. Statistical Analysis
3. Results
3.1. Summary of RNA-Seq Data
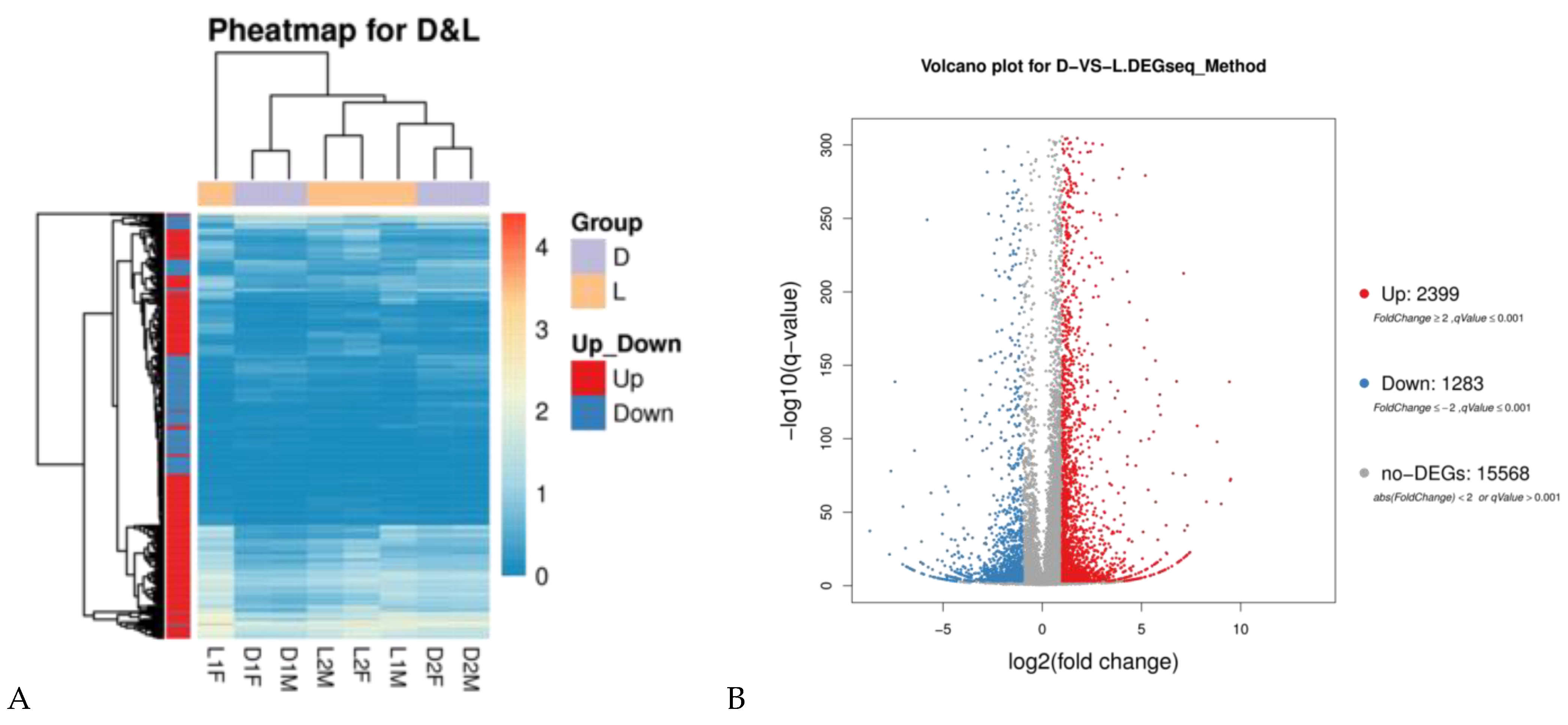
3.2. Identification of Differentially Expressed Genes
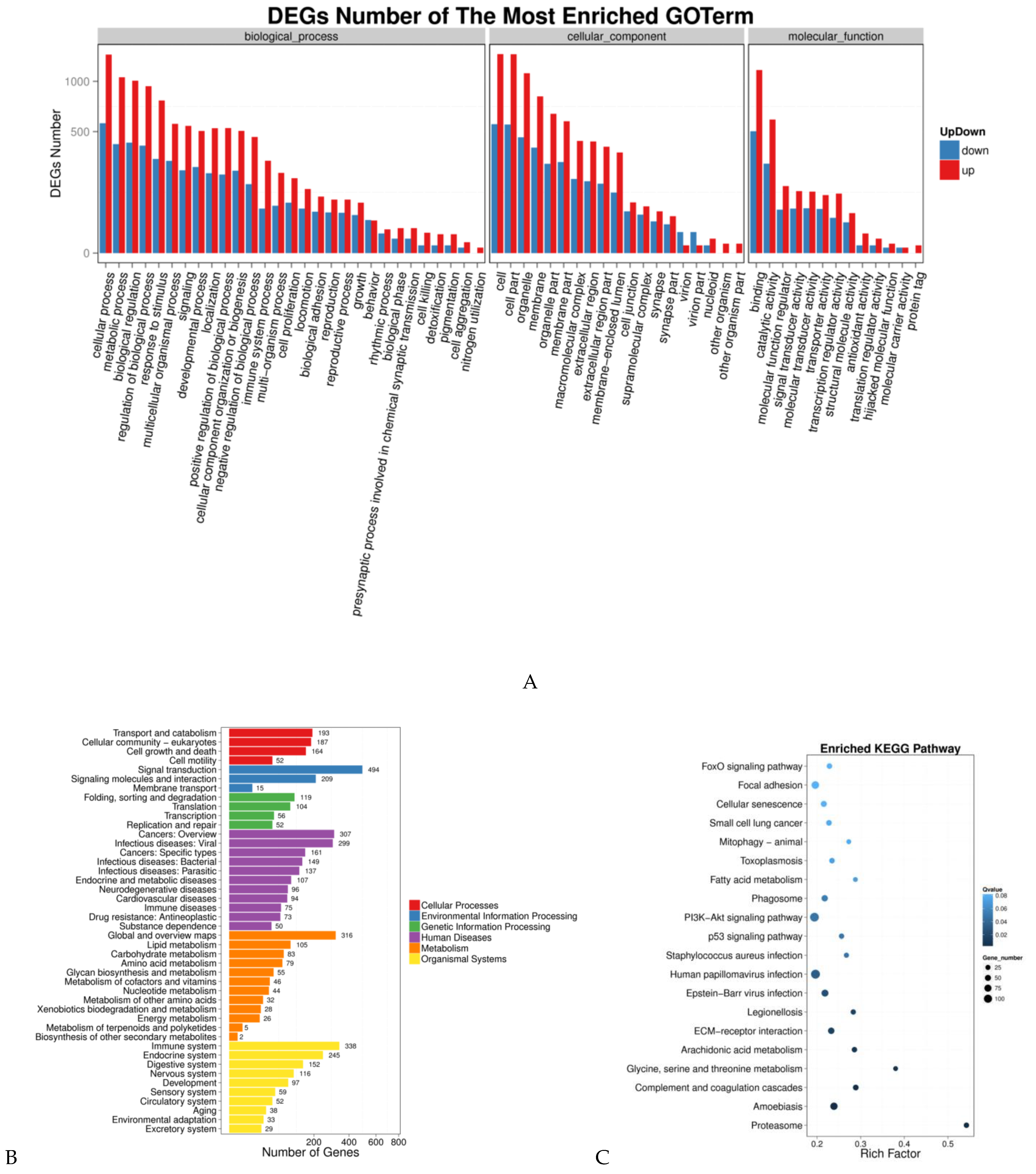
3.3. GO Enrichment and KEGG Pathway Analyses
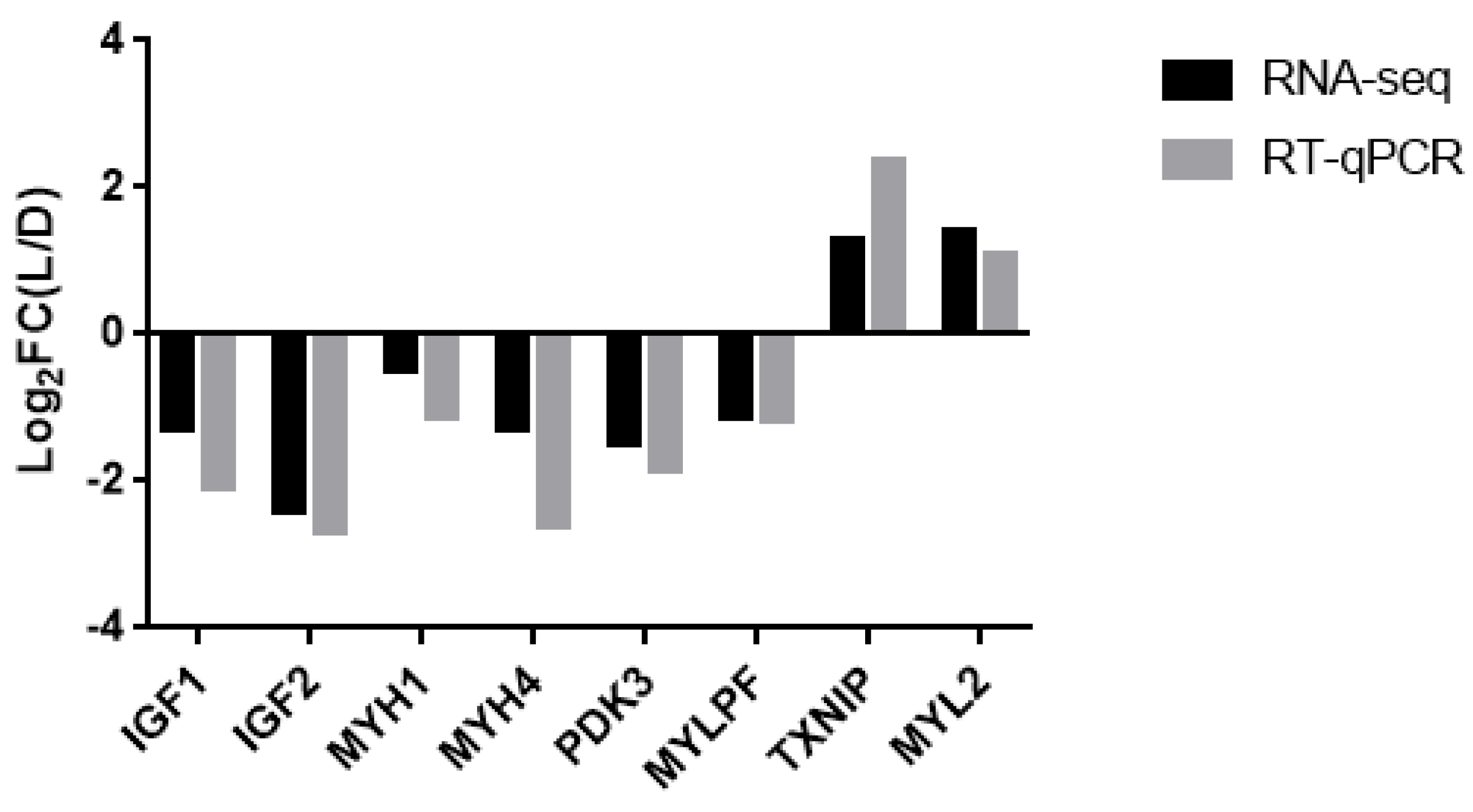
3.4. Verification of the Accuracy of the RNA-Seq Data Using RT-qPCR
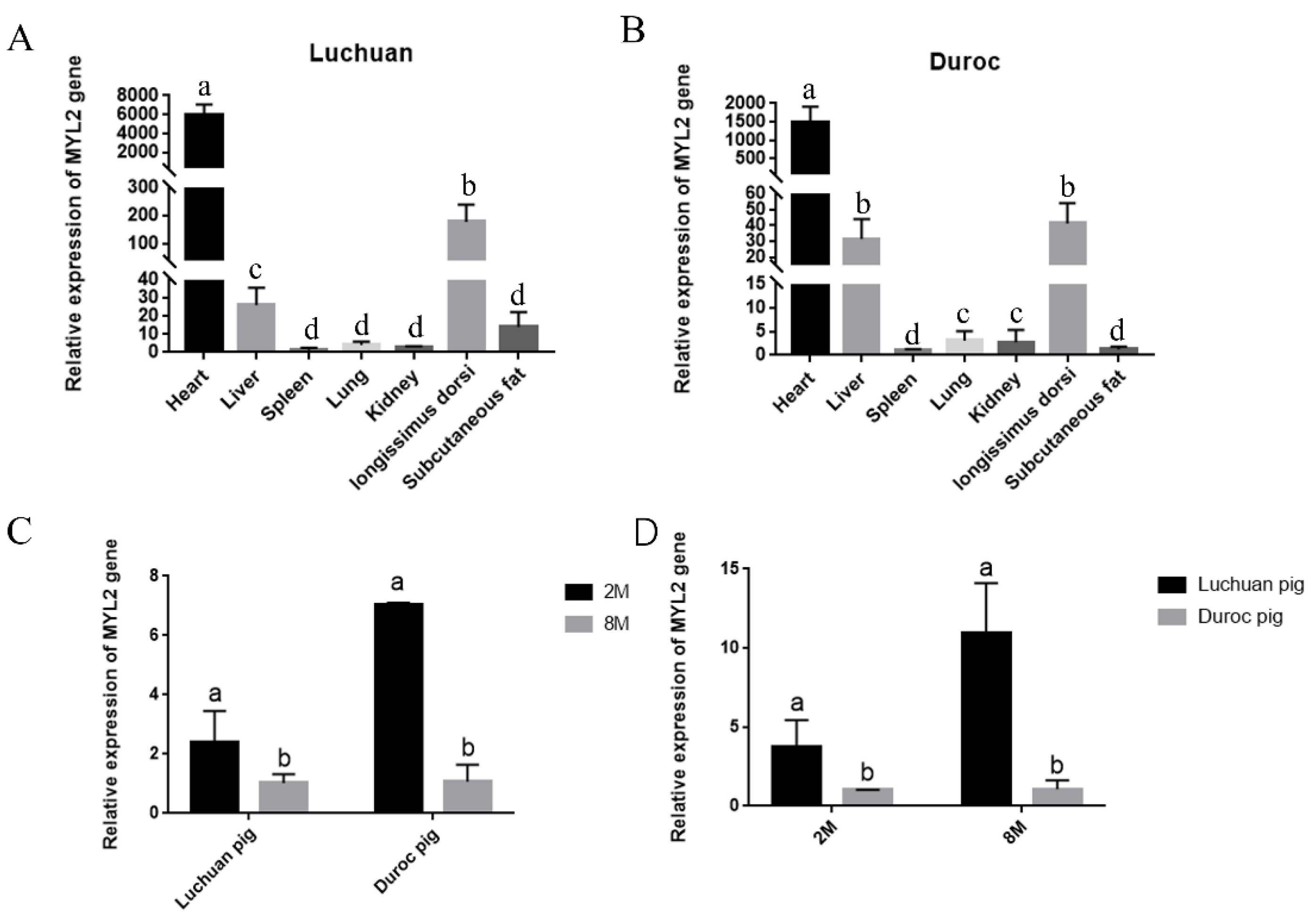
3.5. Analysis of the MYL2 Gene Expression Pattern
3.6. Expression Trend of the MYL2 Gene and Genes Related to Muscle Development in C2C12 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, X.; Mo, D.L.; Zhang, Y.; Guo, W.; Li, A.N.; Chen, Y.S. Progress in research on skeletal muscle growth and development in swine. Chin. Bull. Life Sci. 2011, 23, 37–44. [Google Scholar]
- Nan, L.; Ruan, J.; Yang, S. Regulation of Porcine Muscle Development and Research Progress of Improved Pigs with High Lean Meat Percentage. J. Agric. Biotechnol. 2012, 20, 955–964. [Google Scholar]
- Zhang, Z.; Liao, Q.; Sun, Y.; Pan, T.; Liu, S.; Miao, W.; Li, Y.; Zhou, L.; Xu, G. Lipidomic and Transcriptomic Analysis of the Longissimus Muscle of Luchuan and Duroc Pigs. Front. Nutr. 2021, 8, 667622. [Google Scholar] [CrossRef] [PubMed]
- Edea, Z.; Hong, J.-K.; Jung, J.-H.; Kim, D.-W.; Kim, Y.-M.; Kim, E.-S.; Shin, S.S.; Jung, Y.C.; Kim, K.-S. Detecting selection signatures between Duroc and Duroc synthetic pig populations using high-density SNP chip. Anim. Genet. 2017, 48, 473–477. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, S.M.; Huff-Lonergan, E.; Rowe, L.J.; Kuhlers, D.L.; Jungst, S.B. Selection for lean growth efficiency in Duroc pigs influences pork quality. J. Anim. Sci. 2001, 79, 2075–2085. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-H.; Qin, S.-H.; Liang, W.-Q. Conservation and hybridization of luchuan pigs in Guangxi. Guangxi J. Anim. Husb. 2010, 26, 96–97. [Google Scholar]
- Yu, L.; Tai, L.; Zhang, L.; Chu, Y.; Li, Y.; Zhou, L. Comparative analyses of long non-coding RNA in lean and obese pig. Oncotarget 2017, 8, 41440–41450. [Google Scholar] [CrossRef] [Green Version]
- Wigmore, P.M.; Stickland, N.C. Muscle development in large and small pig fetuses. J. Anat. 1983, 137, 235–245. [Google Scholar]
- Ashmore, C.R.; Addis, P.B.; Doerr, L. Development of Muscle Fibers in the Fetal Pig. J. Anim. Sci. 1973, 36, 1088–1093. [Google Scholar] [CrossRef]
- Kim, N.-K.; Joh, J.-H.; Park, H.-R.; Kim, O.-H.; Park, B.-Y.; Lee, C.-S. Differential expression profiling of the proteomes and their mRNAs in porcine white and red skeletal muscles. Proteomics 2004, 4, 3422–3428. [Google Scholar] [CrossRef]
- Guo, Y.; Li, J.; Shang, J.; Jin, Y. Development of Muscle-related Genes and Their Effects on Meat Quality. Energy Procedia 2012, 16, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, A.H.; Klont, R.E.; Fernandez, X. Skeletal muscle fibres as factors for pork quality. Livest. Prod. Sci. 1999, 60, 255–269. [Google Scholar] [CrossRef]
- Wimmers, K.; Murani, E.; Ngu, N.T.; Schellander, K.; Ponsuksili, S. Structural and functional genomics to elucidate the genetic background of microstructural and biophysical muscle properties in the pig. J. Anim. Breed. Genet. 2007, 124, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Quiroz-Rothe, E.; Rivero, J.L.L. Coordinated expression of myosin heavy chains, metabolic enzymes, and morphological fea-tures of porcine skeletal muscle fiber types. Microsc. Res. Techniq. 2004, 65, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Serra, X.; Gil, F.; Perez-Enciso, M.; Oliver, M.; Vázquez, J.; Gispert, M.; Díaz, I.; Moreno, F.; Latorre, R.; Noguera, J. A comparison of carcass, meat quality and histochemical characteristics of Iberian (Guadyerbas line) and Landrace pigs. Livest. Prod. Sci. 1998, 56, 215–223. [Google Scholar] [CrossRef]
- Essén-Gustavsson, B.; Fjelkner-Modig, S. Skeletal muscle characteristics in different breeds of pigs in relation to sensory properties of meat. Meat Sci. 1985, 13, 33–47. [Google Scholar] [CrossRef]
- Wang, C.-R.; Chen, Q.-M. Study on muscle histological characteristics of Beijing black pig, Min pig and Large White pig. Chin. J. Anim. Sci. 1996, 4, 33–34. [Google Scholar]
- Kang, P.; Griggs, R.C. Advances in Muscular Dystrophies. JAMA Neurol. 2015, 72, 741–742. [Google Scholar] [CrossRef]
- Geeves, M.A.; Fedorov, R.; Manstein, D.J. Molecular mechanism of actomyosin-based motility. Cell Mol. Life Sci. 2005, 62, 1462–1477. [Google Scholar] [CrossRef]
- Edmondson, D.G.; Lyons, G.E.; Martin, J.F.; Olson, E.N. Mef2 gene expression marks the cardiac and skeletal muscle lineages during mouse embryogenesis. Development 1994, 120, 1251–1263. [Google Scholar] [CrossRef]
- Nabeshima, Y.; Hanaoka, K.; Hayasaka, M.; Esuml, E.; Li, S.; Nonaka, I.l.; Nabeshima, Y.I. Myogenin gene disruption results in perinatal lethality because of severe muscle defect. Nature 1993, 364, 532–535. [Google Scholar] [CrossRef] [PubMed]
- Verardo, L.L.; Nascimento, C.S.; Silva, F.F.; Gasparino, E.; Martins, M.F.; Toriyama, E.; Faria, V.R.; Botelho, M.E.; Costa, K.A.; Lopes, P.S.; et al. Identification and validation of differentially expressed genes from pig skeletal muscle. Anim. Breed. Genet. 2013, 130, 372–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.C.; da Costa, N.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Kang, X.; Wu, L.; Yuan, W.; Dong, Y. The study on the function of MYL2 gene in muscle growth. Heilongjiang Anim. Sci. Vet. Med. 2018, 3, 6. [Google Scholar]
- Zhang, S.-Z.; Xu, Y.; Xie, H.-Q.; Li, X.-Q.; Wei, Y.-Q.; Yang, Z.-M. The possible role of myosin light chain in myoblast proliferation. Biol. Res. 2009, 42, 121–132. [Google Scholar] [CrossRef]
- Fu, M.-Z.; Lin, X.-W. Lean Hogs, Specialized in h-fertilized male duroc pigs growth and development performance test. Sichuan Anim. Vet. Sci. 1996, 4, 16–17. [Google Scholar]
- Li, Q.-H. Guangxi Luchuan pig. Chin. J. Vet. Med. 1963. [Google Scholar]
- Park, I.; Han, C.; Jin, S.; Lee, B.; Choi, H.; Kwon, J.T.; Kim, D.; Kim, J.; Lifirsu, E.; Park, W.J.; et al. Myosin regulatory light chains are required to maintain the stability of myosin II and cellular integrity. Bio-Chem. J. 2011, 434, 171–180. [Google Scholar] [CrossRef]
- Potthoff, M.J.; Wu, H.; Arnold, M.A.; Shelton, J.M.; Backs, J.; McAnally, J.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. Faculty Opinions recommendation of Histone deacetylase degradation and MEF2 activation promote the formation of slow-twitch myofibers. J. Clin. Investig. 2007, 117, 2459–2467. [Google Scholar] [CrossRef] [Green Version]
- Fulco, M.; Schiltz, R.L.; Iezzi, S.; King, M.T.; Zhao, P.; Kashiwaya, Y.; Hoffman, E.; Veech, R.L.; Sartorelli, V. Sir2 Regulates Skeletal Muscle Differentiation as a Potential Sensor of the Redox State. Mol. Cell 2003, 12, 51–62. [Google Scholar] [CrossRef]
- Alves, H.J.; Alvares, L.E.; Gabriel, J.E.; Coutinho, L.L. Influence of the neural tube/notochord complex on MyoD expression and cellular proliferation in chicken embryos. Braz. J. Med. Biol. Res. 2003, 36, 191–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-J. Regulation of muscle mass by myostatin. Annu. Rev. Cell Dev. Biol. 2004, 20, 61–86. [Google Scholar] [CrossRef] [PubMed]
- de Koning, D.J.; Janss, L.L.; Rattink, A.P.; Van Oers, P.A.; de Vries, B.J.; Groenen, M.A.; Van der Poel, J.J.; de Groot, P.N.; Brascamp, E.W.; van Arendonk, J.A. Detection of Quantitative Trait Loci for Backfat Thickness and Intramuscular Fat Content in Pigs (Sus scrofa). Genetics 1999, 152, 1679–1690. [Google Scholar] [CrossRef] [PubMed]
- Hoashi, S.; Hinenoya, T.; Tanaka, A.; Ohsaki, H.; Sasazaki, S.; Taniguchi, M.; Oyama, K.; Mukai, F.; Mannen, H. Association between fatty acid compositions and genotypes of FABP4 and LXR-alpha in Japanese black cattle. BMC Genet. 2008, 9, 84. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.R.; Davie, J.K.; Myer, A.; Meadows, E.; Olson, E.N.; Klein, W.H. Loss of myogenin in postnatal life leads to normal skeletal muscle but reduced body size. Development 2006, 133, 601–610. [Google Scholar] [CrossRef] [Green Version]
- Koohmaraie, M. Biochemical factors regulating the toughening and tenderization processes of meat. Meat Sci. 1996, 43, 193–201. [Google Scholar] [CrossRef]
- Vandenburgh, H.H.; Karlisch, P.; Shansky, J.; Feldstein, R. Insulin and IGF-I induce pronounced hypertrophy of skeletal myo-fibers in tissue culture. Am. J. Physiol. 1991, 260, C475–C484. [Google Scholar] [CrossRef]
- Barton-Davis, E.R.; I Shoturma, D.; Sweeney, H.L. Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiol. Scand. 1999, 167, 301–305. [Google Scholar] [CrossRef]
- Elia, D.; Madhala, D.; Ardon, E.; Reshef, R.; Halevy, O. Sonic hedgehog promotes proliferation and differentiation of adult muscle cells: Involvement of MAPK/ERK and PI3K/Akt pathways. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2007, 1773, 1438–1446. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID in NCBI | Primer Type | Primer Sequence (5′→3′) | Product Size (bp) |
|---|---|---|---|---|
| MYL2-sus | NM_213791.2 | RT-qPCR | Forward: ACAGGGATGGCTTCATAGACAA | 156 |
| Reward: TAAGTTTCTCCCCAAACATCGT | ||||
| IGF1-sus | NM_214256.1 | RT-qPCR | Forward: GGACGCTCTTCAGTTCGTGT | 117 |
| Reward: CGGAAGCAGCACTCATCCAC | ||||
| IGF2-sus | NM_213883.2 | RT-qPCR | Forward: ACACCCTCCAGTTTGTCT | 236 |
| Reward: GTCATAGCGGAAGAACTTGCC | ||||
| MYH1-sus | NM_001104951.2 | RT-qPCR | Forward: GTGCCGTACTCTAGAAGATCAAC | 158 |
| Reward: GCTGAGAAACTAACGTGTCCT | ||||
| MYH4-sus | NM_001123141.1 | RT-qPCR | Forward: TACTTAGGAAGAAGCACGCAGAC | 242 |
| Reward: ATTGATTAGGCGCTGATGCTC | ||||
| PDK3-sus | XM_021080538.1 | RT-qPCR | Forward: CACTTCTCTTTGGCGGTGACA | 120 |
| Reward: CACAAAGCATCTTGGCGGTTT | ||||
| MYLPF-sus | NM_001006592.1 | RT-qPCR | Forward: CAGAAGGAAGCTCCAACGTCT Reward: GAGCTTCTCCCCAAACATGGTC | 239 |
| TXNIP-sus | NM_001044614.2 | RT-qPCR | Forward: CTAGTGGATGTCAATACCCCT Reward: TTGCCTCTGACCGATGACAAC | 278 |
| GAPDH-sus | NM_001206359.1 | RT-qPCR | Forward: AACATCATCCCTGCTTCTACCG | 158 |
| Reward: CAGGTCAGATCCACAACCGACA | ||||
| MYH1-mus | NM_030679.2 | RT-qPCR | Forward: GCAGCTCCAAGTTCAGTCT | 289 |
| Reward: TGCCATCTCCTCTGTCAGGT | ||||
| MYH2-mus | NM_001039545.2 | RT-qPCR | Forward: AAGCAGAGGCAAGTAGTGGT | 124 |
| Reward: GTGCTCCTGAGATTGGTCAT | ||||
| MYH4-mus | NM_010855.3 | RT-qPCR | Forward: GTTCTTCTTTCCAGACCGT | 130 |
| Reward: ATGGCACCAGGAGTCTTAG | ||||
| MYH7-mus | NM_080728.3 | RT-qPCR | Forward: GACAAAGGCAAAGGCAAGG | 129 |
| Reward: GATGATGCAGCGTACAAAG | ||||
| MYOD-mus | NM_010866.2 | RT-qPCR | Forward: TGGCATGATGGATTACAGC Reward: TTCCCTGTTCTGTGTCGCTT | 260 |
| MEF2c-mus | NM_001347567.1 | RT-qPCR | Forward: CTCTGTCTGGCTTCAACACT Reward: CTGACTTGATGCTGAGGCTTT | 183 |
| MSTN-mus | NM_010834.3 | RT-qPCR | Forward: ATGACGATTATCACGCTACCAC Reward: ACCTTGTACCGTCTTTCAT | 234 |
| MYL2-clone | NM_213791.2 | PCR | Reward: CCATGTCACCTAAGAAAGCCAAG | 549 |
| Reward: CCACTCACCCAAAGGCAAG | ||||
| MYL2-vc a | NM_213791.2 | PCR | Forward: CCCAAGCTTATGTCACCTAAGAAAGCCAAGAAG | 503 |
| Reward: CGCGGATCCCCTTAGTCCTTCTCTTCTCCGTGG |
| Sample | D1F | D1M | D2F | D2M | L1F | L1M | L2F | L2M |
|---|---|---|---|---|---|---|---|---|
| Total Raw Reads (M) | 75.32 | 73.68 | 77.02 | 77.01 | 77.75 | 80.23 | 75.32 | 73.69 |
| Total Clean Reads (M) | 72.65 | 71.8 | 66.57 | 66.44 | 64.23 | 65.15 | 72.85 | 71.35 |
| Total Clean Bases (Gb) | 7.27 | 7.18 | 6.66 | 6.64 | 6.42 | 6.52 | 7.29 | 7.14 |
| Clean Reads Q20 (%) | 98.75 | 98.87 | 97.97 | 98.07 | 98.04 | 97.83 | 98.66 | 98.64 |
| Clean Reads Q30 (%) | 96.05 | 96.34 | 91.89 | 92.11 | 91.77 | 91.13 | 95.83 | 95.78 |
| Clean Reads Ratio (%) | 96.45 | 97.44 | 86.44 | 86.27 | 82.61 | 81.12 | 96.72 | 96.84 |
| Total Mapping Ratio | 91.79% | 92.36% | 95.39% | 95.22% | 93.05% | 93.56% | 91.10% | 90.56% |
| Uniquely Mapping Ratio | 68.94% | 70.28% | 67.05% | 68.74% | 63.74% | 58.74% | 67.56% | 67.57% |
| Pathway | DEGs Genes with Pathway Annotation (2884) | p Value | Pathway ID | |
|---|---|---|---|---|
| 1 | Proteasome | 32 | 2.74 × 10−12 | ko03050 |
| 2 | Amoebiasis | 80 | 1.09 × 10−5 | ko05146 |
| 3 | Complement and coagulation cascades | 41 | 1.68 × 10−5 | ko04610 |
| 4 | Glycine, serine and threonine metabolism | 19 | 5.83 × 10−5 | ko00260 |
| 5 | Arachidonic acid metabolism | 34 | 0.000107 | ko00590 |
| 6 | ECM-receptor interaction | 60 | 0.000279 | ko04512 |
| 7 | Legionellosis | 30 | 0.000316 | ko05134 |
| 8 | Epstein-Barr virus infection | 72 | 0.000551 | ko05169 |
| 9 | Human papillomavirus infection | 121 | 0.000915 | ko05165 |
| 10 | Staphylococcus aureus infection | 27 | 0.001579 | ko05150 |
| 11 | p53 signaling pathway | 31 | 0.00158 | ko04115 |
| 12 | PI3K-Akt signaling pathway | 117 | 0.001723 | ko04151 |
| 13 | Phagosome | 57 | 0.002097 | ko04145 |
| 14 | Fatty acid metabolism | 19 | 0.003029 | ko01212 |
| 15 | Toxoplasmosis | 37 | 0.003322 | ko05145 |
| 16 | Mitophagy-animal | 21 | 0.003851 | ko04137 |
| 17 | Small cell lung cancer | 40 | 0.004097 | ko05222 |
| 18 | Cellular senescence | 50 | 0.004594 | ko04218 |
| 19 | Focal adhesion | 87 | 0.004764 | ko04510 |
| 20 | FoxO signaling pathway | 37 | 0.00515 | ko04068 |
| 21 | MAPK signaling pathway | 79 | 0.005275 | ko04010 |
| 22 | Hematopoietic cell lineage | 35 | 0.00558 | ko04640 |
| 23 | DNA replication | 19 | 0.006139 | ko03030 |
| 24 | PPAR signaling pathway | 28 | 0.00717 | ko03320 |
| 25 | Transcriptional misregulation in cancer | 64 | 0.007257 | ko05202 |
| 26 | Fc epsilon RI signaling pathway | 26 | 0.008615 | ko04664 |
| 27 | Platinum drug resistance | 25 | 0.008867 | ko01524 |
| 28 | Pathways in cancer | 150 | 0.009306 | ko05200 |
| 29 | Amyotrophic lateral sclerosis (ALS) | 23 | 0.009324 | ko05014 |
| 30 | Gastric cancer | 56 | 0.009609 | ko05226 |
| GeneID | log2Ratio (L/D) | q-Value | Up-Down-Regulation (L/D) | p-Value | Symbol |
|---|---|---|---|---|---|
| 100144306 | −2.585066 | 0 | Down | 0 | MYH4 |
| 100125538 | −1.110562 | 0 | Down | 0 | MYH1 |
| 474162 | −1.163615 | 0 | Down | 0 | MYLPF |
| 396725 | 1.20822 | 0 | Up | 0 | DES |
| 396718 | −1.080459 | 0 | Down | 0 | RYR1 |
| 100286778 | 7.811893 | 0 | Up | 0 | PDK4 |
| 397667 | 1.096899 | 0 | Up | 0 | FHL1 |
| 733657 | 5.046722 | 0 | Up | 0 | FBXO32 |
| 396916 | −2.672299 | 0 | Down | 0 | IGF2 |
| 733688 | 2.369902 | 0 | Up | 0 | TXNIP |
| 100152001 | −1.666755 | 0 | Down | 0 | COL3A1 |
| 414388 | 1.804317 | 0 | Up | 0 | TPM3 |
| 100517321 | 3.11986 | 0 | Up | 0 | TSC22D3 |
| 396690 | 1.081174 | 0 | Up | 0 | MYL2 |
| 100525195 | 2.038505 | 0 | Up | 0 | LMOD2 |
| 100271745 | 1.007136 | 0 | Up | 0 | TCAP |
| 396947 | 1.090353 | 0 | Up | 0 | TNNI1 |
| 397583 | 4.714497 | 0 | Up | 0 | LIPE |
| 397077 | 2.081716 | 0 | Up | 0 | FOXO1 |
| 780431 | 1.834607 | 0 | Up | 0 | NDRG2 |
| 396959 | 1.35284 | 0 | Up | 0 | ANKRD1 |
| 100156435 | 1.058298 | 0 | Up | 0 | TNNC1 |
| 100048933 | 1.669136 | 0 | Up | 0 | ZFAND5 |
| 397116 | 3.909268 | 0 | Up | 0 | UCP3 |
| 100294702 | −1.503496 | 0 | Down | 0 | SMYD1 |
| 100157318 | 3.569646 | 0 | Up | 0 | APOD |
| 100624868 | 1.763672 | 0 | Up | 0 | BNIP3 |
| 100519789 | 1.258786 | 0 | Up | 0 | CRYAB |
| 100152767 | −1.115181 | 0 | Down | 0 | KLHL31 |
| 100627008 | −1.382584 | 0 | Down | 0 | DMPK |
| 100520636 | 1.633977 | 0 | Up | 0 | ACADVL |
| 733702 | 1.161209 | 0 | Up | 0 | CD36 |
| 100134962 | 1.63106 | 0 | Up | 0 | PHYH |
| 100337687 | 1.69399 | 0 | Up | 0 | CSRP3 |
| 100240723 | 1.662207 | 0 | Up | 0 | SGCA |
| 100620966 | 4.171741 | 0 | Up | 0 | SESN2 |
| 100157793 | −1.276461 | 0 | Down | 0 | PRKAB2 |
| 100152267 | −2.014984 | 0 | Down | 0 | ACTC1 |
| 100152836 | 1.60654 | 0 | Up | 0 | GPAT4 |
| 397012 | 1.111651 | 0 | Up | 0 | HADHA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, P.; Qin, Z.; Xie, W.; Chen, B.; Guan, Z.; Xie, B. Identification of Differentially Expressed Genes in the Longissimus Dorsi Muscle of Luchuan and Duroc Pigs by Transcriptome Sequencing. Genes 2023, 14, 132. https://doi.org/10.3390/genes14010132
Pan P, Qin Z, Xie W, Chen B, Guan Z, Xie B. Identification of Differentially Expressed Genes in the Longissimus Dorsi Muscle of Luchuan and Duroc Pigs by Transcriptome Sequencing. Genes. 2023; 14(1):132. https://doi.org/10.3390/genes14010132
Chicago/Turabian StylePan, Pengcheng, Zhaoxian Qin, Wan Xie, Baojian Chen, Zhihui Guan, and Bingkun Xie. 2023. "Identification of Differentially Expressed Genes in the Longissimus Dorsi Muscle of Luchuan and Duroc Pigs by Transcriptome Sequencing" Genes 14, no. 1: 132. https://doi.org/10.3390/genes14010132