
Expression Profile of New Gene Markers Involved in Differentiation of Canine Adipose-Derived Stem Cells into Chondrocytes
 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Material Collection
2.2. Cell Sample Preparation
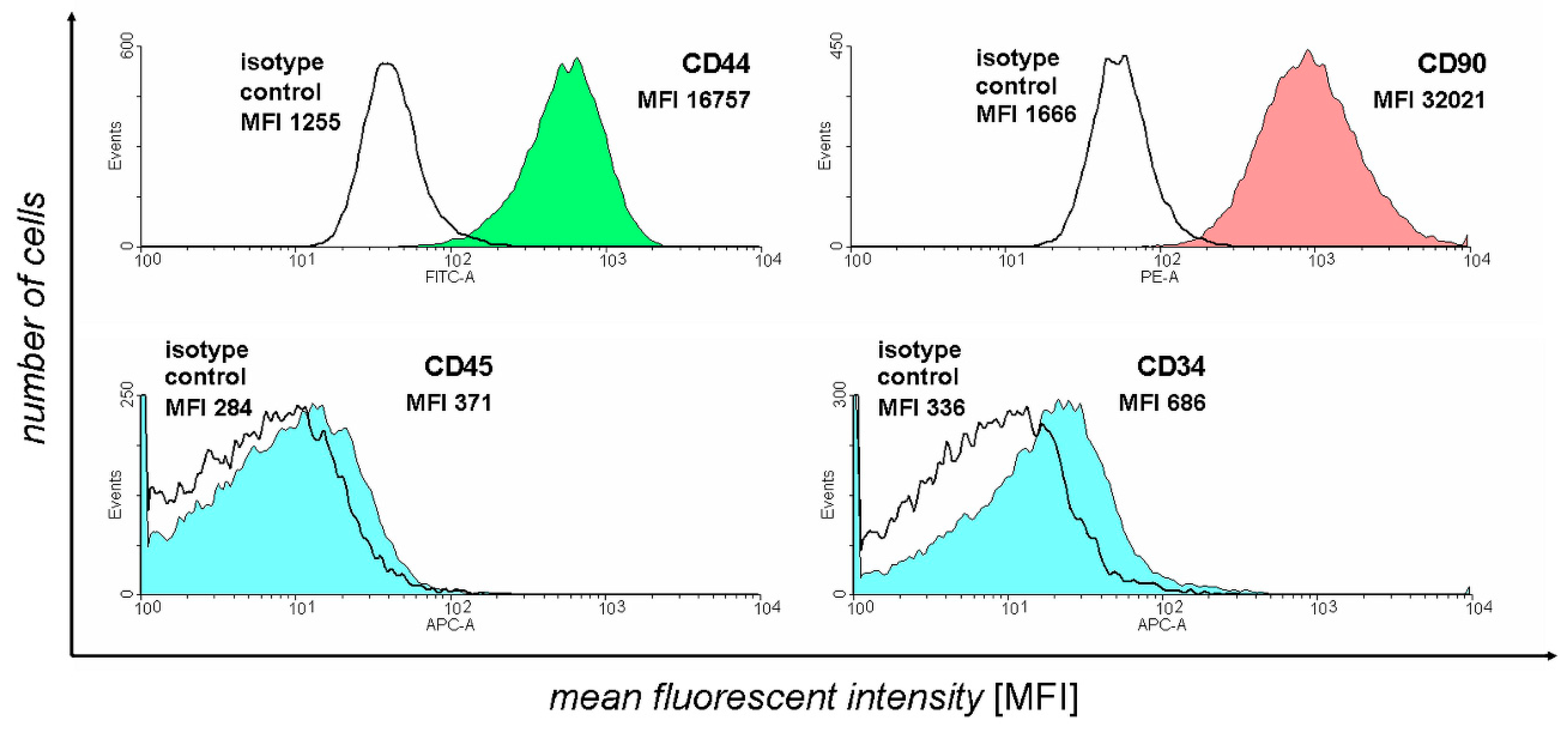
2.3. Flow Cytometry Analysis
2.4. In Vitro Cell Culture
2.5. Chondrogenic Differentiation
2.6. RNA Isolation
2.7. RNAseq Analysis
2.8. RT-qPCR Validation
2.9. Bioinformatical and Statistical Analysis
3. Results
3.1. Flow Cytometry Analysis
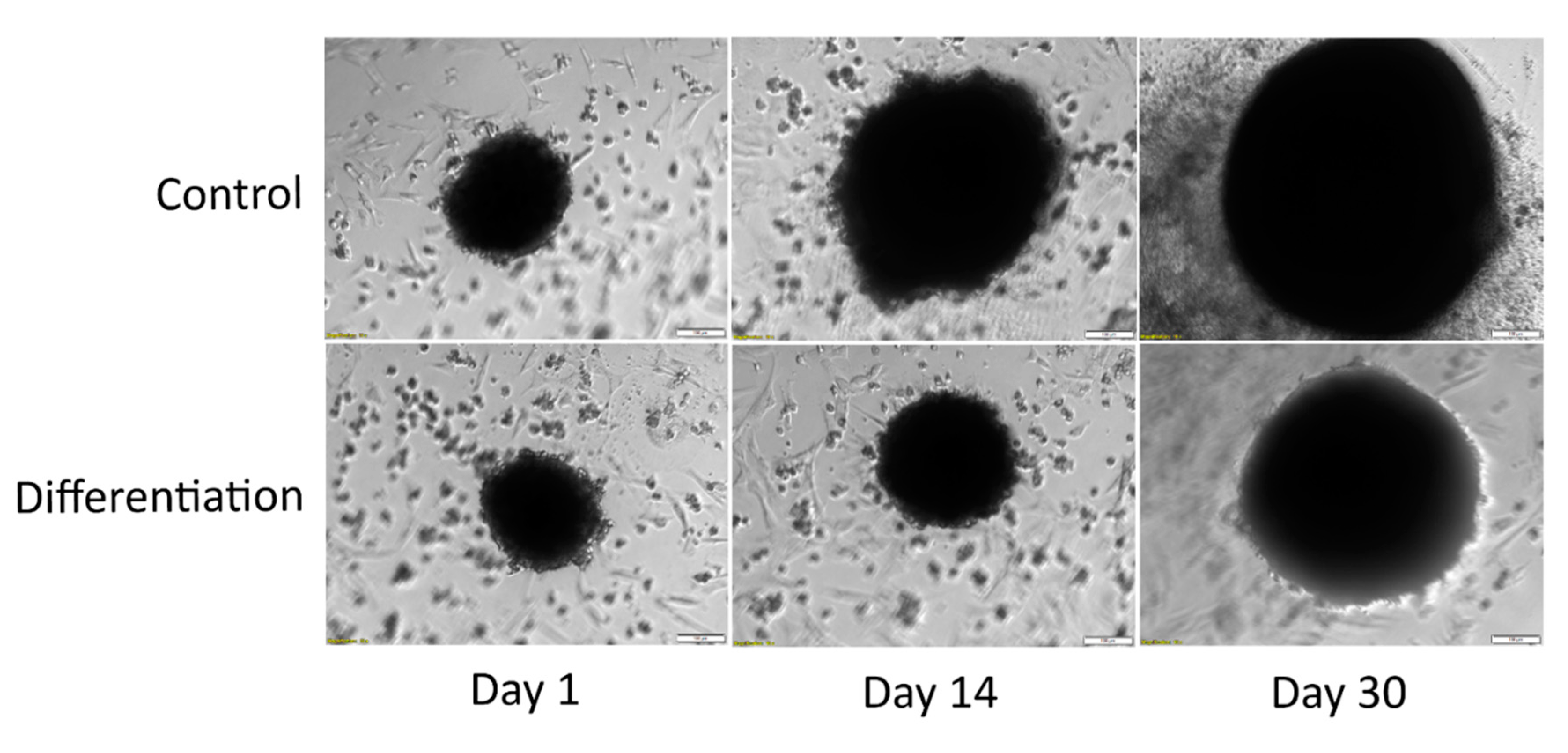
3.2. Morphological Analysis
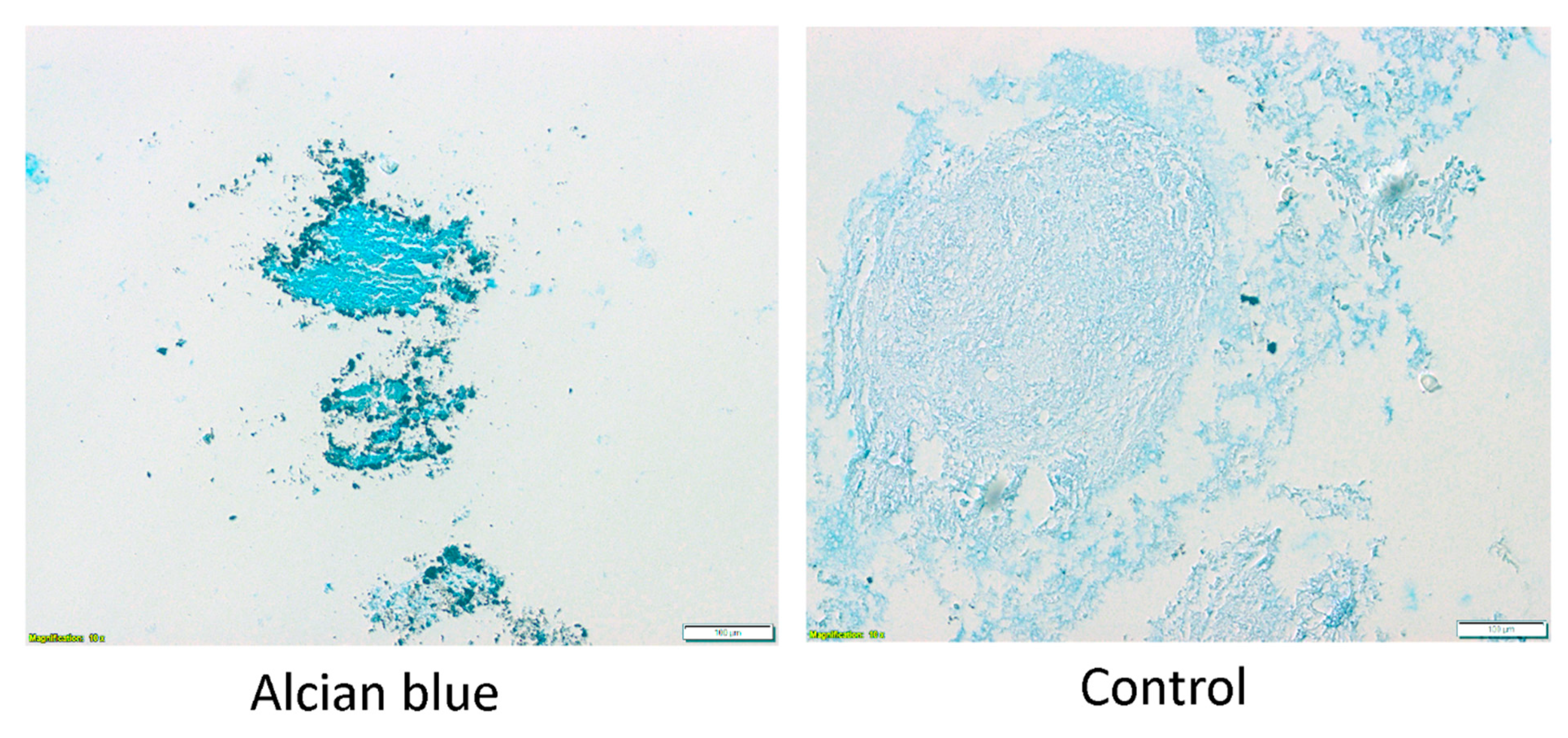
3.3. Evaluation of Chondrogenic Differentiation
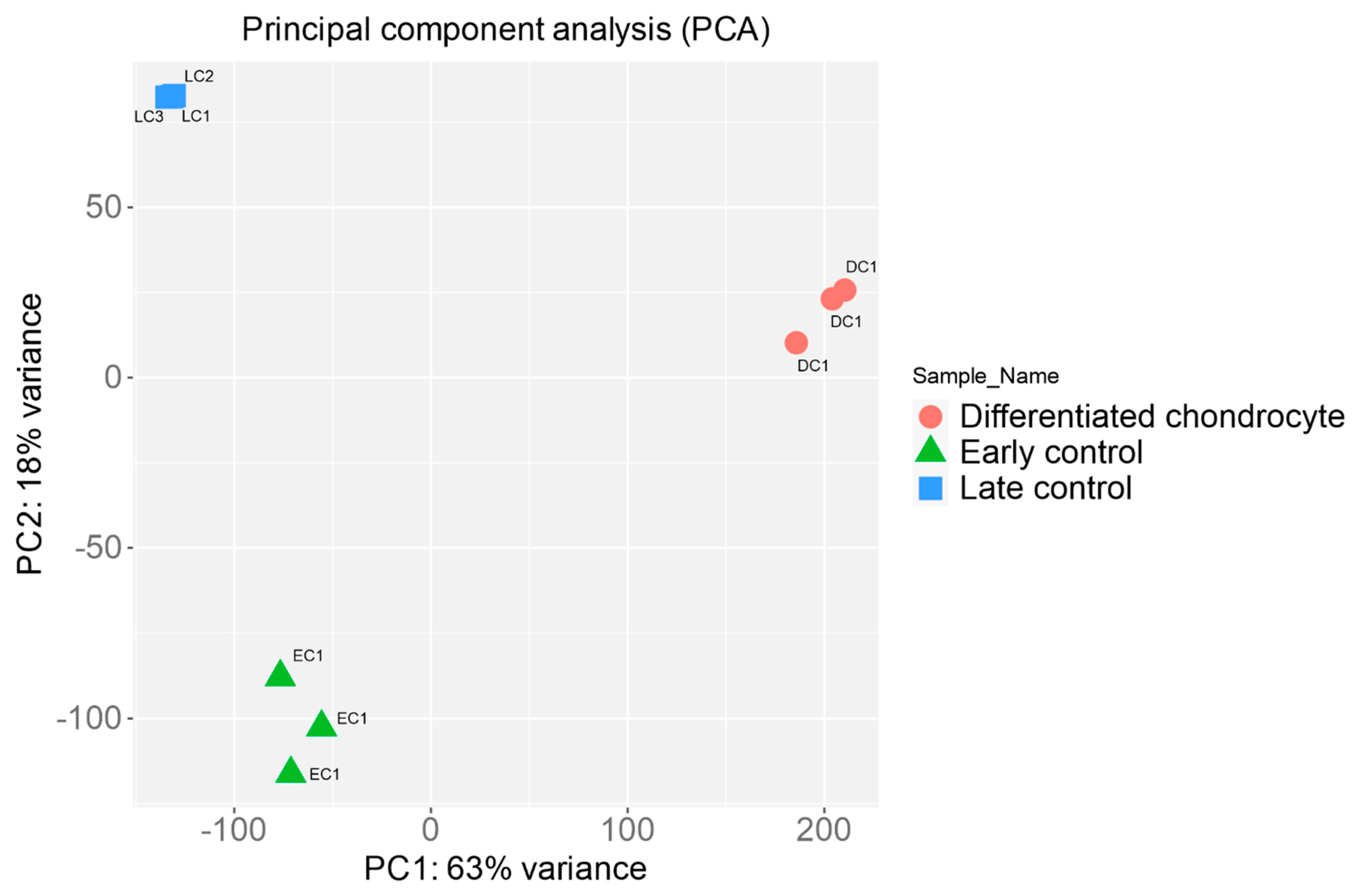
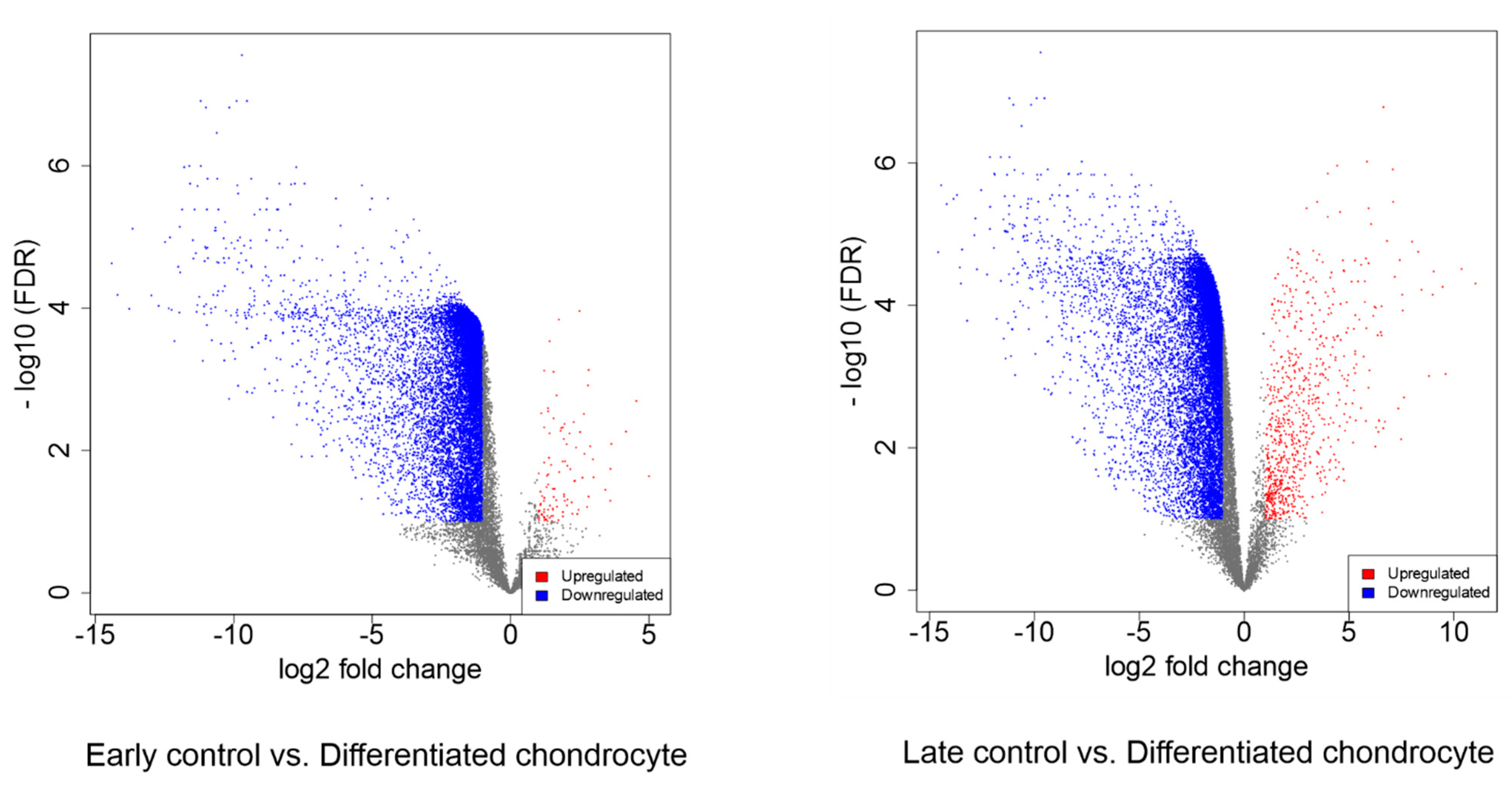
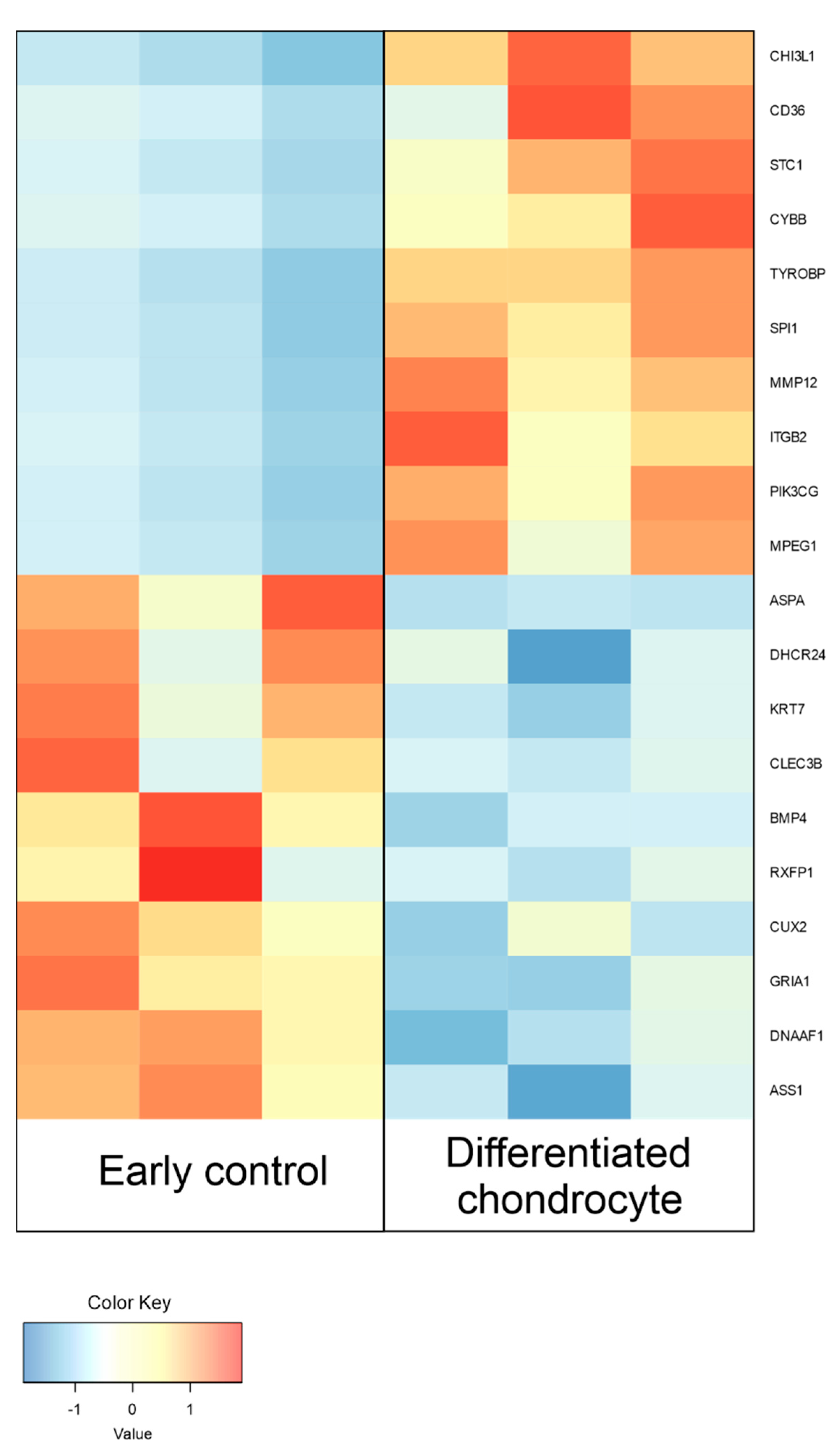
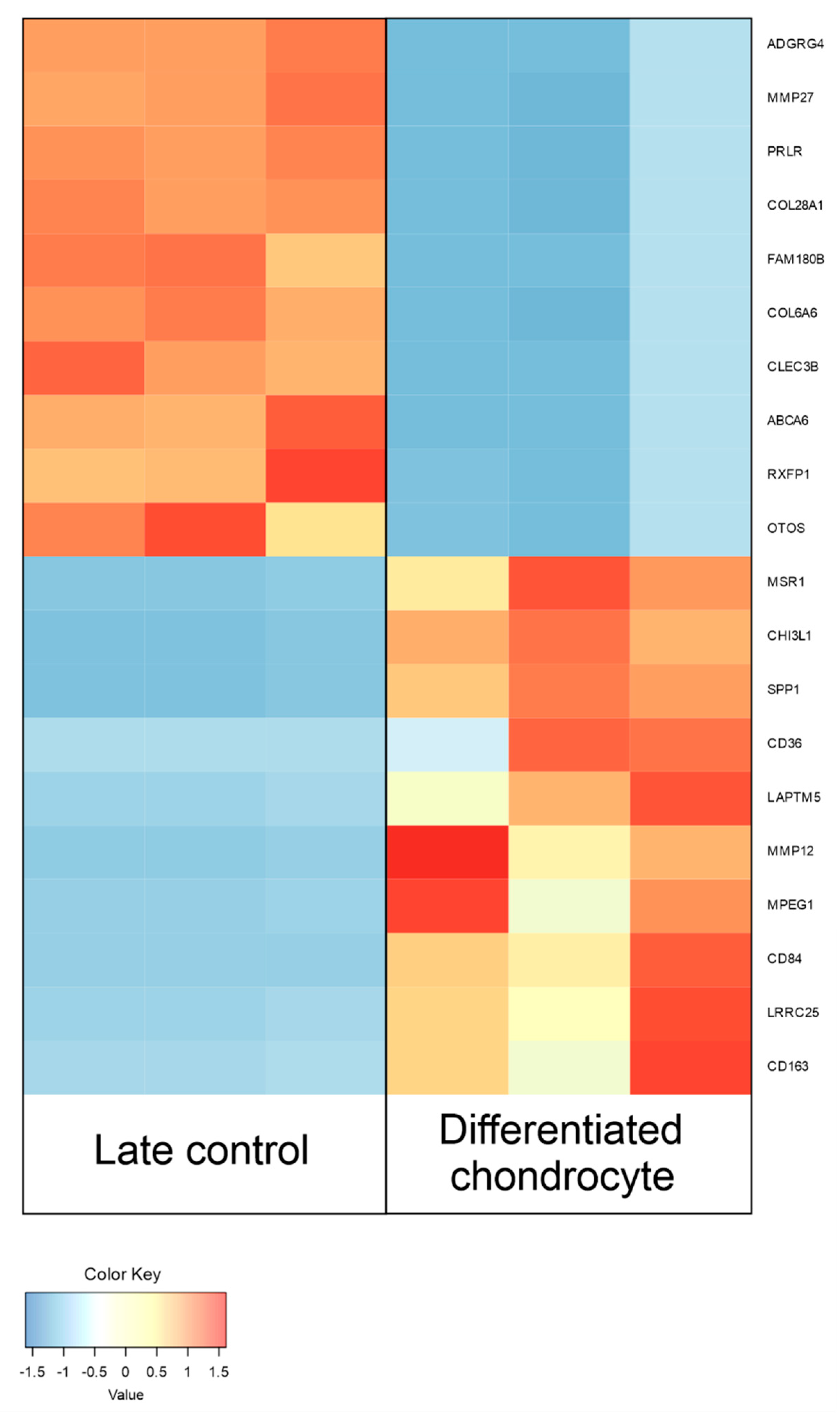
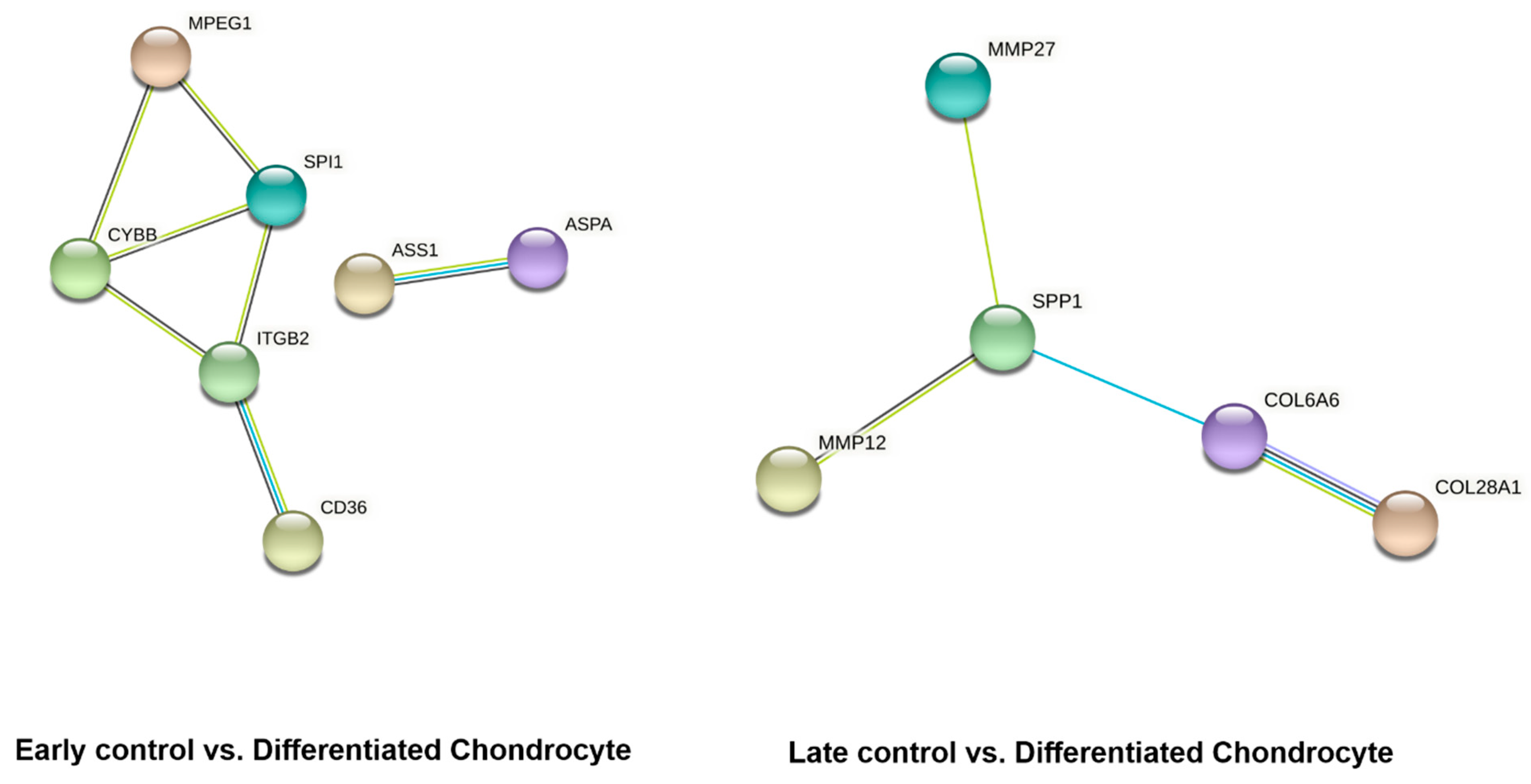
3.4. RNAseq Analysis
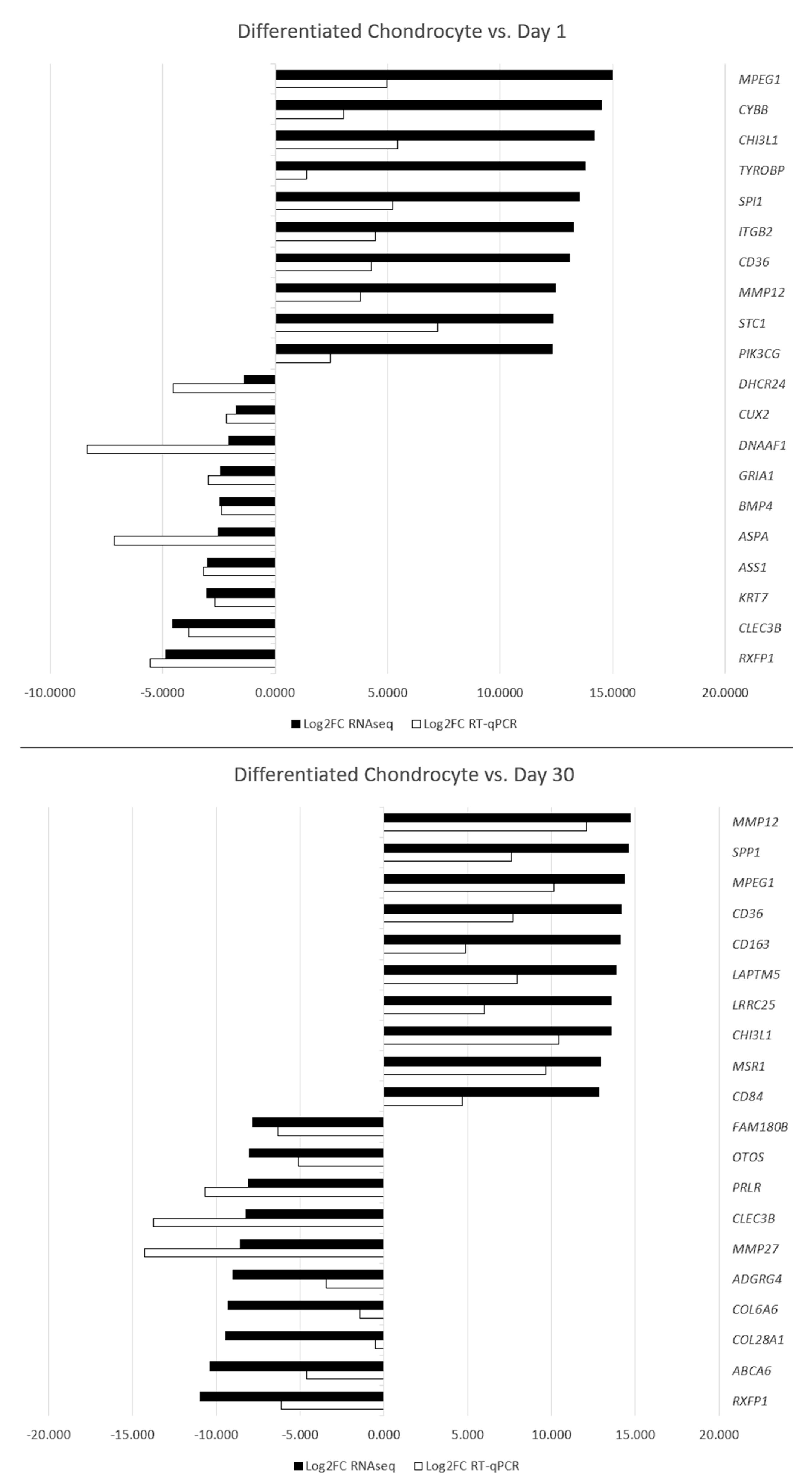
3.5. RT-qPCR Validation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Im, G.-I. Current Status of Regenerative Medicine in Osteoarthritis. Bone Jt. Res. 2021, 10, 134–136. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, P.; Flores, A.I. Current Status and Future Prospects of Perinatal Stem Cells. Genes 2020, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Moncrieff, L.; Mozdziak, P.; Jeseta, M.; Machatkova, M.; Kranc, W.; Kempisty, B. Ovarian Follicular Cells—Living in the Shadow of Stemness Cellular Competence. Med. J. Cell Biol. 2019, 7, 134–140. [Google Scholar] [CrossRef]
- Kulus, M.; Kulus, J.; Jankowski, M.; Borowiec, B.; Jeseta, M.; Bukowska, D.; Brüssow, K.P.; Kempisty, B.; Antosik, P. The Use of Mesenchymal Stem Cells in Veterinary Medicine. Med. J. Cell Biol. 2018, 6, 101–107. [Google Scholar] [CrossRef]
- Stefańska, K.; Sibiak, R.; Hutchings, G.; Dompe, C.; Moncrieff, L.; Janowicz, K.; Jeseta, M.; Kempisty, B.; Machatkova, M.; Mozdziak, P. Evidence for Existence of Molecular Stemness Markers in Porcine Ovarian Follicular Granulosa Cells. Med. J. Cell Biol. 2019, 7, 183–188. [Google Scholar] [CrossRef]
- Jankowski, M.; Dyszkiewicz-Konwińska, M.; Budna, J.; Knap, S.; Bryja, A.; Borys, S.; Kranc, W.; Magas, M.; Jeseta, M.; Bukowska, D.; et al. The Differentiation and Transdifferentiation of Epithelial Cells In Vitro—Is It a New Strategy in Regenerative Biomedicine? Med. J. Cell Biol. 2018, 6, 27–32. [Google Scholar] [CrossRef]
- Medvedev, S.P.; Shevchenko, A.I.; Zakian, S.M. Induced Pluripotent Stem Cells: Problems and Advantages When Applying Them in Regenerative Medicine. Acta Nat. 2010, 2, 18–27. [Google Scholar] [CrossRef]
- Jarrige, M.; Frank, E.; Herardot, E.; Martineau, S.; Darle, A.; Benabides, M.; Domingues, S.; Chose, O.; Habeler, W.; Lorant, J.; et al. The Future of Regenerative Medicine: Cell Therapy Using Pluripotent Stem Cells and Acellular Therapies Based on Extracellular Vesicles. Cells 2021, 10, 240. [Google Scholar] [CrossRef]
- Dompe, C.; Wasiatycz, G.; Mozdziak, P.; Jankowski, M.; Kempisty, B. Current Clinical Applications of Adipose-Derived Stem Cells in Humans and Animals. Med. J. Cell Biol. 2019, 7, 105–111. [Google Scholar] [CrossRef]
- Man, Y.; Yao, X.; Yang, T.; Wang, Y. Hematopoietic Stem Cell Niche During Homeostasis, Malignancy, and Bone Marrow Transplantation. Front. Cell Dev. Biol. 2021, 9, 14. [Google Scholar] [CrossRef]
- De la Morena, M.T.; Gatti, R.A. A History of Bone Marrow Transplantation. Hematol. Oncol. Clin. N. Am. 2011, 25, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Young, H.E.; Black, A.C. Adult Stem Cells. Anat. Rec.—Part A Discov. Mol. Cell. Evol. Biol. 2004, 276, 75–102. [Google Scholar] [CrossRef] [PubMed]
- Rojewska, M.; Popis, M.; Jankowski, M.; Bukowska, D.; Antosik, P.; Kempisty, B. Stemness Specificity of Epithelial Cells—Application of Cell and Tissue Technology in Regenerative Medicine. Med. J. Cell Biol. 2018, 6, 114–119. [Google Scholar] [CrossRef]
- Stefanska, K.; Bryl, R.; Moncrieff, L.; Pinto, N.; Shibli, J.A.; Dyszkiewicz-KonwiÅska, M. Mesenchymal Stem Cells—A Historical Overview. Med. J. Cell Biol. 2020, 8, 83–87. [Google Scholar] [CrossRef]
- Stefanska, K.; Bryl, R.; Hutchings, G.; Shibli, J.A.; Dyszkiewicz-KonwiÅska, M. Human Umbilical Cord Stem Cells-the Discovery, History and Possible Application. Med. J. Cell Biol. 2020, 8, 78–82. [Google Scholar] [CrossRef]
- Müller, M.; Czarnecka, J.; Brzeziński, M.; Prus, J.; Kulak, B.; Hołubowski, A.; Stasiak, M.; Borowiec, B.; Bryl, R.; Moncrieff, L.; et al. Current Stem Cells Technologies Used in Medicine. Med. J. Cell Biol. 2020, 8, 124–138. [Google Scholar] [CrossRef]
- Jankowski, M.; Dompe, C.; Sibiak, R.; Wąsiatycz, G.; Mozdziak, P.; Jaśkowski, J.M.; Antosik, P.; Kempisty, B.; Dyszkiewicz-Konwińska, M. In Vitro Cultures of Adipose-Derived Stem Cells: An Overview of Methods, Molecular Analyses, and Clinical Applications. Cells 2020, 9, 1783. [Google Scholar] [CrossRef]
- Cotman, C.W.; Head, E. The Canine (Dog) Model of Human Aging and Disease: Dietary, Environmental and Immunotherapy Approaches. J. Alzheimer’s Dis. 2008, 15, 685–707. [Google Scholar] [CrossRef]
- Hytönen, M.K.; Lohi, H. Canine Models of Human Rare Disorders. Rare Dis. 2016, 4, e1241362. [Google Scholar] [CrossRef]
- Mildmay-White, A.; Khan, W. Cell Surface Markers on Adipose-Derived Stem Cells: A Systematic Review. Curr. Stem Cell Res. Ther. 2017, 12, 484–492. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Chermuła, B.; Brazert, M.; Jeseta, M.; Ożegowska, K.; Sujka-Kordowska, P.; Konwerska, A.; Bryja, A.; Kranc, W.; Jankowski, M.; Nawrocki, M.J.; et al. The Unique Mechanisms of Cellular Proliferation, Migration and Apoptosis Are Regulated through Oocyte Maturational Development—A Complete Transcriptomic and Histochemical Study. Int. J. Mol. Sci. 2019, 20, 84. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lei, R.; Ding, S.W.; Zhu, S. Skewer: A Fast and Accurate Adapter Trimmer for next-Generation Sequencing Paired-End Reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef]
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3-Masker: Integrating Masking of Template Sequence with Primer Design Software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An Improvement of the 2ˆ(-Delta Delta CT) Method for Quantitative Real-Time Polymerase Chain Reaction Data Analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Ge, S.X.; Son, E.W.; Yao, R. IDEP: An Integrated Web Application for Differential Expression and Pathway Analysis of RNA-Seq Data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Jankowski, M.; Kaczmarek, M.; Wąsiatycz, G.; Dompe, C.; Mozdziak, P.; Jaśkowski, J.M.; Piotrowska-Kempisty, H.; Kempisty, B. Expression Profile of New Marker Genes Involved in Differentiation of Canine Adipose-Derived Stem Cells into Osteoblasts. Int. J. Mol. Sci. 2021, 22, 6663. [Google Scholar] [CrossRef]
- Dyszkiewicz-Konwińska, M.; Nawrocki, M.; Huang, Y.; Bryja, A.; Celichowski, P.; Jankowski, M.; Błochowiak, K.; Mehr, K.; Bruska, M.; Nowicki, M.; et al. New Gene Markers for Metabolic Processes and Homeostasis in Porcine Buccal Pouch Mucosa during Cells Long Term-Cultivation—A Primary Culture Approach. Int. J. Mol. Sci. 2018, 19, 1027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kranc, W.; Brązert, M.; Ożegowska, K.; Nawrocki, M.M.J.; Budna, J.; Celichowski, P.; Dyszkiewicz-Konwińska, M.; Jankowski, M.; Jeseta, M.; Pawelczyk, L.; et al. Expression Profile of Genes Regulating Steroid Biosynthesis and Metabolism in Human Ovarian Granulosa Cells—A Primary Culture Approach. Int. J. Mol. Sci. 2017, 18, 2673. [Google Scholar] [CrossRef] [PubMed]
- Ku, C.S.; Naidoo, N.; Wu, M.; Soong, R. Studying the Epigenome Using next Generation Sequencing. J. Med. Genet. 2011, 48, 721–730. [Google Scholar] [CrossRef] [PubMed]
- James, C.G.; Appleton, C.T.G.; Ulici, V.; Michael Underhill, T.; Beier, F. Microarray Analyses of Gene Expression during Chondrocyte Differentiation Identifies Novel Regulators of Hypertrophy. Mol. Biol. Cell 2005, 16, 5316–5333. [Google Scholar] [CrossRef]
- Di Rosa, M.; Szychlinska, M.A.; Tibullo, D.; Malaguarnera, L.; Musumeci, G. Expression of CHI3L1 and CHIT1 in Osteoarthritic Rat Cartilage Model. A Morphological Study. Eur. J. Histochem. 2014, 58, 213–221. [Google Scholar] [CrossRef]
- Szychlinska, M.A.; Trovato, F.M.; Di Rosa, M.; Malaguarnera, L.; Puzzo, L.; Leonardi, R.; Castrogiovanni, P.; Musumeci, G. Co-Expression and Co-Localization of Cartilage Glycoproteins CHI3L1 and Lubricin in Osteoarthritic Cartilage: Morphological, Immunohistochemical and Gene Expression Profiles. Int. J. Mol. Sci. 2016, 17, 359. [Google Scholar] [CrossRef]
- LaPointe, V.L.S.; Verpoorte, A.; Stevens, M.M. The Changing Integrin Expression and a Role for Integrin Β8 in the Chondrogenic Differentiation of Mesenchymal Stem Cells. PLoS ONE 2013, 8, e82035. [Google Scholar] [CrossRef]
- Bendele, F.; Beier, R.; Terkeltaub, J.S.; Mort, A.M.; Schmidt, A.; Denise, L.; Cecil, C.; Thomas, G.; Appleton, M.D. The Pattern Recognition Receptor CD36 Is a Chondrocyte Hypertrophy Marker Associated with Suppression of Catabolic Responses and Promotion of Repair Responses to Inflammatory Stimuli. J. Immunol. 2009, 182, 5024–5031. [Google Scholar] [CrossRef]
- KerkelÄ, E.; Böhling, T.; Herva, R.; Uria, J.A.; Saarialho-Kere, U. Human Macrophage Metalloelastase (MMP-12) Expression Is Induced in Chondrocytes during Fetal Development and Malignant Transformation. Bone 2001, 29, 487–493. [Google Scholar] [CrossRef]
- Wu, S.; Yoshiko, Y.; De Luca, F. Stanniocalcin 1 Acts as a Paracrine Regulator of Growth Plate Chondrogenesis*. J. Biol. Chem. 2006, 281, 5120–5127. [Google Scholar] [CrossRef]
- Ramesova, A.; Vesela, B.; Svandova, E.; Lesot, H.; Matalova, E. Caspase-9 Inhibition Decreases Expression of Mmp9 during Chondrogenesis. Histochem. Cell Biol. 2022, 157, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.A.; Kalsoom, K.; Iqbal, A.; Asif, H.; Rahman, H.; Farooq, S.O.; Naveed, H.; Nasir, U.; Amin, M.U.; Hussain, M.; et al. A Novel Missense Mutation in the NADPH Binding Domain of CYBB Abolishes the NADPH Oxidase Activity in a Male Patient with Increased Susceptibility to Infections. Microb. Pathog. 2016, 100, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jiang, T.; Tan, L.; Yu, J.T. TYROBP in Alzheimer’s Disease. Mol. Neurobiol. 2015, 51, 820–826. [Google Scholar] [CrossRef]
- Liang, T.; Chen, J.; Xu, G.Y.; Zhang, Z.; Xue, J.; Zeng, H.; Jiang, J.; Chen, T.; Qin, Z.; Li, H.; et al. STAT3 and SPI1, May Lead to the Immune System Dysregulation and Heterotopic Ossification in Ankylosing Spondylitis. BMC Immunol. 2022, 23, 3. [Google Scholar] [CrossRef]
- Mirza, R.; Qiao, S.; Tateyama, K.; Miyamoto, T.; Xiuli, L.; Seo, H. 3β-Hydroxysterol-Delta24 Reductase Plays an Important Role in Long Bone Growth by Protecting Chondrocytes from Reactive Oxygen Species. J. Bone Miner. Metab. 2012, 30, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Jang, Y.; Jung, H.; Nam, Y.; Rim, Y.A.; Kim, J.; Jeong, S.H.; Ju, J.H. Centrifugal Gravity-Induced BMP4 Induces Chondrogenic Differentiation of Adipose-Derived Stem Cells via SOX9 Upregulation. Stem Cell Res. Ther. 2016, 7, 184. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Kang, K.S.; Kang, S.K. Soluble Factors from ASCs Effectively Direct Control of Chondrogenic Fate. Cell Prolif. 2010, 43, 249–261. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but Not BMP4, Is Crucial for Chondrocyte Proliferation and Maturation during Endochondral Bone Development. J. Cell Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef]
- Sarem, M.; Otto, O.; Tanaka, S.; Shastri, V.P. Cell Number in Mesenchymal Stem Cell Aggregates Dictates Cell Stiffness and Chondrogenesis. Stem Cell Res. Ther. 2019, 10, 10. [Google Scholar] [CrossRef]
- Mishra, P.J.; Mishra, P.J.; Humeniuk, R.; Medina, D.J.; Alexe, G.; Mesirov, J.P.; Ganesan, S.; Glod, J.W.; Banerjee, D. Carcinoma-Associated Fibroblast–Like Differentiation of Human Mesenchymal Stem Cells. Cancer Res. 2008, 68, 4331–4339. [Google Scholar] [CrossRef]
- Steinberg, J.; Ritchie, G.R.S.; Roumeliotis, T.I.; Jayasuriya, R.L.; Clark, M.J.; Brooks, R.A.; Binch, A.L.A.; Shah, K.M.; Coyle, R.; Pardo, M.; et al. Integrative Epigenomics, Transcriptomics and Proteomics of Patient Chondrocytes Reveal Genes and Pathways Involved in Osteoarthritis. Sci. Rep. 2017, 7, 8935. [Google Scholar] [CrossRef] [Green Version]
- Gingras, H.; Cases, O.; Krasilnikova, M.; Bérubé, G.; Nepveu, A. Biochemical Characterization of the Mammalian Cux2 Protein. Gene 2005, 344, 273–285. [Google Scholar] [CrossRef]
- Hartill, V.L.; van de Hoek, G.; Patel, M.P.; Little, R.; Watson, C.M.; Berry, I.R.; Shoemark, A.; Abdelmottaleb, D.; Parkes, E.; Bacchelli, C.; et al. DNAAF1 Links Heart Laterality with the AAA+ ATPase RUVBL1 and Ciliary Intraflagellar Transport. Hum. Mol. Genet. 2018, 27, 529–545. [Google Scholar] [CrossRef]
- Miao, C.; Jiang, Q.; Li, H.; Zhang, Q.; Bai, B.; Bao, Y.; Zhang, T. Mutations in the Motile Cilia Gene DNAAF1 Are Associated with Neural Tube Defects in Humans. G3 Genes Genomes Genet. 2016, 6, 3307–3316. [Google Scholar] [CrossRef]
- Chiesa, A.; Crisafulli, C.; Porcelli, S.; Han, C.; Patkar, A.A.; Lee, S.J.; Park, M.H.; Jun, T.Y.; Serretti, A.; Pae, C.U. Influence of GRIA1, GRIA2 and GRIA4 Polymorphisms on Diagnosis and Response to Treatment in Patients with Major Depressive Disorder. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 305–311. [Google Scholar] [CrossRef]
- Kerner, B.; Jasinska, A.J.; DeYoung, J.; Almonte, M.; Choi, O.W.; Freimer, N.B. Polymorphisms in the GRIA1 Gene Region in Psychotic Bipolar Disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2009, 150, 24–32. [Google Scholar] [CrossRef]
- Tilley, S.K.; Kim, W.Y.; Fry, R.C. Analysis of Bladder Cancer Tumor CpG Methylation and Gene Expression within The Cancer Genome Atlas Identifies GRIA1 as a Prognostic Biomarker for Basal-like Bladder Cancer. Am. J. Cancer Res. 2017, 7, 1850. [Google Scholar]
- Moffett, J.R.; Arun, P.; Ariyannur, P.S.; Garbern, J.Y.; Jacobowitz, D.M.; Namboodiri, A.M.A. Extensive Aspartoacylase Expression in the Rat Central Nervous System. Glia 2011, 59, 1414–1434. [Google Scholar] [CrossRef]
- Chen, T.Y.; Li, X.; Hung, C.H.; Bahudhanapati, H.; Tan, J.; Kass, D.J.; Zhang, Y. The Relaxin Family Peptide Receptor 1 (RXFP1): An Emerging Player in Human Health and Disease. Mol. Genet. Genom. Med. 2020, 8, e1194. [Google Scholar] [CrossRef]
- Valenti, M.T.; Carbonare, L.D.; Donatelli, L.; Bertoldo, F.; Zanatta, M.; Lo Cascio, V. Gene Expression Analysis in Osteoblastic Differentiation from Peripheral Blood Mesenchymal Stem Cells. Bone 2008, 43, 1084–1092. [Google Scholar] [CrossRef]
- Jiao, K.; Zhang, J.; Zhang, M.; Wei, Y.; Wu, Y.; Qiu, Z.Y.; He, J.; Cao, Y.; Hu, J.; Zhu, H.; et al. The Identification of CD163 Expressing Phagocytic Chondrocytes in Joint Cartilage and Its Novel Scavenger Role in Cartilage Degradation. PLoS ONE 2013, 8, e53312. [Google Scholar] [CrossRef]
- Du, Y.; Duan, T.; Feng, Y.; Liu, Q.; Lin, M.; Cui, J.; Wang, R.-F. LRRC25 Inhibits Type I IFN Signaling by Targeting ISG15-Associated RIG-I for Autophagic Degradation. EMBO J. 2018, 37, 351–366. [Google Scholar] [CrossRef]
- Wu, L.; Bluguermann, C.; Kyupelyan, L.; Latour, B.; Gonzalez, S.; Shah, S.; Galic, Z.; Ge, S.; Zhu, Y.; Petrigliano, F.A.; et al. Human Developmental Chondrogenesis as a Basis for Engineering Chondrocytes from Pluripotent Stem Cells. Stem Cell Rep. 2013, 1, 575–589. [Google Scholar] [CrossRef]
- Ogueta, S.; Muoz, J.; Obregon, E.; Delgado-Baeza, E.; García-Ruiz, J.P. Prolactin Is a Component of the Human Synovial Liquid and Modulates the Growth and Chondrogenic Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells. Mol. Cell. Endocrinol. 2002, 190, 51–63. [Google Scholar] [CrossRef]
- Seriwatanachai, D.; Krishnamra, N.; Charoenphandhu, N. Chondroregulatory Action of Prolactin on Proliferation and Differentiation of Mouse Chondrogenic ATDC5 Cells in 3-Dimensional Micromass Cultures. Biochem. Biophys. Res. Commun. 2012, 420, 108–113. [Google Scholar] [CrossRef]
- Cui, Y.; Yu, J.; Urban, J.P.G.; Young, D.A. Differential Gene Expression Profiling of Metalloproteinases and Their Inhibitors: A Comparison between Bovine Intervertebral Disc Nucleus Pulposus Cells and Articular Chondrocytes. Spine 2010, 35, 1101–1108. [Google Scholar] [CrossRef]
- Ozkul, Y.; Galderisi, U. The Impact of Epigenetics on Mesenchymal Stem Cell Biology. J. Cell. Physiol. 2016, 231, 2393–2401. [Google Scholar] [CrossRef]
- Horton, E.R.; Vallmajo-Martin, Q.; Martin, I.; Snedeker, J.G.; Ehrbar, M.; Blache, U. Extracellular Matrix Production by Mesenchymal Stromal Cells in Hydrogels Facilitates Cell Spreading and Is Inhibited by FGF-2. Adv. Healthc. Mater. 2020, 9, 1901669. [Google Scholar] [CrossRef]
- Barbet, R.; Peiffer, I.; Hutchins, J.R.A.; Hatzfeld, A.; Garrido, E.; Hatzfeld, J.A. Expression of the 49 Human ATP Binding Cassette (ABC) Genes in Pluripotent Embryonic Stem Cells and in Early- and Late-Stage Multipotent Mesenchymal Stem Cells: Possible Role of ABC Plasma Membrane Transporters in Maintaining Human Stem Cell Pluripotency. Cell Cycle 2012, 11, 1611–1620. [Google Scholar] [CrossRef]
- Sun, H.; Huang, A.; Cao, S. Current Status and Prospects of Gene Therapy for the Inner Ear. Hum. Gene Ther. 2011, 22, 1311–1322. [Google Scholar] [CrossRef]
- Xiao, P.; Guo, S.; Wen, X.; He, Q.T.; Lin, H.; Huang, S.M.; Gou, L.; Zhang, C.; Yang, Z.; Zhong, Y.N.; et al. Tethered Peptide Activation Mechanism of the Adhesion GPCRs ADGRG2 and ADGRG4. Nature 2022, 604, 771–778. [Google Scholar] [CrossRef]
- Chung, J.; Marini, S.; Pera, J.; Norrving, B.; Jimenez-Conde, J.; Roquer, J.; Fernandez-Cadenas, I.; Tirschwell, D.L.; Selim, M.; Brown, D.L.; et al. Genome-Wide Association Study of Cerebral Small Vessel Disease Reveals Established and Novel Loci. Brain 2019, 142, 3176. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GENE NAME | FORWARD PRIMER | REVERSE PRIMER | ENSEMBL ID |
|---|---|---|---|
| ABCA6 | GGTCAACTTCCTGGGCTACT | TTCGCCTGAACATTGAGCTG | ENSCAFG00845005313 |
| ACTB | TCGAGACTTTCAACACCCCA | CATGAGGTAGTCGGTCAGGT | ENSCAFG00030015381 |
| ADGRG4 | TACAGCCTTGACTCTTGGGG | CCAGACTCAGAGGCCTTGAA | ENSCAFG00000018940 |
| ASPA | CAAGGGGTTCTGAGAGCTGA | GCGGTTTCCAGTCTTGATCC | ENSCAFG00000019330 |
| ASS1 | GTGTGAATTTGTCCGCCACT | TCTGGAGGCGGTGATATTCC | ENSCAFG00030010628 |
| BMP4 | GAGAAGCAGCCAAACTACGG | CTTATTCTTCTTGCGGGCCC | ENSCAFG00030003207 |
| CD163 | CTCTGCAACTCTCACTGGGA | CAATCTCCCATGTGCTGCTC | ENSCAFG00030019149 |
| CD36 | CAGGAAGTGGTTGCGAACAG | AGCCAGATTGAGAACGGTCA | ENSCAFG00030010230 |
| CD84 | AACATACAGCTGGAGTCCCC | ACAGAGAGAGCATGACCAGC | ENSCAFG00000012569 |
| CHI3L1 | CACGTCATCTACAGCTTCGC | ACCAAAGCTCCATCCTCCAA | ENSCAFG00030003216 |
| CLEC3B | AAATGCTTCCTGGCCTTCAC | CTCCGTCTCCCAGTTCTTGT | ENSCAFG00000014049 |
| COL28A1 | GGGACAAGGGAGATTTGGGA | GTCTGGCCTACTTCACCCTT | ENSCAFG00000029367 |
| COL6A6 | CTTCCGGGAGAGATGGGATC | TTCATGCGCTCGAATTCCTG | ENSCAFG00000006035 |
| CUX2 | GGGACCCAAGATGAACCAGA | CCGCTTCTTCTTCTGCATCC | ENSCAFG00000008575 |
| CYBB | CTGAGCGAATTGTACGTGGG | GAGATCGCCAAAACCGTACC | ENSCAFG00030009532 |
| DHCR24 | AAGCAGGTACGGGAATGGAA | GTCCACCTCCAGAATGTCCA | ENSCAFG00040000097 |
| DNAAF1 | ACCCAAGCAAGCAGAAACAG | TCCAGCCAGAGACAACGTAG | ENSCAFG00000019964 |
| FAM180B | AGTTCCAGGACCTGCGTAAA | GGGAAAGGGGTCAAGGATCA | ENSCAFG00000032106 |
| GRIA1 | TGACATTTCTCCCAGGTCCC | CTAGGTCCTCAGCACTCTCG | ENSCAFG00030007058 |
| HPRT1 | CCCAGCGTCGTGATTAGTGA | AGAGGGCTACGATGTGATGG | ENSCAFG00030008563 |
| ITGB2 | ATCAACGTCCCGATCACCTT | TCGCAGTTCTTCCCGATGTA | ENSCAFG00000011039 |
| KRT7 | TTCGCCTCCTTCATCGACAA | CGAAGATGCTGGGGAGGC | ENSCAFG00030022127 |
| LAPTM5 | TCCAAGGTCCCACTGATGAC | TCCACACGCACTTGAACATG | ENSCAFG00030004981 |
| LRRC25 | TGGTTCTAGGTCTGTGGCTG | GGGCCCTCGTAGTTCATGTA | ENSCAFG00000014879 |
| MMP12 | AGATTCTTGTGGTGGAGGCA | TGGCTGTGGTCTCAAATTGC | ENSCAFG00030013517 |
| MMP27 | TTCCCAAACCCATCCGTACA | CTGGAAAGCAGCATCGACTC | ENSCAFG00000015066 |
| MPEG1 | GAGGTCAAGGGAGAAGGGAC | GGGTGCACACGTGTATGATC | ENSCAFG00000007649 |
| MSR1 | ATGCTCGTTCAATGACAGCC | GTGCTGCCATGATTCCGATG | ENSCAFG00030021708 |
| OTOS | CCTACTGGCCTTTCTCCACT | CTGCTGATAGGGGACATGGA | ENSCAFG00000031916 |
| PIK3CG | AGGCAGCTGTGGAGAGATTT | AGGAAGTCTGGGGTTAGCAC | ENSCAFG00030020245 |
| PRLR | TGGGCAGCAGACTCAGTTTA | ATGACAGCAGAAAGAACGGC | ENSCAFG00040010421 |
| RXFP1 | AGGTCTGAGAACAAGCTGCA | AGATCCCACAAGCTGACAGT | ENSCAFG00000008672 |
| SPI1 | GACTATCTCCCAGTGGCAGG | TTTGCACGCCTGTAACATCC | ENSCAFG00000008723 |
| SPP1 | TATTCACTCCAGCTGTCCCC | TGTCTTTTGCATGGCTGTCC | ENSCAFG00000009569 |
| STC1 | TTCTGTGAGCCCCAGGAAAT | CAGCGCTGTACAAGAAGGAT | ENSCAFG00000009104 |
| TYROBP | CAACTGCCCCGTGGTGAG | GATGCGCTGTTTCCTGGTC | ENSCAFG00030007988 |
| Differentiated Chondrocytes vs. Day 1 | |||
|---|---|---|---|
| Gene Symbol | Log2FC | adj. p Value | Entrez Gene ID |
| MPEG1 | 14.9622 | 9.90 × 103 | 475960 |
| CYBB | 14.4821 | 4.24 × 102 | 491825 |
| CHI3L1 | 14.1706 | 9.09 × 106 | 490222 |
| TYROBP | 13.7521 | 5.66 × 104 | 476477 |
| SPI1 | 13.5224 | 1.59 × 103 | 611255 |
| ITGB2 | 13.2486 | 1.11 × 102 | 403770 |
| CD36 | 13.0788 | 4.36 × 102 | 475931 |
| MMP12 | 12.4532 | 2.41 × 103 | 611789 |
| STC1 | 12.3310 | 1.92 × 102 | 486112 |
| PIK3CG | 12.3146 | 6.00 × 103 | 483266 |
| DHCR24 | −1.3651 | 1.65 × 104 | 489573 |
| CUX2 | −1.7343 | 3.13 × 104 | 486267 |
| DNAAF1 | −2.0651 | 4.95 × 102 | 479628 |
| GRIA1 | −2.4020 | 2.51 × 102 | 489168 |
| BMP4 | −2.4524 | 6.73 × 103 | 490695 |
| ASPA | −2.5116 | 2.23 × 102 | 611064 |
| ASS1 | −2.9978 | 1.37 × 102 | 480693 |
| KRT7 | −3.0224 | 2.13 × 102 | 477602 |
| CLEC3B | −4.5737 | 2.69 × 103 | 609596 |
| RXFP1 | −4.8416 | 1.36 × 102 | 100855494 |
| Differentiated Chondrocyte vs. Day 30 | |||
|---|---|---|---|
| Gene Symbol | Log2FC | adj. p Value | Entrez Gene ID |
| MMP12 | 14.673 | 2.36 × 103 | 611789 |
| SPP1 | 14.597 | 4.75 × 105 | 478471 |
| MPEG1 | 14.369 | 9.75 × 103 | 475960 |
| CD36 | 14.138 | 4.26 × 102 | 475931 |
| CD163 | 14.096 | 2.81 × 102 | 477704 |
| LAPTM5 | 13.879 | 1.58 × 102 | 487324 |
| LRRC25 | 13.590 | 1.83 × 102 | 609889 |
| CHI3L1 | 13.562 | 6.28 × 106 | 490222 |
| MSR1 | 12.912 | 6.36 × 104 | 482891 |
| CD84 | 12.843 | 6.40 × 103 | 488641 |
| FAM180B | −7.843 | 1.89 × 105 | 100685781 |
| OTOS | −8.031 | 2.57 × 104 | 477428 |
| PRLR | −8.092 | 1.70 × 106 | 479363 |
| CLEC3B | −8.239 | 2.67 × 105 | 609596 |
| MMP27 | −8.569 | 6.44 × 106 | 489430 |
| ADGRG4 | −8.987 | 3.93 × 106 | 492163 |
| COL6A6 | −9.286 | 2.61 × 106 | 610649 |
| COL28A1 | −9.442 | 7.12 × 107 | 482315 |
| ABCA6 | −10.357 | 4.77 × 105 | 480456 |
| RXFP1 | −10.948 | 1.65 × 104 | 100855494 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankowski, M.; Kaczmarek, M.; Wąsiatycz, G.; Konwerska, A.; Dompe, C.; Bukowska, D.; Antosik, P.; Mozdziak, P.; Kempisty, B. Expression Profile of New Gene Markers Involved in Differentiation of Canine Adipose-Derived Stem Cells into Chondrocytes. Genes 2022, 13, 1664. https://doi.org/10.3390/genes13091664
Jankowski M, Kaczmarek M, Wąsiatycz G, Konwerska A, Dompe C, Bukowska D, Antosik P, Mozdziak P, Kempisty B. Expression Profile of New Gene Markers Involved in Differentiation of Canine Adipose-Derived Stem Cells into Chondrocytes. Genes. 2022; 13(9):1664. https://doi.org/10.3390/genes13091664
Chicago/Turabian StyleJankowski, Maurycy, Mariusz Kaczmarek, Grzegorz Wąsiatycz, Aneta Konwerska, Claudia Dompe, Dorota Bukowska, Paweł Antosik, Paul Mozdziak, and Bartosz Kempisty. 2022. "Expression Profile of New Gene Markers Involved in Differentiation of Canine Adipose-Derived Stem Cells into Chondrocytes" Genes 13, no. 9: 1664. https://doi.org/10.3390/genes13091664