
G-quadruplexes Mark Sites of Methylation Instability Associated with Ageing and Cancer


Abstract
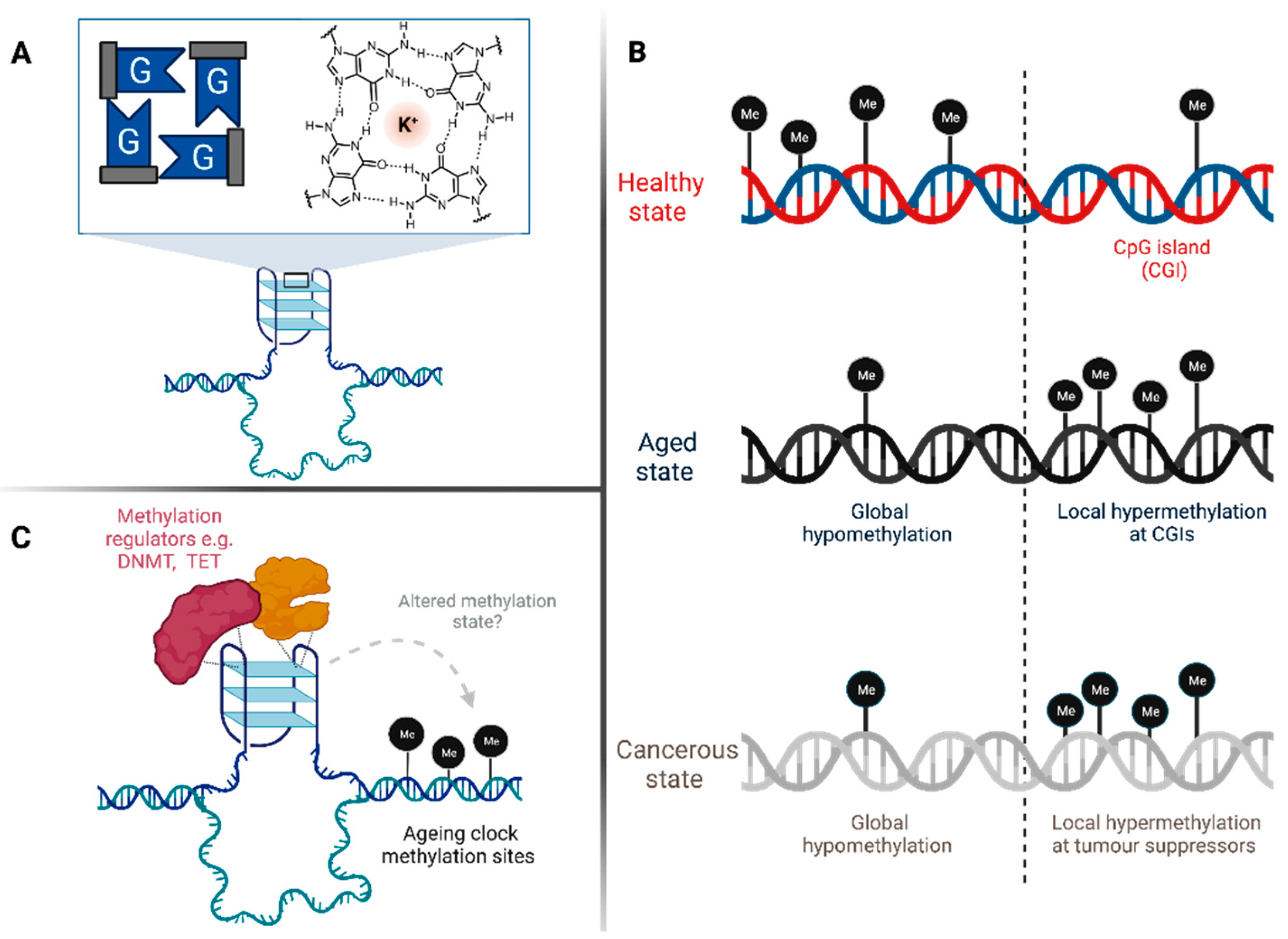
:1. Introduction
2. Materials and Methods
2.1. Enrichment and Statistical Tests
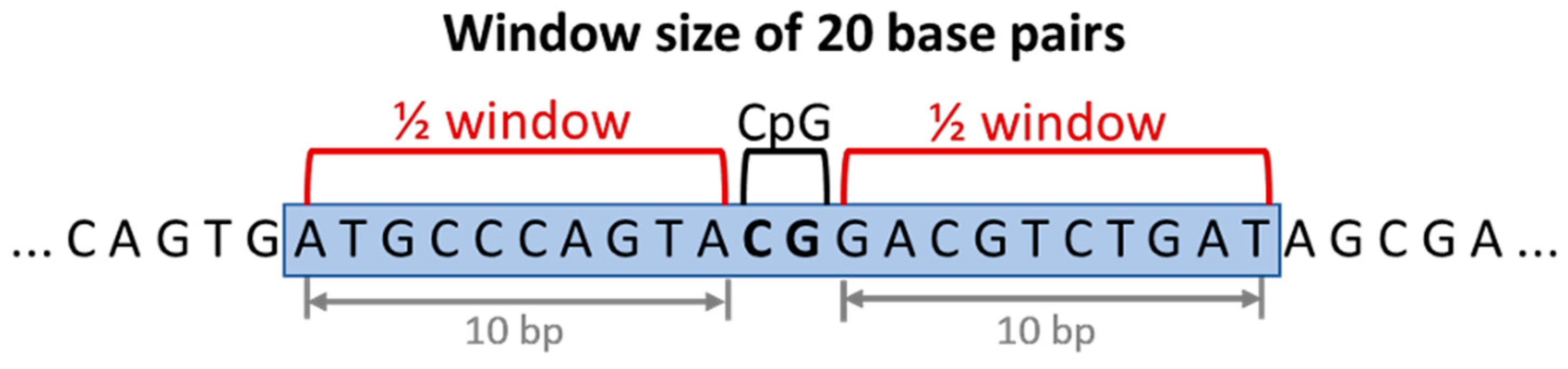
2.1.1. Window Size Analysis
2.1.2. G4 Enrichment Relative to Random
2.1.3. G4 Enrichment Relative to Global CpG Enrichment
- All instances of the CG motif were extracted from the reference genomes using the function matchPattern(‘CG’, chromosome) from the Biostrings package;
- The coordinates of CpGs that overlap with G4s were found using intersect from the GenomicRanges package;
- The fraction (f) of CpGs that intersect with G4s was found by applying subsetByOverlaps to the intersected file;
- The expected number (E[n]) of ageing clock CpGs intersecting with G4s was found by multiplying f by the total number of ageing clock CpGs in the genome;
- The actual number (n) of G4s intersecting with ageing clock CpGs was calculated as before using intersect and subsetByOverlap applied to the respective ageing clock files;
- The fold-enrichment of G4s at ageing clock CpGs relative to the expected from all CpGs was then found as n/E[n].
2.1.4. G4 Enrichment at Protein-Binding Sites
3. Results
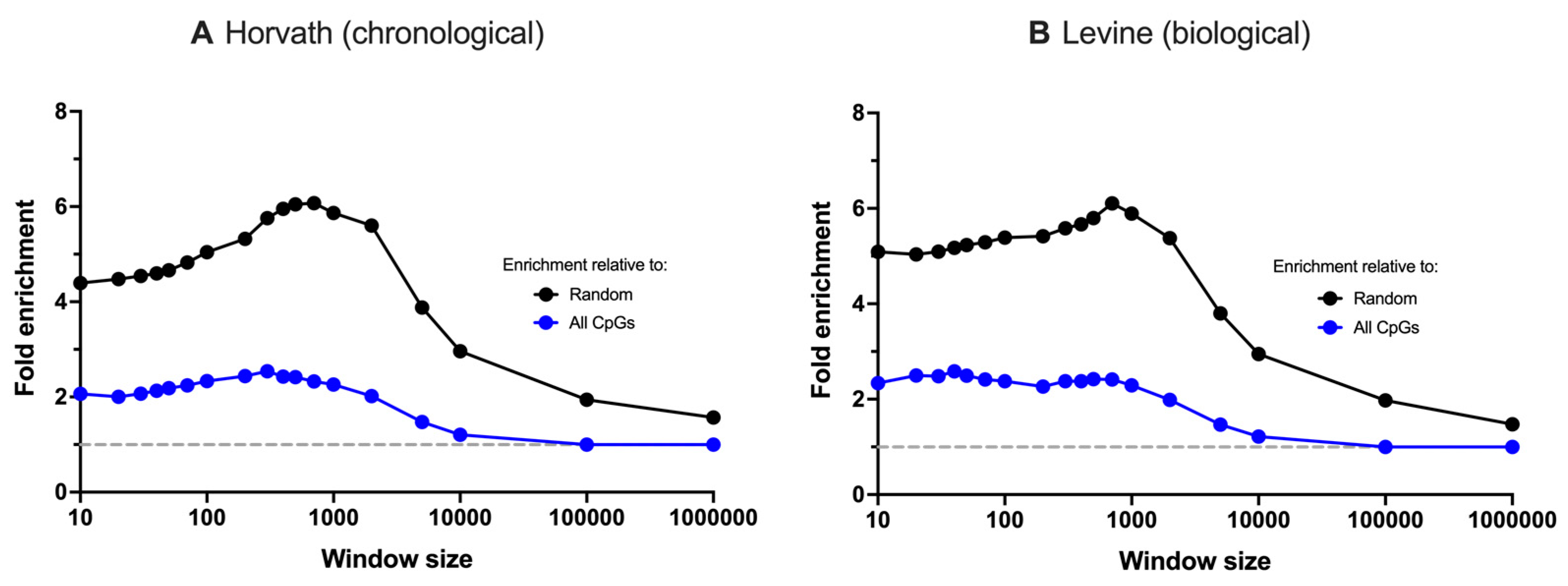
3.1. G4 Sequences Co-Localise with Chronological and Biological Ageing Clocks in Humans
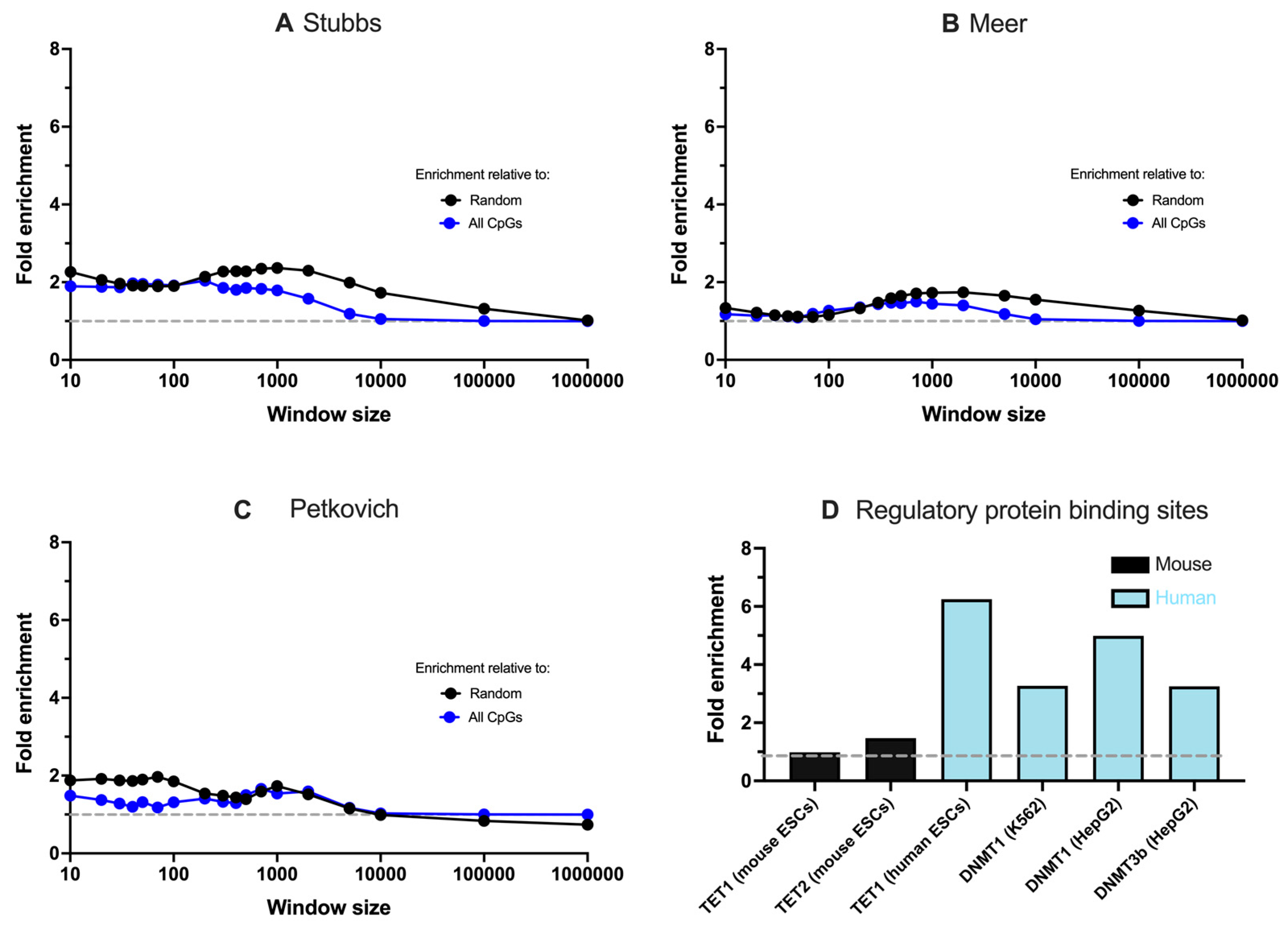
3.2. Associations between G4s, Ageing Clocks, and Methylation Regulators Are Specific to Humans
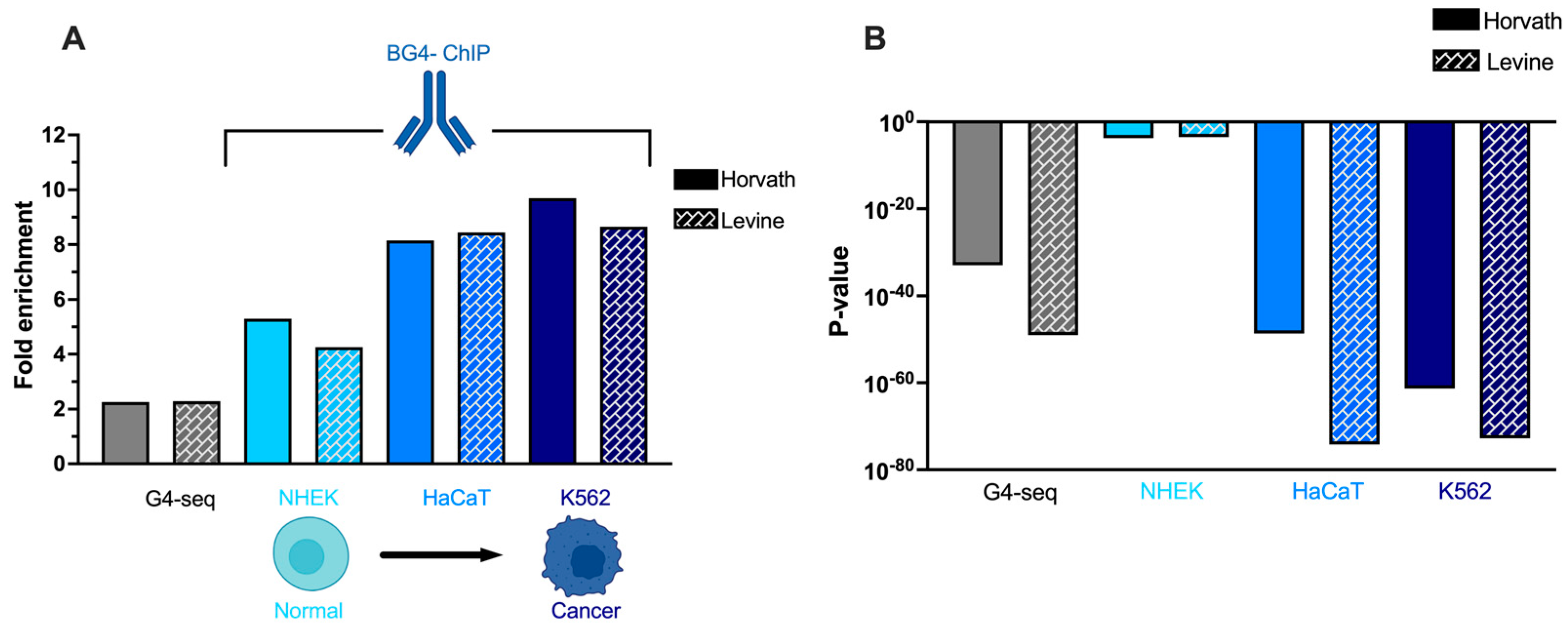
3.3. G4 Enrichment at Ageing Clocks Is Greatest in Immortalised and Tumorigenic Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mukherjee, A.K.; Sharma, S.; Chowdhury, S. Non-duplex G-Quadruplex Structures Emerge as Mediators of Epigenetic Modifications. Trends Genet. 2019, 35, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Varizhuk, A.; Isaakova, E.; Pozmogova, G. DNA G-Quadruplexes (G4s) Modulate Epigenetic (Re)Programming and Chromatin Remodeling. BioEssays 2019, 41, e1900091. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Raguseo, F.; Nuccio, S.P.; Liano, D.; Di Antonio, M. DNA G-quadruplex structures: More than simple roadblocks to transcription? Nucleic Acids Res. 2021, 49, 8419–8431. [Google Scholar] [CrossRef]
- Spiegel, J.; Adhikari, S.; Balasubramanian, S. The Structure and Function of DNA G-Quadruplexes. Trends Chem. 2020, 2, 123–136. [Google Scholar] [CrossRef]
- Raguseo, F.; Chowdhury, S.; Minard, A.; Di Antonio, M. Chemical-biology approaches to probe DNA and RNA G-quadruplex structures in the genome. Chem. Commun. 2020, 56, 1317–1324. [Google Scholar] [CrossRef]
- Puig Lombardi, E.; Londoño-Vallejo, A. A guide to computational methods for G-quadruplex prediction. Nucleic Acids Res. 2020, 48, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Marsico, G.; Chambers, V.S.; Sahakyan, A.B.; McCauley, P.; Boutell, J.M.; Di Antonio, M.; Balasubramanian, S. Whole genome experimental maps of DNA G-quadruplexes in multiple species. Nucleic Acids Res. 2019, 47, 3862–3874. [Google Scholar] [CrossRef]
- Haensel-Hertsch, R.; Beraldi, D.; Lensing, S.V.; Marsico, G.; Zyner, K.; Parry, A.; Di Antonio, M.; Pike, J.; Kimura, H.; Narita, M.; et al. G-quadruplex structures mark human regulatory chromatin. Nat. Genet. 2016, 48, 1267–1272. [Google Scholar] [CrossRef]
- Haensel-Hertsch, R.; Spiegel, J.; Marsico, G.; Tannahill, D.; Balasubramanian, S. Genome-wide mapping of endogenous G-quadruplex DNA structures by chromatin immunoprecipitation and high-throughput sequencing. Nat. Protoc. 2018, 13, 551–564. [Google Scholar] [CrossRef]
- Kosiol, N.; Juranek, S.; Brossart, P.; Heine, A.; Paeschke, K. G-quadruplexes: A promising target for cancer therapy. Mol. Cancer 2021, 20, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Liano, D.; Chowdhury, S.; Di Antonio, M. Cockayne Syndrome B Protein Selectively Resolves and Interact with Intermolecular DNA G-Quadruplex Structures. J. Am. Chem. Soc. 2021, 143, 20988–21002. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.E.; Cao, K.; Ryvkin, P.; Wang, L.-S.; Johnson, F.B. Altered gene expression in the Werner and Bloom syndromes is associated with sequences having G-quadruplex forming potential. Nucleic Acids Res. 2010, 38, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.-Q.; Ghanbarian, A.T.; Spiegel, J.; Cuesta, S.M.; Beraldi, D.; Di Antonio, M.; Marsico, G.; Haensel-Hertsch, R.; Tannahill, D.; Balasubramanian, S. DNA G-quadruplex structures mold the DNA methylome. Nat. Struct. Mol. Biol. 2018, 25, 951–957. [Google Scholar] [CrossRef]
- Halder, R.; Halder, K.; Sharma, P.; Garg, G.; Sengupta, S.; Chowdhury, S. Guanine quadruplex DNA structure restricts methylation of CpG dinucleotides genome-wide. Mol. BioSyst. 2010, 6, 2439–2447. [Google Scholar] [CrossRef]
- Cree, S.L.; Fredericks, R.; Miller, A.; Pearce, F.G.; Filichev, V.; Fee, C.; Kennedy, M.A. DNA G-quadruplexes show strong interaction with DNA methyltransferases in vitro. FEBS Lett. 2016, 590, 2870–2883. [Google Scholar] [CrossRef]
- Bell, C.G.; Lowe, R.; Adams, P.D.; Baccarelli, A.A.; Beck, S.; Bell, J.T.; Christensen, B.C.; Gladyshev, V.N.; Heijmans, B.T.; Horvath, S.; et al. DNA methylation aging clocks: Challenges and recommendations. Genome Biol. 2019, 20, 249. [Google Scholar] [CrossRef]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, 3156. [Google Scholar] [CrossRef]
- Johnson, A.A.; Akman, K.; Calimport, S.R.G.; Wuttke, D.; Stolzing, A.; de Magalhães, J.P. The Role of DNA Methylation in Aging, Rejuvenation, and Age-Related Disease. Rejuv. Res. 2012, 15, 483. [Google Scholar] [CrossRef]
- Malousi, A.; Andreou, A.-Z.; Georgiou, E.; Tzimagiorgis, G.; Kovatsi, L.; Kouidou, S. Age-dependent methylation in epigenetic clock CpGs is associated with G-quadruplex, co-transcriptionally formed RNA structures and tentative splice sites. Epigenetics 2018, 13, 808–821. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, T.M.; Bonder, M.J.; Stark, A.K.; Krueger, F.; von Meyenn, F.; Stegle, O.; Reik, W.; Bolland, D.; Butcher, G.; Chandra, T.; et al. Multi-Tissue DNA Methylation Age Predictor in Mouse. Genome Biol. 2017, 18, 68. [Google Scholar] [CrossRef] [PubMed]
- Meer, M.V.; Podolskiy, D.I.; Tyshkovskiy, A.; Gladyshev, V.N. A Whole Lifespan Mouse Multi-Tissue DNA Methylation Clock. eLife 2018, 7, e40675. [Google Scholar] [CrossRef] [PubMed]
- Petkovich, D.A.; Podolskiy, D.I.; Lobanov, A.V.; Lee, S.-G.; Miller, R.A.; Gladyshev, V.N. Using DNA Methylation Profiling to Evaluate Biological Age and Longevity Interventions. Cell Metab. 2017, 25, 954–960.e6. [Google Scholar] [CrossRef]
- Williams, K.; Christensen, J.; Pedersen, M.T.; Johansen, J.V.; Cloos, P.A.C.; Rappsilber, J.; Helin, K. TET1 and Hydroxymethylcytosine in Transcription and DNA Methylation Fidelity. Nature 2011, 473, 343–348. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, Y.; Bian, C.; Fujiki, R.; Yu, X. TET2 promotes histone O-GlcNAcylation during gene transcription. Nature 2013, 493, 561–564. [Google Scholar] [CrossRef]
- Verma, N.; Pan, H.; Doré, L.C.; Shukla, A.; Li, Q.V.; Pelham-Webb, B.; Teijeiro, V.; González, F.; Krivtsov, A.; Chang, C.-J.; et al. TET proteins safeguard bivalent promoters from de novo methylation in human embryonic stem cells. Nat. Genet. 2017, 50, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Dunham, I.; Kundaje, A.; Aldred, S.F.; Collins, P.J.; Davis, C.A.; Doyle, F.; Epstein, C.B.; Frietze, S.; Harrow, J.; Kaul, R.; et al. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Scheibye-Knudsen, M.; Longo, D.L.; de Cabo, R. Animal Models of Aging Research: Implications for Human Aging and Age-Related Diseases. Annu. Rev. Anim. Biosci. 2015, 3, 283–303. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Description | Data Location | Reference |
|---|---|---|
| Horvath ageing clock | Publication suppl. data | Horvath (2013) [19] |
| Levine ageing clock | Publication suppl. Data | Levine et al. (2018) [24] |
| Stubbs ageing clock | GEO: GSE93957/publication suppl. Data | Stubbs et al. (2017) [25] |
| Meer ageing clock | GEO: GSE121141/publication suppl. Data) | Meer et al. (2018) [26] |
| Petkovich ageing clock | GEO: GSE80672/publication suppl. Data) | Petkovich et al. (2017) [27] |
| Human G4-seq (K+) | GEO: GSM3003539 | Marsico et al. (2019) [8] |
| Mice G4-seq (K+) | GEO: GSM3003547 | Marsico et al. (2019) [8] |
| TET1 (mouse ESCs) ChIP-Seq | GEO: GSE24841 | Williams et al. (2011) [25] |
| TET2 (mouse ESCs) ChIP-seq | GEO: GSE41720 | Chen et al. (2013) [26] |
| TET1 (human ESCs) ChIP-seq | GEO: GSE99346 | Verma et al. (2018) [27] |
| DNMT1 (K562) ChIP-seq | GEO: GSE92213 | ENCODE [28] |
| DNMT1 (HepG2) ChIP-seq | GEO: GSE170872 | ENCODE [28] |
| DNMT3b (HepG2) ChIP-seq | GEO: GSE95953 | ENCODE [28] |
| NHEK BG4-ChIP | GEO: GSE76688 | Hänsel-Hertsch et al. (2016) [9] |
| HaCaT BG4-ChIP | GEO: GSE76688 | Hänsel-Hertsch et al. (2016) [9] |
| K562 BG4-ChIP | GEO: GSE107690 | Mao et al. (2018) [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rauchhaus, J.; Robinson, J.; Monti, L.; Di Antonio, M. G-quadruplexes Mark Sites of Methylation Instability Associated with Ageing and Cancer. Genes 2022, 13, 1665. https://doi.org/10.3390/genes13091665
Rauchhaus J, Robinson J, Monti L, Di Antonio M. G-quadruplexes Mark Sites of Methylation Instability Associated with Ageing and Cancer. Genes. 2022; 13(9):1665. https://doi.org/10.3390/genes13091665
Chicago/Turabian StyleRauchhaus, Jonas, Jenna Robinson, Ludovica Monti, and Marco Di Antonio. 2022. "G-quadruplexes Mark Sites of Methylation Instability Associated with Ageing and Cancer" Genes 13, no. 9: 1665. https://doi.org/10.3390/genes13091665