
MiR-18a-5p Targets Connective Tissue Growth Factor Expression and Inhibits Transforming Growth Factor β2-Induced Trabecular Meshwork Cell Contractility


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Donor Anterior Segment Collection
2.2. Isolation, Culture, and Characterisation of Primary Human Trabecular Meshwork Cells
2.3. In Silico miRNA Prediction and Validation
2.4. Treatment of Primary Human TM Cells with TGFβ2 for miRNA Expression Measurement
2.5. miRNA and mRNA RT-qPCR
2.6. miRNA and siRNA Transfection
2.7. shRNA Transduction, Selection and Experimentation
2.8. Collagen Gel Contraction Assay
2.9. Statistical Analysis
3. Results
3.1. In Silico Identification of CTGF-Targeting miRNAs
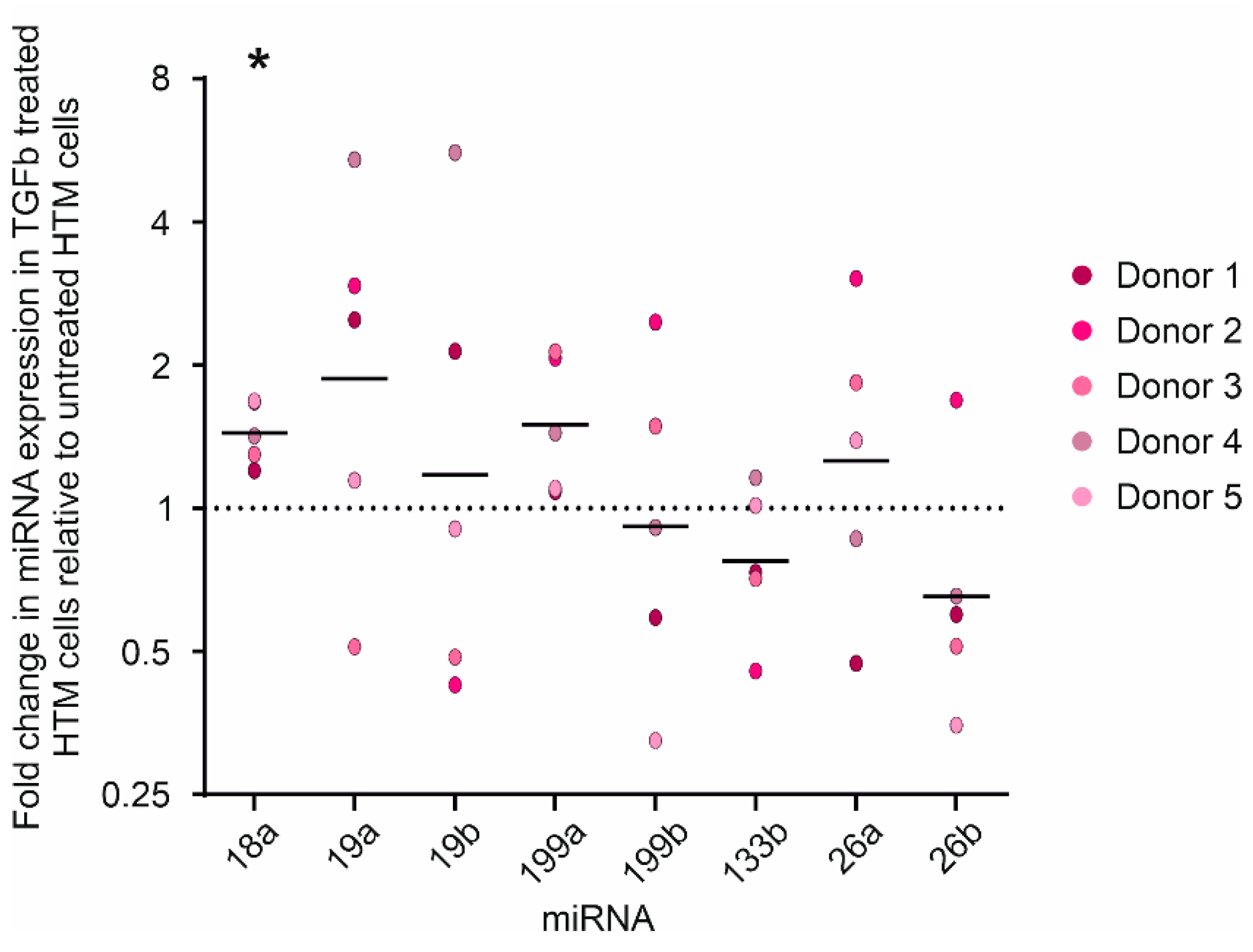
3.2. Expression of miR-18a-5p Increases in Primary Human TM Cells after TGFβ2 Treatment
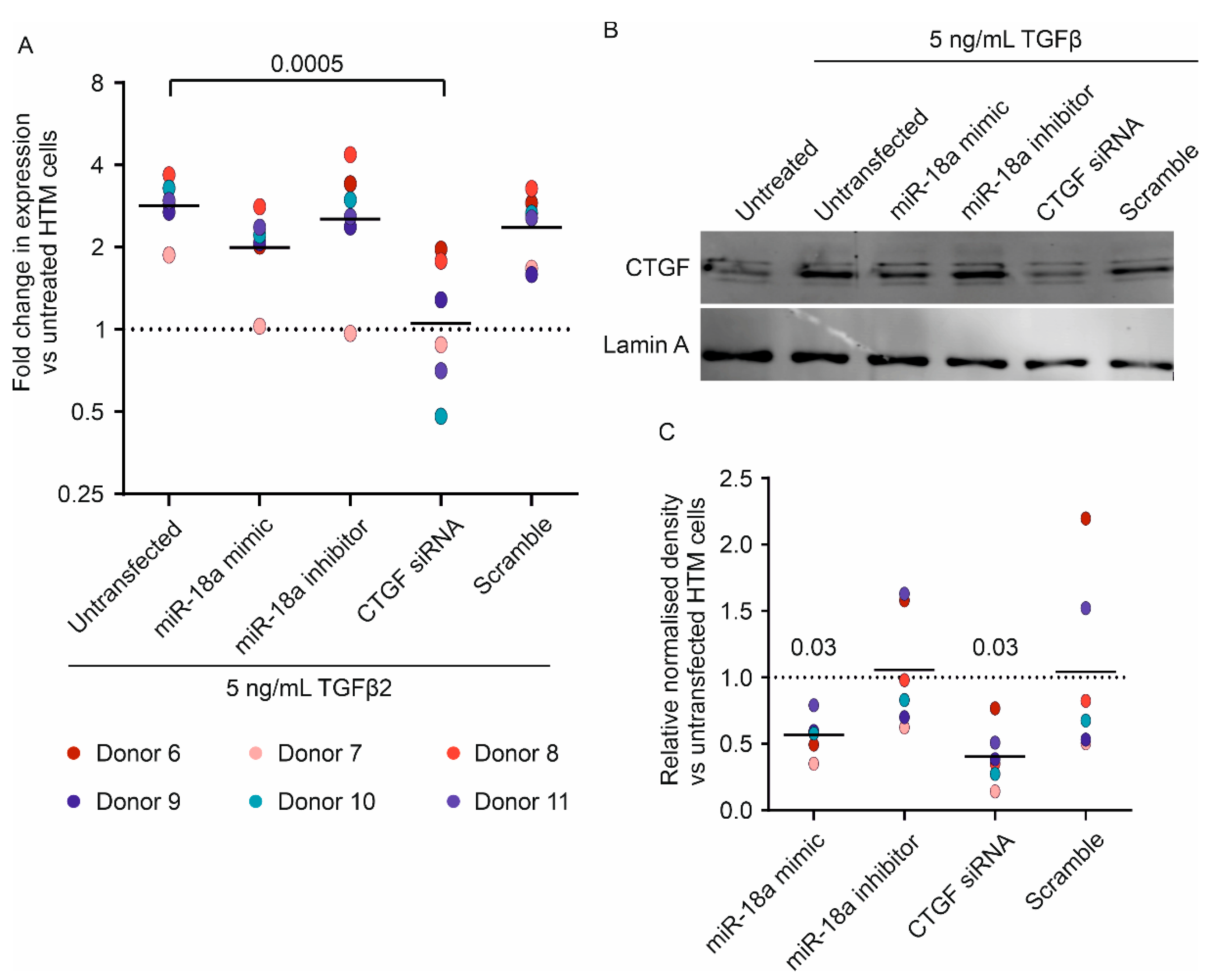
3.3. miR-18a-5p Mimic Reduces TGFβ2-Induced CTGF mRNA and Protein Upregulation in Human TM Cells
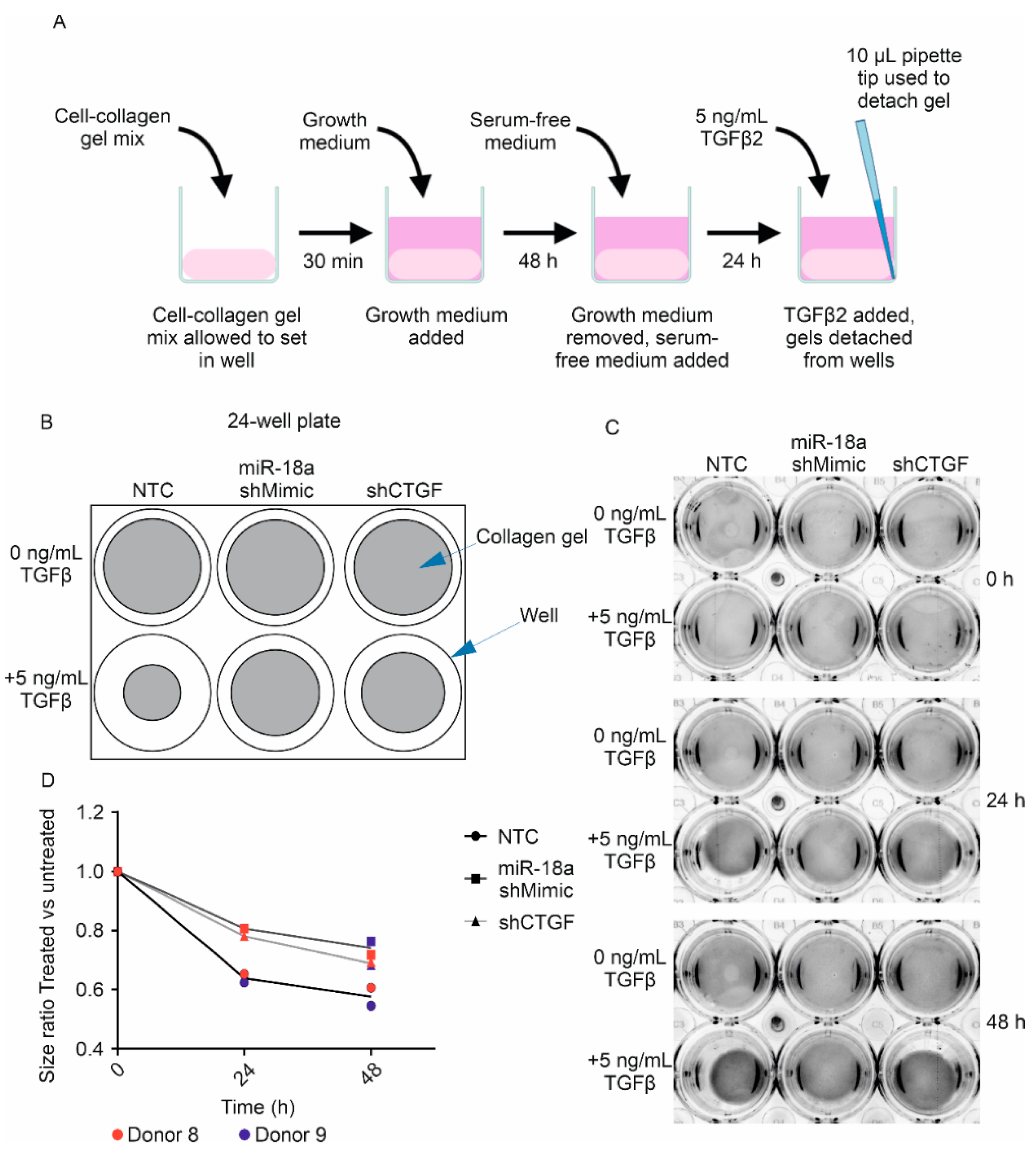
3.4. Lentiviral-Driven Overexpression of miR-18a-5p Leads to Reduced Human TM Cell Contractility in Response to TGFβ2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourne, R.R.A.; Steinmetz, J.D.; Flaxman, S.; Briant, P.S.; Taylor, H.R.; Resnikoff, S.; Casson, R.J.; Abdoli, A.; Abu-Gharbieh, E.; Afshin, A.; et al. Trends in prevalence of blindness and distance and near vision impairment over 30 years: An analysis for the Global Burden of Disease Study. Lancet Glob. Health 2021, 9, e130–e143. [Google Scholar] [CrossRef]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Alqawlaq, S.; Flanagan, J.G.; Sivak, J.M. All roads lead to glaucoma: Induced retinal injury cascades contribute to a common neurodegenerative outcome. Exp. Eye Res. 2019, 183, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Wormald, R.; Virgili, G.; Azuara-Blanco, A. Systematic reviews and randomised controlled trials on open angle glaucoma. Eye 2019, 34, 161–167. [Google Scholar] [CrossRef]
- Gottanka, J.; Johnson, D.H.; Martus, P.; Lütjen-Drecoll, E. Severity of optic nerve damage in eyes with POAG is correlated with changes in the trabecular meshwork. J. Glaucoma 1997, 6, 123–132. [Google Scholar] [CrossRef]
- Lepple-Wienhues, A.; Stahl, F.; Wiederholt, M. Differential smooth muscle-like contractile properties of trabecular meshwork and ciliary muscle. Exp. Eye Res. 1991, 53, 33–38. [Google Scholar] [CrossRef]
- Wiederholt, M. Direct involvement of trabecular meshwork in the regulation of aqueous humor outflow. Curr. Opin. Ophthalmol. 1998, 9, 46–49. [Google Scholar] [CrossRef]
- Tian, B.; Gabelt, B.T.; Geiger, B.; Kaufman, P.L. The role of the actomyosin system in regulating trabecular fluid outflow. Exp. Eye Res. 2009, 88, 713–717. [Google Scholar] [CrossRef] [Green Version]
- Fuchshofer, R.; Tamm, E.R. The role of TGF-β in the pathogenesis of primary open-angle glaucoma. Cell Tissue Res. 2012, 347, 279–290. [Google Scholar] [CrossRef]
- Keller, K.E.; Aga, M.; Bradley, J.M.; Kelley, M.J.; Acott, T.S. Extracellular matrix turnover and outflow resistance. Exp. Eye Res. 2009, 88, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Wordinger, R.J.; Sharma, T.; Clark, A.F. The role of TGF-β2 and bone morphogenetic proteins in the trabecular meshwork and glaucoma. J. Ocul. Pharmacol. Ther. 2014, 30, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Stamer, W.D.; Acott, T.S. Current Understanding of Conventional Outflow Dysfunction in Glaucoma. Curr. Opin. Ophthalmol. 2012, 23, 135. [Google Scholar] [CrossRef] [Green Version]
- Prendes, M.A.; Harris, A.; Wirostko, B.M.; Gerber, A.L.; Siesky, B. The role of transforming growth factor β in glaucoma and the therapeutic implications. Br. J. Ophthalmol. 2013, 97, 680–686. [Google Scholar] [CrossRef]
- Junglas, B.; Kuespert, S.; Seleem, A.A.; Struller, T.; Ullmann, S.; Bösl, M.; Bosserhoff, A.; Köstler, J.; Wagner, R.; Tamm, E.R.; et al. Connective tissue growth factor causes glaucoma by modifying the actin cytoskeleton of the trabecular meshwork. Am. J. Pathol. 2012, 180, 2386–2403. [Google Scholar] [CrossRef]
- Browne, J.G.; Ho, S.L.; Kane, R.; Oliver, N.; Clark, A.F.; O’Brien, C.J.; Crean, J.K. Connective tissue growth factor is increased in pseudoexfoliation glaucoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 3660–3666. [Google Scholar] [CrossRef] [Green Version]
- Wallace, D.M.; Murphy-Ullrich, J.E.; Downs, J.C.; O’Brien, C.J. The role of matricellular proteins in glaucoma. Matrix Biol. 2014, 37, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Junglas, B.; Yu, A.H.L.; Welge-Lüssen, U.; Tamm, E.R.; Fuchshofer, R. Connective tissue growth factor induces extracellular matrix deposition in human trabecular meshwork cells. Exp. Eye Res. 2009, 88, 1065–1075. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, N.; Chu, H.Y.; Yu, Y.; Zhang, Z.K.; Zhang, G.; Zhang, B.T. Connective Tissue Growth Factor: From Molecular Understandings to Drug Discovery. Front. Cell Dev. Biol. 2020, 8, 1239. [Google Scholar] [CrossRef]
- Lee, E.J.; Han, J.C.; Park, D.Y.; Cho, J.; Kee, C. Effect of connective tissue growth factor gene editing using adeno-associated virus-mediated CRISPR-Cas9 on rabbit glaucoma filtering surgery outcomes. Gene Ther. 2021, 28, 277–286. [Google Scholar] [CrossRef]
- Wallace, D.M.; Clark, A.F.; Lipson, K.E.; Andrews, D.; Crean, J.K.; O’Brien, C.J. Anti-connective tissue growth factor antibody treatment reduces Extracellular matrix production in Trabecular meshwork and Lamina Cribrosa cells. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7836–7848. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Bilir, E.K.; Kingston, O.A.; Oldershaw, R.A.; Kearns, V.R.; Willoughby, C.E.; Sheridan, C.M. Replacement of the Trabecular Meshwork Cells-A Way Ahead in IOP Control? Biomolecules 2021, 11, 1371. [Google Scholar] [CrossRef]
- Huntzinger, E.; Izaurralde, E. Gene silencing by microRNAs: Contributions of translational repression and mRNA decay. Nat. Rev. Genet. 2011, 12, 99–110. [Google Scholar] [CrossRef]
- Kehl, T.; Backes, C.; Kern, F.; Fehlmann, T.; Ludwig, N.; Meese, E.; Lenhof, H.P.; Keller, A. About miRNAs, miRNA seeds, target genes and target pathways. Oncotarget 2017, 8, 107167–107175. [Google Scholar] [CrossRef] [Green Version]
- Kern, F.; Krammes, L.; Danz, K.; Diener, C.; Kehl, T.; Küchler, O.; Fehlmann, T.; Kahraman, M.; Rheinheimer, S.; Aparicio-Puerta, E.; et al. Validation of human microRNA target pathways enables evaluation of target prediction tools. Nucleic Acids Res. 2021, 49, 127. [Google Scholar] [CrossRef]
- Kehl, T.; Kern, F.; Backes, C.; Fehlmann, T.; Stöckel, D.; Meese, E.; Lenhof, H.P.; Keller, A. MiRPathDB 2.0: A novel release of the miRNA Pathway Dictionary Database. Nucleic Acids Res. 2020, 48, D142–D147. [Google Scholar] [CrossRef]
- Chen, Y.C.; Chen, B.C.; Yu, C.C.; Lin, S.H.; Lin, C.H. miR-19a, -19b, and -26b Mediate CTGF Expression and Pulmonary Fibroblast Differentiation. J. Cell. Physiol. 2016, 231, 2236–2248. [Google Scholar] [CrossRef]
- Koga, K.; Yokoi, H.; Mori, K.; Kasahara, M.; Kuwabara, T.; Imamaki, H.; Ishii, A.; Mori, K.P.; Kato, Y.; Ohno, S.; et al. MicroRNA-26a inhibits TGF-β-induced extracellular matrix protein expression in podocytes by targeting CTGF and is downregulated in diabetic nephropathy. Diabetologia 2015, 58, 2169–2180. [Google Scholar] [CrossRef] [Green Version]
- Duisters, R.F.; Tijsen, A.J.; Schroen, B.; Leenders, J.J.; Lentink, V.; van der Made, I.; Herias, V.; van Leeuwen, R.E.; Schellings, M.W.; Barenbrug, P.; et al. miR-133 and miR-30 regulate connective tissue growth factor: Implications for a role of microRNAs in myocardial matrix remodeling. Circ. Res. 2009, 104, 170–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Li, X.; Lin, C.; Zhang, Y.; Hu, G.; Zhou, J.; Du, J.; Gao, K.; Gan, Y.; Deng, H. MicroRNA-133b inhibits connective tissue growth factor in colorectal cancer and correlates with the clinical stage of the disease. Mol. Med. Rep. 2015, 11, 2805–2812. [Google Scholar] [CrossRef] [PubMed]
- Lun, W.; Wu, X.; Deng, Q.; Zhi, F. MiR-218 regulates epithelial-mesenchymal transition and angiogenesis in colorectal cancer via targeting CTGF. Cancer Cell Int. 2018, 18, 83. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Han, S.; Liu, C.; Zhou, R.; Sun, W.; Zhang, Z.; Qu, J. Microrna-199a-5p functions as a tumor suppressor via suppressing connective tissue growth factor (CTGF) in follicular thyroid carcinoma. Med. Sci. Monit. 2016, 22, 1210–1217. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Bier, A.; Cazacu, S.; Finniss, S.; Xiang, C.; Twito, H.; Poisson, L.M.; Mikkelsen, T.; Slavin, S.; Jacoby, E.; et al. MicroRNA-145 Is Downregulated in Glial Tumors and Regulates Glioma Cell Migration by Targeting Connective Tissue Growth Factor. PLoS ONE 2013, 8, e54652. [Google Scholar] [CrossRef] [Green Version]
- Xie, H.; Zhao, Y.; Caramuta, S.; Larsson, C.; Lui, W.-O. miR-205 Expression Promotes Cell Proliferation and Migration of Human Cervical Cancer Cells. PLoS ONE 2012, 7, e46990. [Google Scholar] [CrossRef]
- Mu, S.; Kang, B.; Zeng, W.; Sun, Y.; Yang, F. MicroRNA-143-3p inhibits hyperplastic scar formation by targeting connective tissue growth factor CTGF/CCN2 via the Akt/mTOR pathway. Mol. Cell. Biochem. 2016, 416, 99–108. [Google Scholar] [CrossRef]
- Guo, Y.; Lu, X.; Wang, H. Downregulation of miR-18a induces CTGF and promotes proliferation and migration of sodium hyaluronate treated human corneal epithelial cells. Gene 2016, 592, 129–136. [Google Scholar] [CrossRef]
- van Almen, G.C.; Verhesen, W.; van Leeuwen, R.E.W.; van de Vrie, M.; Eurlings, C.; Schellings, M.W.M.; Swinnen, M.; Cleutjens, J.P.M.; van Zandvoort, M.A.M.J.; Heymans, S.; et al. MicroRNA-18 and microRNA-19 regulate CTGF and TSP-1 expression in age-related heart failure. Aging Cell 2011, 10, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Wecker, T.; Hoffmeier, K.; Plötner, A.; Grüning, B.A.; Horres, R.; Backofen, R.; Reinhard, T.; Schlunck, G. MicroRNA profiling in aqueous humor of individual human eyes by next-generation sequencing. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1706–1713. [Google Scholar] [CrossRef]
- Drewry, M.; Helwa, I.; Allingham, R.R.; Hauser, M.A.; Liu, Y. miRNA profile in three different normal human ocular tissues by miRNA-seq. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3731–3739. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Luna, C.; Qiu, J.; Epstein, D.L.; Gonzalez, P. Alterations in microRNA expression in stress-induced cellular senescence. Mech. Ageing Dev. 2009, 130, 731–741. [Google Scholar] [CrossRef] [Green Version]
- Butz, H.; Rácz, K.; Hunyady, L.; Patócs, A. Crosstalk between TGF-β signaling and the microRNA machinery. Trends Pharm. Sci. 2012, 33, 382–393. [Google Scholar] [CrossRef]
- Keller, K.E.; Bhattacharya, S.K.; Borrás, T.; Brunner, T.M.; Chansangpetch, S.; Clark, A.F.; Dismuke, W.M.; Du, Y.; Elliott, M.H.; Ethier, C.R.; et al. Consensus recommendations for trabecular meshwork cell isolation, characterization and culture. Exp. Eye Res. 2018, 171, 164–173. [Google Scholar] [CrossRef]
- Callaghan, B.; Lester, K.; Lane, B.; Fan, X.; Goljanek-Whysall, K.; Simpson, D.A.; Sheridan, C.; Willoughby, C.E. Genome-wide transcriptome profiling of human trabecular meshwork cells treated with TGF-β2. Sci. Rep. 2022, 12, 9564. [Google Scholar] [CrossRef]
- Kathirvel, K.; Karen, L.; Haribalaganesh, R.; Krishnadas, R.; Muthukkaruppan, V.; Lane, B.; Simpson, D.A.; Goljanek-Whysall, K.; Sheridan, C.; Bharanidharan, D.; et al. Short and long-term effect of dexamethasone on the transcriptome profile of primary human trabecular meshwork cells in vitro. Sci. Rep. 2022, 12, 8299. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. Elife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. MiRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wang, X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. 2019, 20, 18. [Google Scholar] [CrossRef] [PubMed]
- Reczko, M.; Maragkakis, M.; Alexiou, P.; Grosse, I.; Hatzigeorgiou, A.G. Functional microRNA targets in protein coding sequences. Bioinformatics 2012, 28, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.S.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, A.J.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.S. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [Green Version]
- John, B.; Enright, A.J.; Aravin, A.; Tuschl, T.; Sander, C.; Marks, D.S. Human microRNA targets. PLoS Biol. 2004, 2, e363. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.H.; Shrestha, S.; Yang, C.D.; Chang, N.W.; Lin, Y.L.; Liao, K.W.; Huang, W.C.; Sun, T.H.; Tu, S.J.; Lee, W.H.; et al. MiRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Duan, L.J.; Qi, J.; Kong, X.J.; Huang, T.; Qian, X.Q.; Xu, D.; Liang, J.H.; Kang, J. MiR-133 modulates TGF-β1-induced bladder smooth muscle cell hypertrophic and fibrotic response: Implication for a role of microRNA in bladder wall remodeling caused by bladder outlet obstruction. Cell. Signal. 2015, 27, 215–227. [Google Scholar] [CrossRef]
- Lin, C.-H.H.; Yu, M.-C.C.; Tung, W.-H.H.; Chen, T.-T.T.; Yu, C.-C.C.; Weng, C.-M.M.; Tsai, Y.-J.J.; Bai, K.-J.J.; Hong, C.-Y.Y.; Chien, M.-H.H.; et al. Connective tissue growth factor induces collagen I expression in human lung fibroblasts through the Rac1/MLK3/JNK/AP-1 pathway. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 2823–2833. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Harris, A.; Prendes, M.A.; Alshawa, L.; Gross, J.C.; Wentz, S.M.; Rao, A.B.; Kim, N.J.; Synder, A.; Siesky, B. Targeting Transforming Growth Factor-β Signaling in Primary Open-Angle Glaucoma. J. Glaucoma 2017, 26, 390–395. [Google Scholar] [CrossRef]
- Mietzner, R.; Breunig, M. Causative glaucoma treatment: Promising targets and delivery systems. Drug Discov. Today 2019, 24, 1606–1613. [Google Scholar] [CrossRef]
- Tomarev, S.I.; Wistow, G.; Raymond, V.; Dubois, S.; Malyukova, I. Gene expression profile of the human trabecular meshwork: NEIBank sequence tag analysis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2588–2596. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.V.; Pattabiraman, P.P.; Kopczynski, C. Role of the Rho GTPase/Rho kinase signaling pathway in pathogenesis and treatment of glaucoma: Bench to bedside research. Exp. Eye Res. 2017, 158, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.K.; Chang, R.T. An emerging treatment option for glaucoma: Rho kinase inhibitors. Clin. Ophthalmol. 2014, 8, 883. [Google Scholar] [CrossRef] [Green Version]
- Dallon, J.C.; Ehrlich, H.P. A review of fibroblast-populated collagen lattices. Wound Repair Regen. 2008, 16, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Pattabiraman, P.P.; Maddala, R.; Rao, P.V. Regulation of plasticity and fibrogenic activity of trabecular meshwork cells by rho GTPase signaling. J. Cell. Physiol. 2014, 229, 927–942. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ye, H.; Xiang, F.; Song, L.J.; Zhou, L.L.; Cai, P.C.; Zhang, J.C.; Yu, F.; Shi, H.Z.; Su, Y.; et al. miR-18a-5p Inhibits Sub-pleural Pulmonary Fibrosis by Targeting TGF-β Receptor II. Mol. Ther. 2017, 25, 728–738. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Shi, J.Y.; Zhu, G.Q.; Shi, B. MiR-17-92 cluster regulates cell proliferation and collagen synthesis by targeting TGFB pathway in mouse palatal mesenchymal cells. J. Cell. Biochem. 2012, 113, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, T.; Guglas, K.; Kopczyńska, M.; Sobocińska, J.; Teresiak, A.; Bliźniak, R.; Lamperska, K. Good or not good: Role of miR-18a in cancer biology. Reports Pract. Oncol. Radiother. 2020, 25, 808. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, R.P.; Leonard, M.O.; Murphy, M.; Clark, A.F.; O’Brien, C.J. Transforming growth factor-beta-regulated gene transcription and protein expression in human GFAP-negative lamina cribrosa cells. Glia 2005, 52, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Dillinger, A.E.; Weber, G.R.; Mayer, M.; Schneider, M.; Göppner, C.; Ohlmann, A.; Shamonin, M.; Monkman, G.J.; Fuchshofer, R. CCN2/CTGF-A Modulator of the Optic Nerve Head Astrocyte. Front. Cell Dev. Biol. 2022, 10, 864433. [Google Scholar] [CrossRef]
- Mahale, A.; Othman, M.W.; Al Shahwan, S.; Al Jadaan, I.; Owaydha, O.; Khan, Z.; Edward, D.P. Altered expression of fibrosis genes in capsules of failed Ahmed glaucoma valve implants. PLoS ONE 2015, 10, e0122409. [Google Scholar] [CrossRef] [Green Version]
- Esson, D.W.; Neelakantan, A.; Iyer, S.A.; Blalock, T.D.; Balasubramanian, L.; Grotendorst, G.R.; Schultz, G.S.; Sherwood, M.B. Expression of connective tissue growth factor after glaucoma filtration surgery in a rabbit model. Investig. Ophthalmol. Vis. Sci. 2004, 45, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Klaassen, I.; van Geest, R.J.; Kuiper, E.J.; van Noorden, C.J.F.; Schlingemann, R.O. The role of CTGF in diabetic retinopathy. Exp. Eye Res. 2015, 133, 37–48. [Google Scholar] [CrossRef]
- Kothary, P.C.; Badhwar, J.; Weng, C.; Del Monte, M.A. Impaired Intracellular Signaling May Allow Up-Regulation of CTGF-Synthesis and Secondary Peri-Retinal Fibrosis in Human Retinal Pigment Epithelial Cells from Patients with Age-Related Macular Degeneration; Springer: New York, NY, USA, 2010; pp. 419–428. [Google Scholar]
- He, S.; Chen, Y.; Khankan, R.; Barron, E.; Burton, R.; Zhu, D.H.; Ryan, S.J.; Oliver, N.; Hinton, D.R. Connective tissue growth factor as a mediator of intraocular fibrosis. Investig. Ophthalmol. Vis. Sci. 2008, 49, 4078–4088. [Google Scholar] [CrossRef]
- Greene, K.M.; Stamer, W.D.; Liu, Y. The role of microRNAs in glaucoma. Exp. Eye Res. 2022, 215, 108909. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Synthetic RNA | Sequence | Qiagen ID |
|---|---|---|
| hsa-miR-18a-5p mimic | 5′-UAAGGUGCAUCUAGUGCAGAUAG | MSY0000072 |
| hsa-miR-18a-5p inhibitor | 5′-CUAUCUGCACUAGAUGCACCUUA | MIN0000072 |
| FlexiTube CTGF siRNA | 5′-UAUCUGAUGAUACUAACCUTT | SI00029680 |
| Scrambled negative control | Proprietary; no known homology to any mammalian gene | 1027280 |
| miRNA | TargetScan | miRDB TS | miRANDA-mirSVR | MicroT-CDS miTG Score | miRTarBase Validation | Literature | TM/AH Expression |
|---|---|---|---|---|---|---|---|
| miR-124-3p | WCS: −0.38 PCT: 0.22 | 74 | N/A | 0.66 | RA, microA | ||
| miR-1297 | WCS: −0.29 PCT: 0.69 | 65 | SVR: −0.1096 PC: 0.7355 | 0.96 | Not validated | ||
| miR-132-3p | WCS: −0.28 PCT: 0.26 | 93 | SVR: −1.2409 PC: 0.7586 | 0.68 | Not validated | TM [38] | |
| miR-133a-3p | WCS: −0.31 PCT: 0.8 | 56 | N/A | 0.91 | Not validated | [54] | AH [37] TM [38] |
| miR-133b | WCS: −0.31 PCT: 0.8 | 93 | SVR: −0.2487 PC: 0.7793 | 0.91 | RA, WB, qPCR | [28,29,54] | AH [37] |
| miR-143-3p | WCS: −0.24 PCT: 0.26 | 76 | SVR: −0.2242 PC: 0.7355 | 0.84 | RA | [34] | AH [37] TM [38] |
| miR-18a-5p | WCS: −0.37 PCT: 0.45 | 86 | SVR: −1.0860 PC: 0.7793 | 0.91 | RA, WB, microA | [36] | TM [39] |
| miR-18b-5p | WCS: −0.37 PCT: 0.45 | 86 | SVR: −1.0860 PC: 0.7793 | 0.91 | RA, WB | [55] | |
| miR-199a-5p | WCS: −0.19 PCT: 0.24 | 63 | SVR: −0.4603 PC: 0.6405 | 0.65 | Not validated | [31] | AH [37] TM [38] |
| miR-199b-5p | WCS: −0.2 PCT: 0.24 | 63 | SVR: −0.4603 PC: 0.6405 | 0.65 | Not validated | AH [37] TM [38,39] | |
| miR-19a-3p | WCS: −0.26 PCT: 0.82 | No score | SVR: −0.7564 PC: 0.7793 | 0.75 | Other † | [26] | AH [37] TM [38] |
| miR-19b-3p | WCS: −0.27 PCT: 0.82 | No score | SVR: −0.7278 PC: 0.7793 | 0.76 | Other † | [26,36] | AH [37] TM [38] |
| miR-212-3p | WCS: −0.3 PCT: 0.26 | 93 | SVR: −1.2409 PC: 0.7586 | 0.70 | Not validated | ||
| miR-26a-5p | WCS: −0.31 PCT: 0.69 | 57 | SVR: −0.1085 PC: 0.7355 | 0.95 | WB, microA | [27] | AH [37] TM [38] |
| miR-26b-5p | WCS: −0.29 PCT: 0.69 | 57 | SVR: −0.1085 PC: 0.7355 | 0.95 | RA, WB, qPCR | [26] | AH [37] TM [38] |
| miR-4735-3p | WCS: −0.36 PCT: 0.45 | 86 | N/A | 0.89 | Not validated | ||
| miR-4770 | WCS: −0.24 PCT: 0.26 | 76 | N/A | 0.83 | Not validated | ||
| miR-6088 | WCS: −0.24 PCT: 0.26 | 72 | N/A | 0.62 | Not validated |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knox, J.; Bou-Gharios, G.; Hamill, K.J.; Willoughby, C.E. MiR-18a-5p Targets Connective Tissue Growth Factor Expression and Inhibits Transforming Growth Factor β2-Induced Trabecular Meshwork Cell Contractility. Genes 2022, 13, 1500. https://doi.org/10.3390/genes13081500
Knox J, Bou-Gharios G, Hamill KJ, Willoughby CE. MiR-18a-5p Targets Connective Tissue Growth Factor Expression and Inhibits Transforming Growth Factor β2-Induced Trabecular Meshwork Cell Contractility. Genes. 2022; 13(8):1500. https://doi.org/10.3390/genes13081500
Chicago/Turabian StyleKnox, John, George Bou-Gharios, Kevin J. Hamill, and Colin E. Willoughby. 2022. "MiR-18a-5p Targets Connective Tissue Growth Factor Expression and Inhibits Transforming Growth Factor β2-Induced Trabecular Meshwork Cell Contractility" Genes 13, no. 8: 1500. https://doi.org/10.3390/genes13081500