
Differences in Adipose Gene Expression Profiles between Male and Female Even Reindeer (Rangifer tarandus) in Sakha (Yakutia)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection for Transcriptome Analysis
2.2. RNA Extraction, Library Preparation, and Sequencing
2.3. Bioinformatic Analyses
3. Results and Discussion
3.1. RNA Sequencing and Mapping
3.2. Gene Expression Overview
3.3. Uniquely Expressed Genes
3.4. Differential Gene Expression Analyses
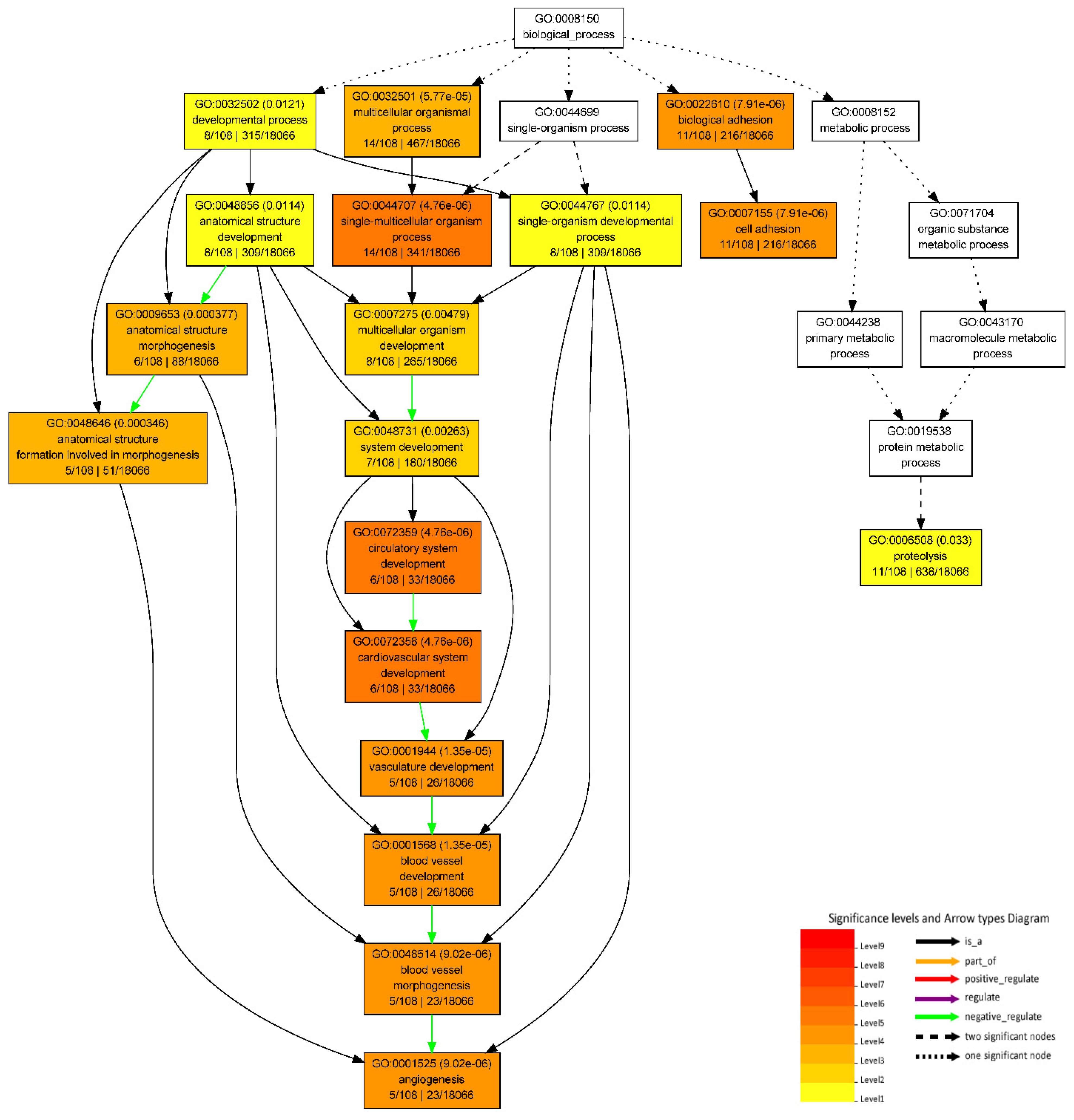
3.5. GO Enrichment Analyses
3.6. KEGG Pathway Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pond, C.M. An Evolutionary and Functional View of Mammalian Adipose Tissue. Proc. Nutr. Soc. 1992, 51, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S. Central Actions of Adipocyte Hormones. Trends Endocrinol. Metab. 2005, 16, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Niedźwiedzka-Rystwej, P.; Deptuła, W. Review PaperAdipose Tissue—Component of the Immune System. Cent. Eur. J. OfnbspImmunol. 2011, 36, 95–99. [Google Scholar]
- Palmer, B.F.; Clegg, D.J. The Sexual Dimorphism of Obesity. Mol. Cell. Endocrinol. 2015, 402, 113–119. [Google Scholar] [CrossRef]
- Chang, E.; Varghese, M.; Singer, K. Gender and Sex Differences in Adipose Tissue. Curr. Diabetes Rep. 2018, 18, 69. [Google Scholar] [CrossRef]
- Power, M.L.; Schulkin, J. Sex Differences in Fat Storage, Fat Metabolism, and the Health Risks from Obesity: Possible Evolutionary Origins. Br. J. Nutr. 2008, 99, 931–940. [Google Scholar] [CrossRef]
- Michaud, A.; Drolet, R.; Noël, S.; Paris, G.; Tchernof, A. Visceral Fat Accumulation Is an Indicator of Adipose Tissue Macrophage Infiltration in Women. Metab. Clin. Exp. 2012, 61, 689–698. [Google Scholar] [CrossRef]
- Fuente-Martín, E.; Argente-Arizón, P.; Ros, P.; Argente, J.; Chowen, J.A. Sex Differences in Adipose Tissue. Adipocyte 2013, 2, 128–134. [Google Scholar] [CrossRef]
- Fletcher, L.A.; Kim, K.; Leitner, B.P.; Cassimatis, T.M.; O’Mara, A.E.; Johnson, J.W.; Halprin, M.S.; McGehee, S.M.; Brychta, R.J.; Cypess, A.M.; et al. Sexual Dimorphisms in Adult Human Brown Adipose Tissue. Obesity 2020, 28, 241–246. [Google Scholar] [CrossRef]
- Le Magueresse-Battistoni, B. Adipose Tissue and Endocrine-Disrupting Chemicals: Does Sex Matter? Int. J. Environ. Res. Public. Health 2020, 17, 9403. [Google Scholar] [CrossRef]
- Karastergiou, K.; Smith, S.R.; Greenberg, A.S.; Fried, S.K. Sex Differences in Human Adipose Tissues—The Biology of Pear Shape. Biol. Sex Differ. 2012, 3, 13. [Google Scholar] [CrossRef]
- Mongraw-Chaffin, M.L.; Anderson, C.A.M.; Allison, M.A.; Ouyang, P.; Szklo, M.; Vaidya, D.; Woodward, M.; Golden, S.H. Association Between Sex Hormones and Adiposity: Qualitative Differences in Women and Men in the Multi-Ethnic Study of Atherosclerosis. J. Clin. Endocrinol. Metab. 2015, 100, E596–E600. [Google Scholar] [CrossRef] [PubMed]
- Ter Horst, K.W.; Gilijamse, P.W.; de Weijer, B.A.; Kilicarslan, M.; Ackermans, M.T.; Nederveen, A.J.; Nieuwdorp, M.; Romijn, J.A.; Serlie, M.J. Sexual Dimorphism in Hepatic, Adipose Tissue, and Peripheral Tissue Insulin Sensitivity in Obese Humans. Front. Endocrinol. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xie, M.; Yuan, S.; Zeng, Y.; Dong, Y.; Wang, Z.; Xiao, Q.; Dong, B.; Ma, J.; Hu, J. Sex Differences in the Associations between Adiposity Distribution and Cardiometabolic Risk Factors in Overweight or Obese Individuals: A Cross-Sectional Study. BMC Public Health 2021, 21, 1232. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; McClusky, R.; Chen, J.; Beaven, S.W.; Tontonoz, P.; Arnold, A.P.; Reue, K. The Number of X Chromosomes Causes Sex Differences in Adiposity in Mice. PLoS Genet. 2012, 8, e1002709. [Google Scholar] [CrossRef]
- Anderson, W.D.; Soh, J.Y.; Innis, S.E.; Dimanche, A.; Ma, L.; Langefeld, C.D.; Comeau, M.E.; Das, S.K.; Schadt, E.E.; Björkegren, J.L.M.; et al. Sex Differences in Human Adipose Tissue Gene Expression and Genetic Regulation Involve Adipogenesis. Genome Res. 2020, 30, 1379–1392. [Google Scholar] [CrossRef]
- Iglesias, M.J.; Eiras, S.; Piñeiro, R.; López-Otero, D.; Gallego, R.; Fernández, A.L.; Lago, F.; González-Juanatey, J.R. Gender differences in adiponectin and leptin expression in epicardial and subcutaneous adipose tissue. Findings in patients undergoing cardiac surgery. Rev. Esp. Cardiol. 2006, 59, 1252–1260. [Google Scholar] [CrossRef]
- Viguerie, N.; Montastier, E.; Maoret, J.-J.; Roussel, B.; Combes, M.; Valle, C.; Villa-Vialaneix, N.; Iacovoni, J.S.; Martinez, J.A.; Holst, C.; et al. Determinants of Human Adipose Tissue Gene Expression: Impact of Diet, Sex, Metabolic Status, and Cis Genetic Regulation. PLoS Genet. 2012, 8, e1002959. [Google Scholar] [CrossRef]
- Nookaew, I.; Svensson, P.-A.; Jacobson, P.; Jernås, M.; Taube, M.; Larsson, I.; Andersson-Assarsson, J.C.; Sjöström, L.; Froguel, P.; Walley, A.; et al. Adipose Tissue Resting Energy Expenditure and Expression of Genes Involved in Mitochondrial Function Are Higher in Women than in Men. J. Clin. Endocrinol. Metab. 2013, 98, E370–E378. [Google Scholar] [CrossRef]
- Link, J.C.; Hasin-Brumshtein, Y.; Cantor, R.M.; Chen, X.; Arnold, A.P.; Lusis, A.J.; Reue, K. Diet, Gonadal Sex, and Sex Chromosome Complement Influence White Adipose Tissue MiRNA Expression. BMC Genom. 2017, 18, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianconi, E.; Casadei, R.; Frabetti, F.; Ventura, C.; Facchin, F.; Canaider, S. Sex-Specific Transcriptome Differences in Human Adipose Mesenchymal Stem Cells. Genes 2020, 11, 909. [Google Scholar] [CrossRef] [PubMed]
- Kaikaew, K.; Grefhorst, A.; Steenbergen, J.; Swagemakers, S.M.A.; McLuskey, A.; Visser, J.A. Sex Difference in the Mouse BAT Transcriptome Reveals a Role of Progesterone. J. Mol. Endocrinol. 2021, 66, 97–113. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Messa, L.; Pandini, C.; Maghraby, E.; Barzaghini, B.; Garofalo, M.; Micheletto, G.; Raimondi, M.T.; Bertoli, S.; Cereda, C.; et al. RNA-Seq Characterization of Sex-Differences in Adipose Tissue of Obesity Affected Patients: Computational Analysis of Differentially Expressed Coding and Non-Coding RNAs. J. Pers. Med. 2021, 11, 352. [Google Scholar] [CrossRef] [PubMed]
- Weldenegodguad, M.; Pokharel, K.; Niiranen, L.; Soppela, P.; Ammosov, I.; Honkatukia, M.; Lindeberg, H.; Peippo, J.; Reilas, T.; Mazzullo, N.; et al. Adipose Gene Expression Profiles Reveal Insights into the Adaptation of Northern Eurasian Semi-Domestic Reindeer (Rangifer tarandus). Commun. Biol. 2021, 4, 1170. [Google Scholar] [CrossRef]
- Andrews, S. FASTQC. A Quality Control Tool for High Throughput Sequence Data. Available online: https://github.com/s-andrews/FastQC (accessed on 18 September 2020).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed]
- Weldenegodguad, M.; Pokharel, K.; Ming, Y.; Honkatukia, M.; Peippo, J.; Reilas, T.; Røed, K.H.; Kantanen, J. Genome Sequence and Comparative Analysis of Reindeer (Rangifer tarandus) in Northern Eurasia. Sci. Rep. 2020, 10, 8980. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, J.; Li, C.; Wang, Y.; Li, L.; Cai, H.; Lan, X.; Lei, C.; Zhao, X.; Chen, H. Characterization of Transcriptional Complexity during Adipose Tissue Development in Bovines of Different Ages and Sexes. PLoS ONE 2014, 9, e101261. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: Generally Applicable Gene Set Enrichment for Pathway Analysis. BMC Bioinform. 2009, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, K.; Peippo, J.; Honkatukia, M.; Seppälä, A.; Rautiainen, J.; Ghanem, N.; Hamama, T.-M.; Crowe, M.A.; Andersson, M.; Li, M.-H.; et al. Integrated Ovarian MRNA and MiRNA Transcriptome Profiling Characterizes the Genetic Basis of Prolificacy Traits in Sheep (Ovis aries). BMC Genom. 2018, 19, 104. [Google Scholar] [CrossRef]
- Pelletier, M.; Kotiaho, A.; Niinimäki, S.; Salmi, A.-K. Identifying Early Stages of Reindeer Domestication in the Archaeological Record: A 3D Morphological Investigation on Forelimb Bones of Modern Populations from Fennoscandia. Archaeol. Anthropol. Sci. 2020, 12, 169. [Google Scholar] [CrossRef]
- Mizuno, M.; Kuboki, Y. Osteoblast-Related Gene Expression of Bone Marrow Cells during the Osteoblastic Differentiation Induced by Type I Collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef]
- Lee, M.-J.; Fried, S.K. Sex-Dependent Depot Differences in Adipose Tissue Development and Function; Role of Sex Steroids. J. Obes. Metab. Syndr. 2017, 26, 172–180. [Google Scholar] [CrossRef]
- Valencak, T.G.; Osterrieder, A.; Schulz, T.J. Sex Matters: The Effects of Biological Sex on Adipose Tissue Biology and Energy Metabolism. Redox Biol. 2017, 12, 806–813. [Google Scholar] [CrossRef]
- Mukaj, A.; Piálek, J.; Fotopulosova, V.; Morgan, A.P.; Odenthal-Hesse, L.; Parvanov, E.D.; Forejt, J. Prdm9 Intersubspecific Interactions in Hybrid Male Sterility of House Mouse. Mol. Biol. Evol. 2020, 37, 3423–3438. [Google Scholar] [CrossRef]
- Davies, B.; Hatton, E.; Altemose, N.; Hussin, J.G.; Pratto, F.; Zhang, G.; Hinch, A.G.; Moralli, D.; Biggs, D.; Diaz, R.; et al. Re-Engineering the Zinc Fingers of PRDM9 Reverses Hybrid Sterility in Mice. Nature 2016, 530, 171–176. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.; Smith, H.E. E1 Ubiquitin-Activating Enzyme UBA-1 Plays Multiple Roles throughout C. Elegans Development. PLoS Genet. 2008, 4, e1000131. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, Y.; Riel, J.M.; Stoytcheva, Z.; Ward, M.A. Two Y Genes Can Replace the Entire Y Chromosome for Assisted Reproduction in the Mouse. Science 2014, 343, 1242544. [Google Scholar] [CrossRef]
- Ruthig, V.A.; Nielsen, T.; Riel, J.M.; Yamauchi, Y.; Ortega, E.A.; Salvador, Q.; Ward, M.A. Testicular Abnormalities in Mice with Y Chromosome Deficiencies. Biol. Reprod. 2017, 96, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Lardone, M.C.; Parodi, D.A.; Valdevenito, R.; Ebensperger, M.; Piottante, A.; Madariaga, M.; Smith, R.; Pommer, R.; Zambrano, N.; Castro, A. Quantification of DDX3Y, RBMY1, DAZ and TSPY MRNAs in Testes of Patients with Severe Impairment of Spermatogenesis. Mol. Hum. Reprod. 2007, 13, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Ramathal, C.; Angulo, B.; Sukhwani, M.; Cui, J.; Durruthy-Durruthy, J.; Fang, F.; Schanes, P.; Turek, P.J.; Orwig, K.E.; Reijo Pera, R. DDX3Y Gene Rescue of a Y Chromosome AZFa Deletion Restores Germ Cell Formation and Transcriptional Programs. Sci. Rep. 2015, 5, 15041. [Google Scholar] [CrossRef]
- Matsumura, T.; Endo, T.; Isotani, A.; Ogawa, M.; Ikawa, M. An Azoospermic Factor Gene, Ddx3y and Its Paralog, Ddx3x Are Dispensable in Germ Cells for Male Fertility. J. Reprod. Dev. 2019, 65, 121–128. [Google Scholar] [CrossRef]
- Luo, M.; Peng, D. ANGPTL8: An Important Regulator in Metabolic Disorders. Front. Endocrinol. 2018, 9, 169. [Google Scholar] [CrossRef]
- Kishi, K.; Uchida, A.; Takase, H.M.; Suzuki, H.; Kurohmaru, M.; Tsunekawa, N.; Kanai-Azuma, M.; Wood, S.A.; Kanai, Y. Spermatogonial Deubiquitinase USP9X Is Essential for Proper Spermatogenesis in Mice. Reproduction 2017, 154, 135–143. [Google Scholar] [CrossRef]
- Grey, C.; Baudat, F.; de Massy, B. PRDM9, a Driver of the Genetic Map. PLoS Genet. 2018, 14, e1007479. [Google Scholar] [CrossRef]
- Mihola, O.; Landa, V.; Pratto, F.; Brick, K.; Kobets, T.; Kusari, F.; Gasic, S.; Smagulova, F.; Grey, C.; Flachs, P.; et al. Rat PRDM9 Shapes Recombination Landscapes, Duration of Meiosis, Gametogenesis, and Age of Fertility. BMC Biol. 2021, 19, 86. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male | Female | |
|---|---|---|
| Metacarpal | 3 | 3 |
| Perirenal | 3 | 3 |
| Prescapular | 3 | 3 |
| Comparison | Total | Upregulated | Downregulated |
|---|---|---|---|
| M-F vs. M-M | 225 | 78 | 147 |
| P-F vs. P-M | 104 | 54 | 50 |
| S-F vs. S-M | 49 | 27 | 22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weldenegodguad, M.; Kantanen, J.; Peippo, J.; Pokharel, K. Differences in Adipose Gene Expression Profiles between Male and Female Even Reindeer (Rangifer tarandus) in Sakha (Yakutia). Genes 2022, 13, 1645. https://doi.org/10.3390/genes13091645
Weldenegodguad M, Kantanen J, Peippo J, Pokharel K. Differences in Adipose Gene Expression Profiles between Male and Female Even Reindeer (Rangifer tarandus) in Sakha (Yakutia). Genes. 2022; 13(9):1645. https://doi.org/10.3390/genes13091645
Chicago/Turabian StyleWeldenegodguad, Melak, Juha Kantanen, Jaana Peippo, and Kisun Pokharel. 2022. "Differences in Adipose Gene Expression Profiles between Male and Female Even Reindeer (Rangifer tarandus) in Sakha (Yakutia)" Genes 13, no. 9: 1645. https://doi.org/10.3390/genes13091645