
Identification of Alkaline Salt Tolerance Genes in Brassica napus L. by Transcriptome Analysis

 , and
, and 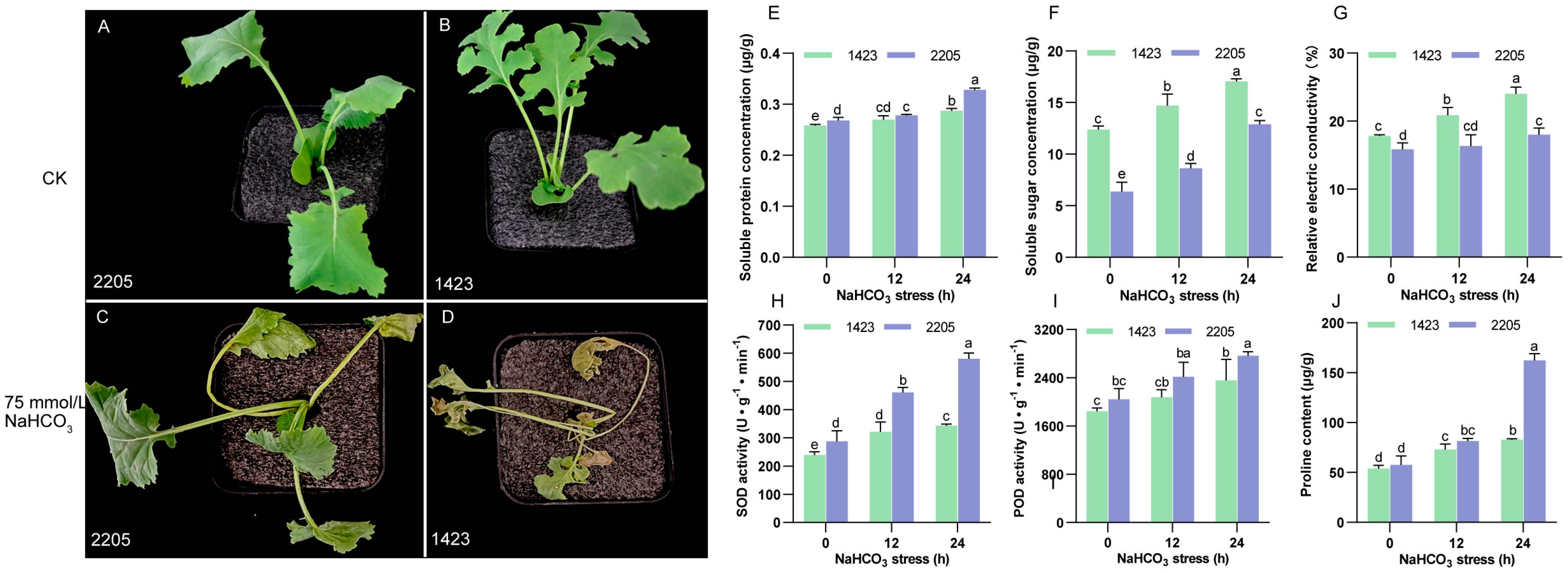{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Measurement of Physiological Indexes
2.3. Electron Microscopic Observation of Seedling Cells under NaHCO3 Stress
2.4. RNA Extraction, Library Construction, and Sequencing
2.5. RNA-Seq Analysis
2.6. Functional Enrichment Analysis of DEGs
2.7. Verified DEGs by qRT-PCR
2.8. Statistical Analysis
3. Results
3.1. Differences of Alkaline Salt Tolerance and Physiological Indicators between Lines 2205 and 1423 under Alkaline Salt Stress
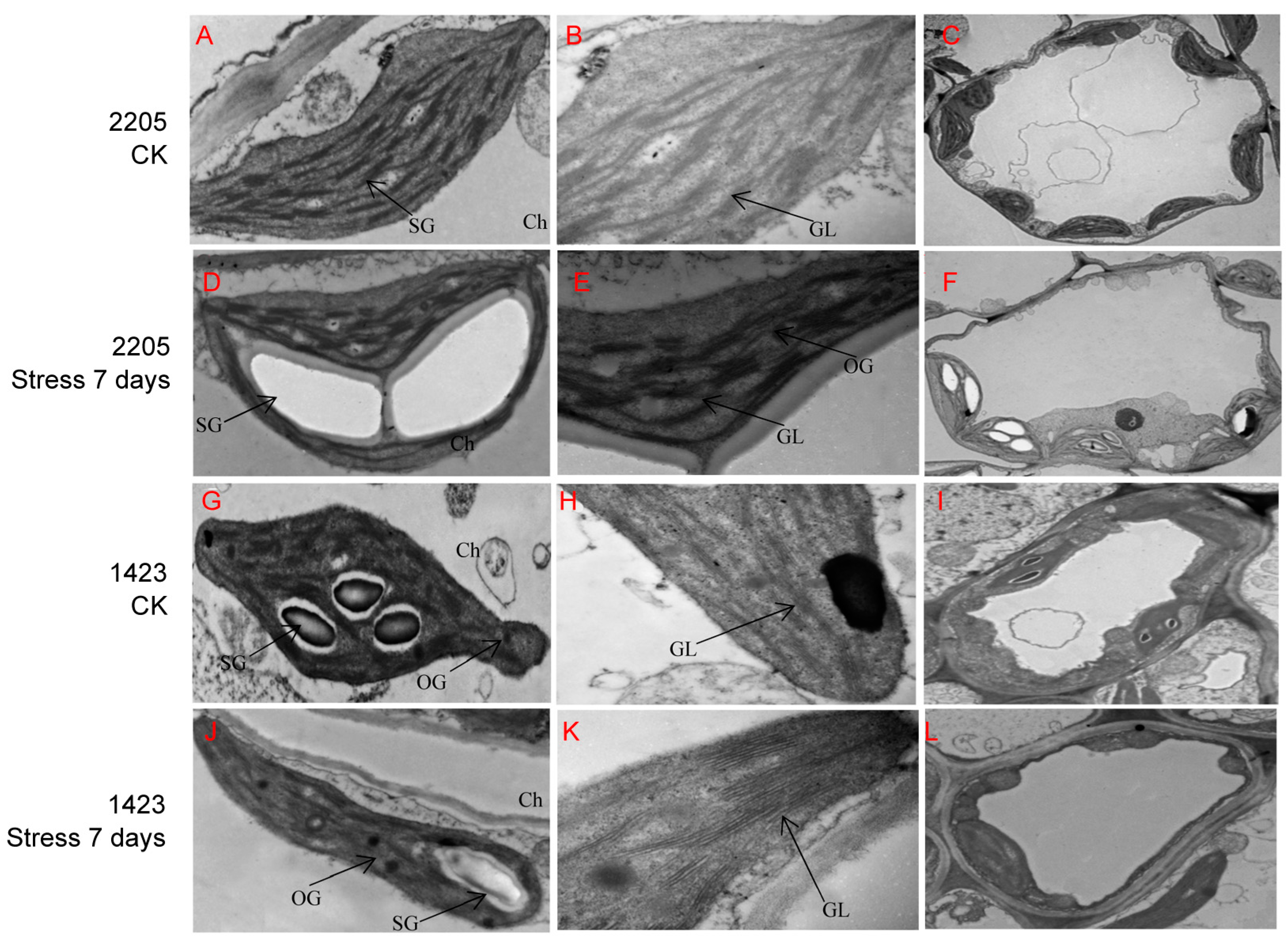
3.2. Ultrastructural-Feature Differences between Line 2205 and Line 1423 under Alkaline Salt Stress
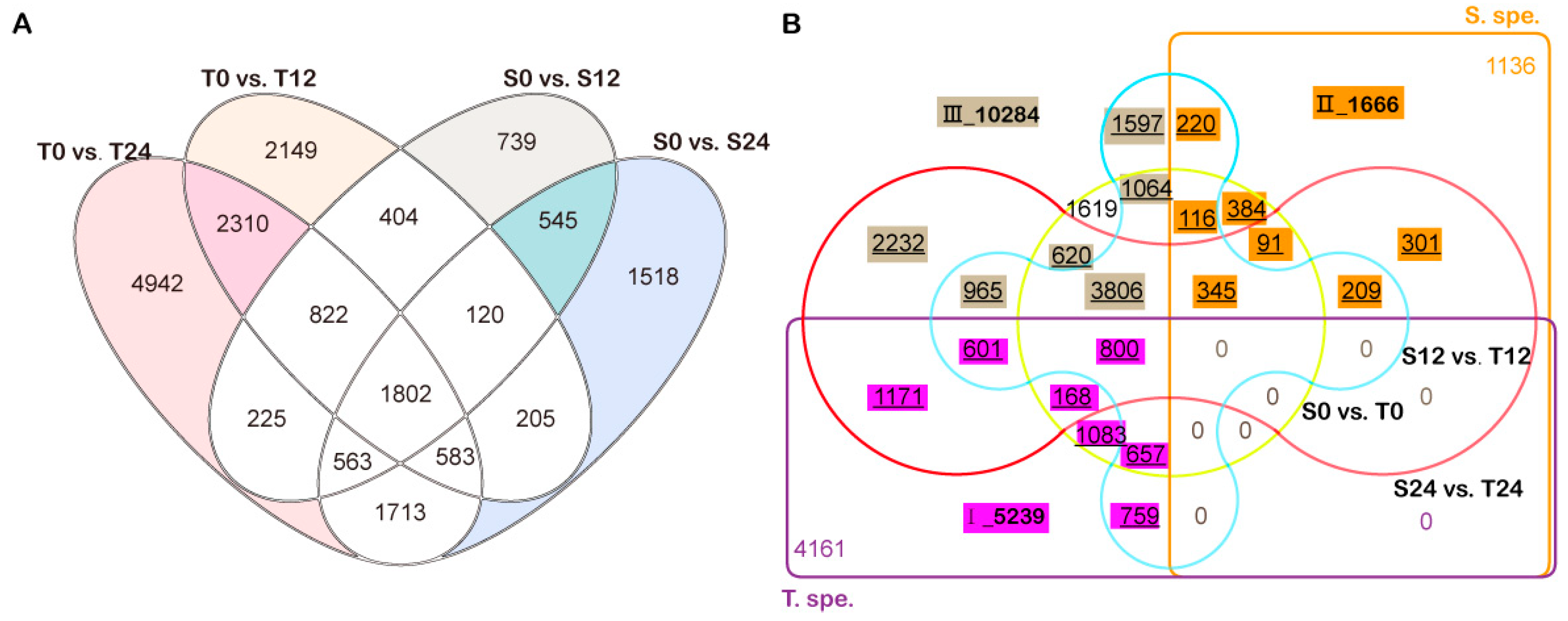
3.3. Illumina Sequencing and Differentially Expressed Genes between Line 2205 and Line 1423
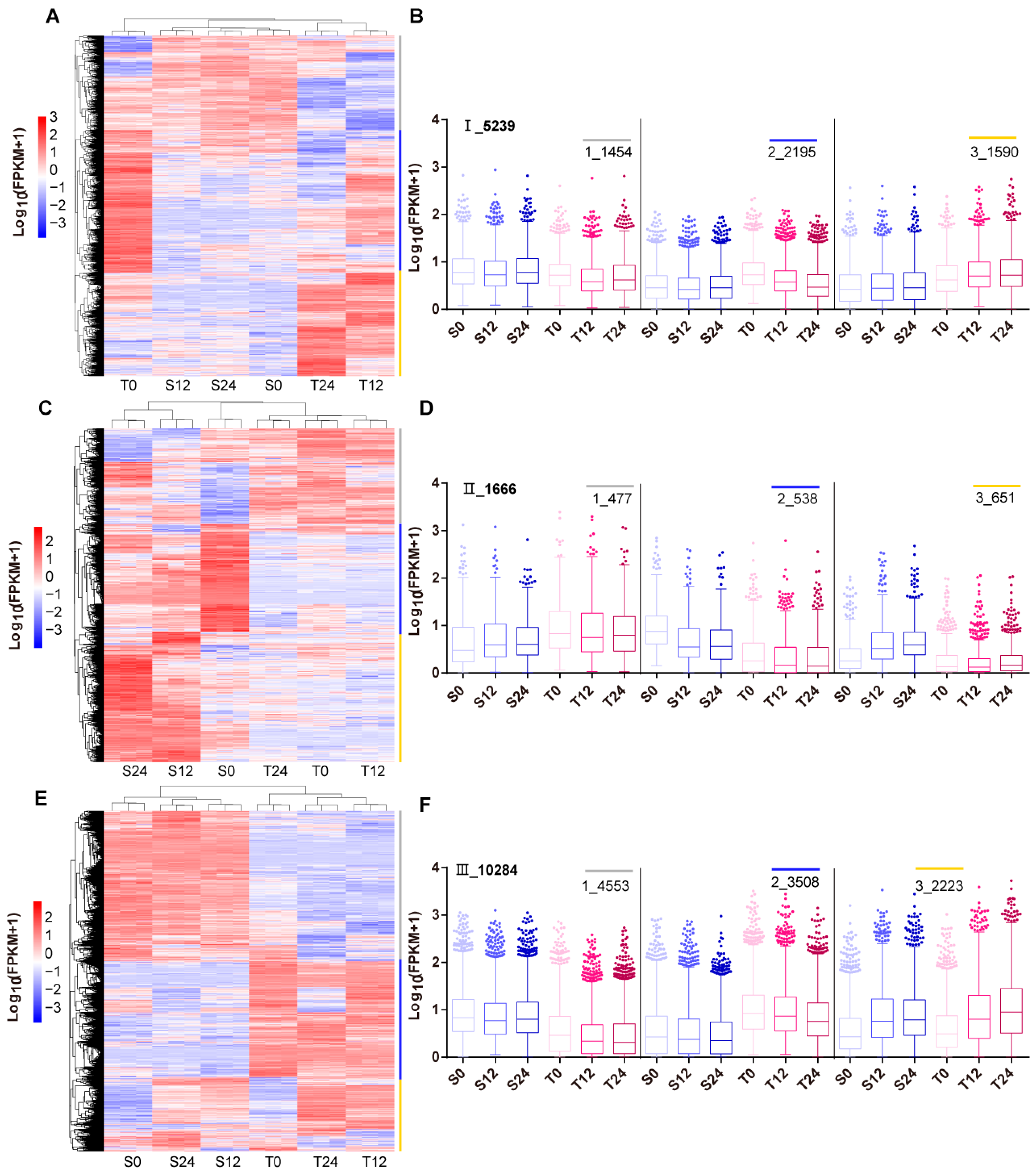
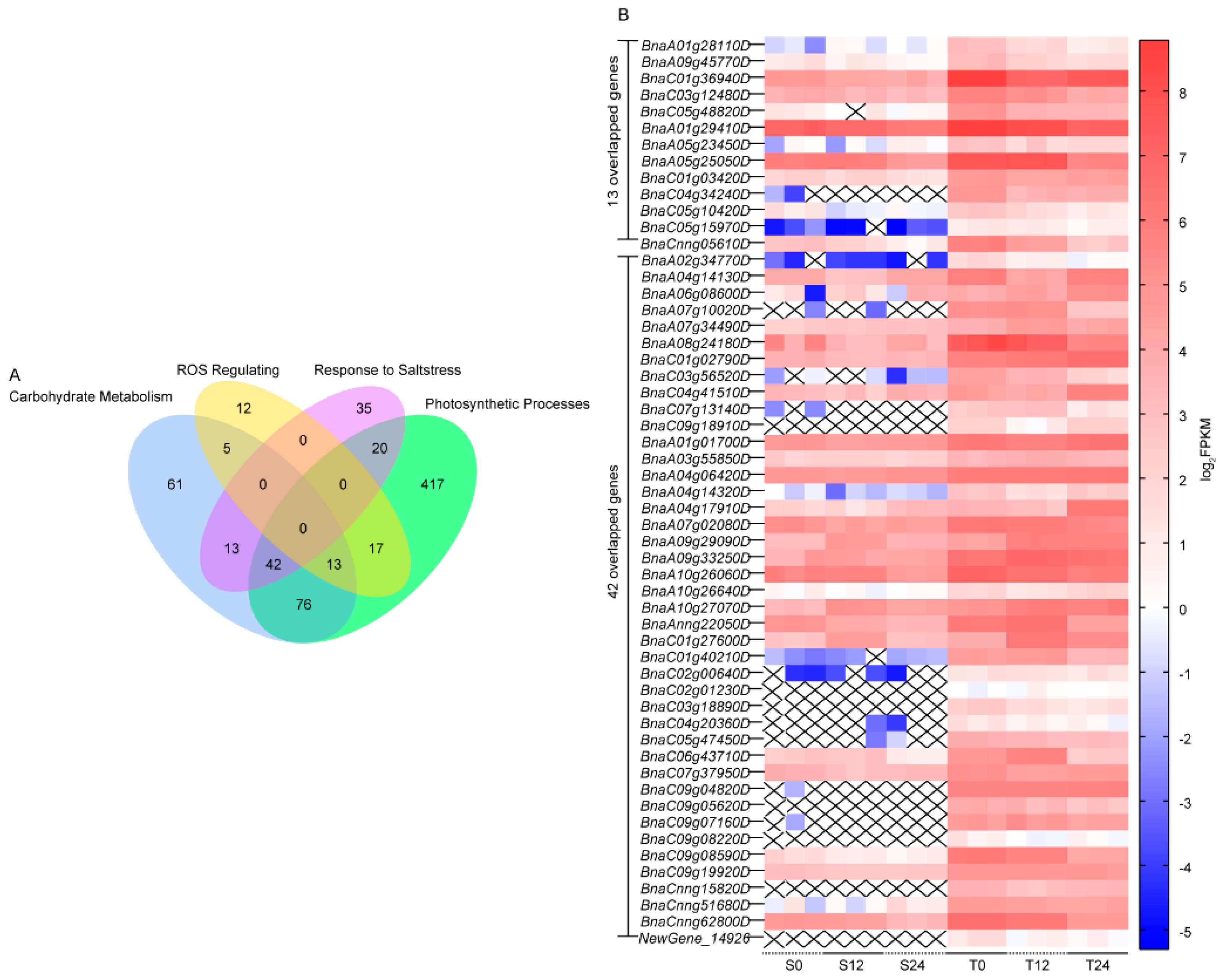
3.4. Co-expression Clustering of Filtered DEGs
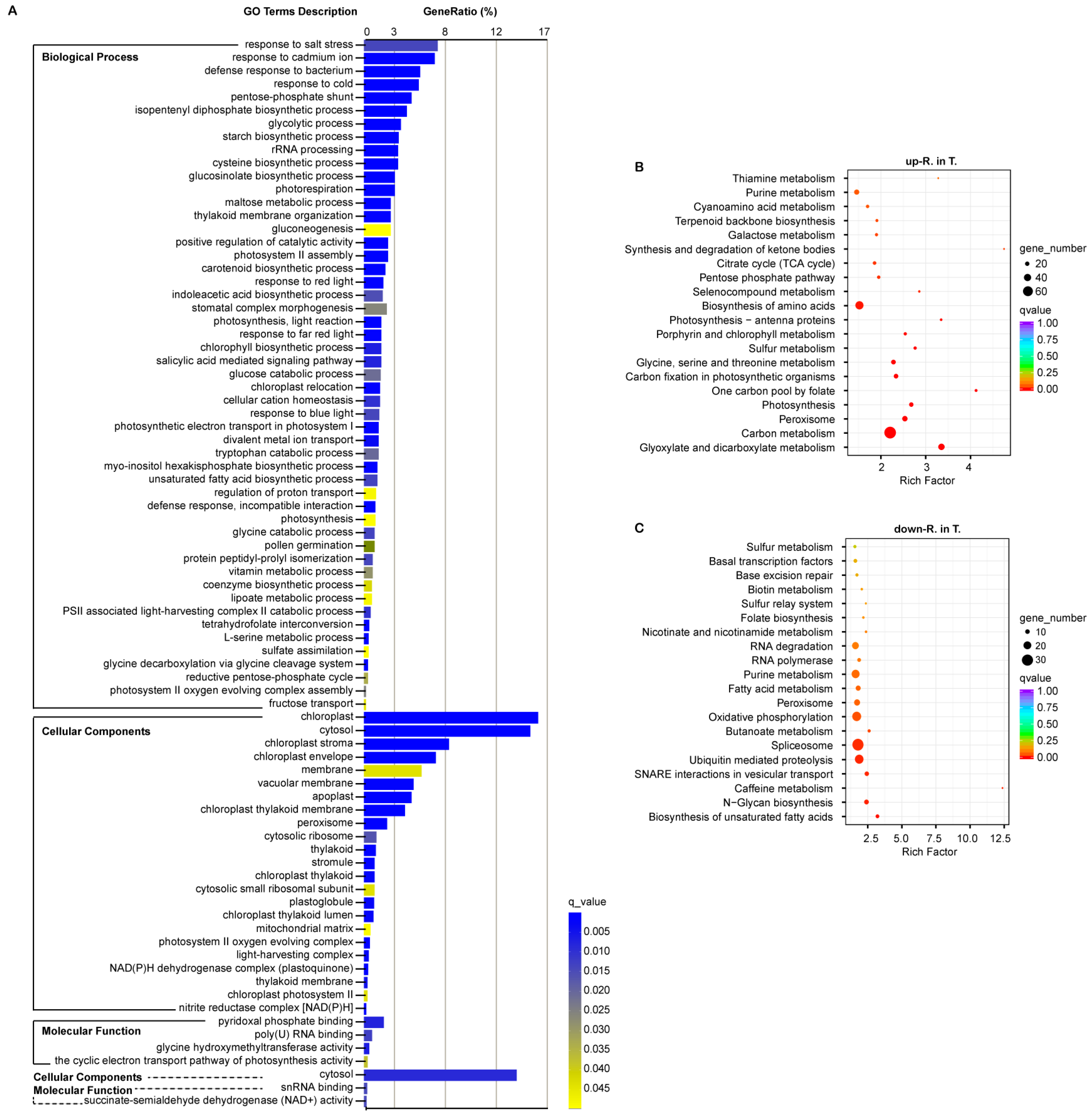
3.5. Gene Ontology Enrichment of the Important DEGs
3.6. Kyoto Encyclopedia of Genes and Genomes Enrichment of the Important DEGs
3.7. Candidate Gene Analysis
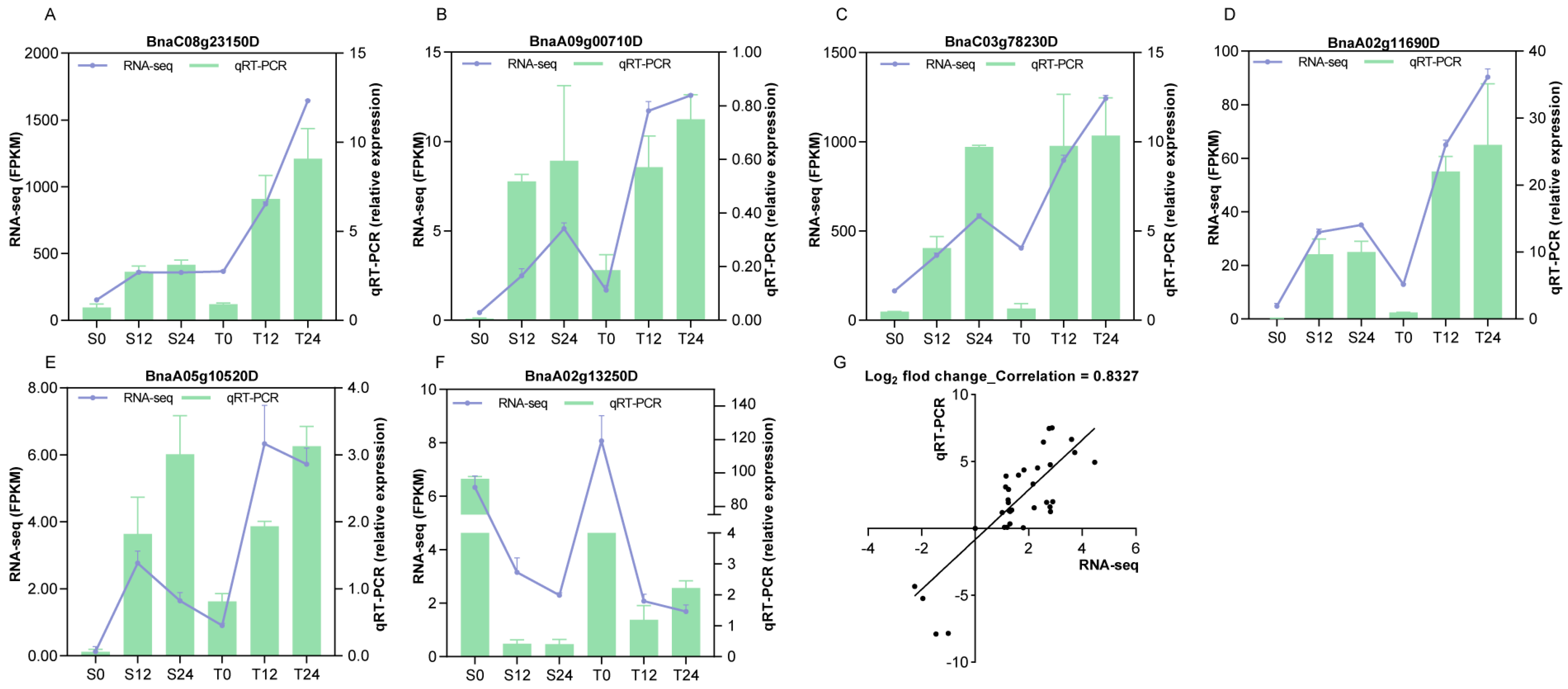
3.8. Validation of the RNA-Seq Data by qRT-PCR
4. Discussion
4.1. Alkali Salt Tolerance of Line 2205 Is Reflected at Multiple Levels
4.2. Synergistic Upregulation of Genes in Carbohydrates Metabolism, Photosynthetic Process, and ROS Equilibrium under NaHCO3 Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose. Front. Plant Sci. 2016, 7, 2035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J. Development and Prospect of the Research on Salt-Affected Soils in China. Acta Pedol. Snica 2008, 45, 837–845. [Google Scholar] [CrossRef]
- Yang, J.; Yao, R.; Wang, X.; Xie, W.; Zhang, X.; Zhu, W.; Zhang, L.; Sun, R. Research on Salt-Affected Soils in China: History, Status Quo and Prospect. Acta Pedol. Snica 2022, 59, 10–27. [Google Scholar] [CrossRef]
- Tuyen, D.D.; Lal, S.K.; Xu, D.H. Identification of a Major Qtl Allele from Wild Soybean (Glycine Soja Sieb. & Zucc.) for Increasing Alkaline Salt Tolerance in Soybean. Theor. Appl. Genet. 2010, 121, 229–236. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, Z.; Gao, S.; Lv, Y.; Chen, Z.; Cao, B.; Xu, K. Different Responses of Two Chinese Cabbage (Brassica rapa L. ssp. pekinensis) Cultivars in Photosynthetic Characteristics and Chloroplast Ultrastructure to Salt and Alkali Stress. Planta 2021, 254, 102. [Google Scholar] [CrossRef]
- Ruili, L.I.; Shi, F.; Fukuda, K.; Yang, Y. Effects of Salt and Alkali Stresses on Germination, Growth, Photosynthesis and Ion Accumulation in Alfalfa (Medicago Sativa L.). Soil Sci. Plant Nutr. 2010, 56, 725–733. [Google Scholar] [CrossRef]
- Choi, W.G.; Toyota, M.; Kim, S.H.; Hilleary, R.; Gilroy, S. Salt Stress-Induced Ca2+ Waves Are Associated with Rapid, Long-Distance Root-to-Shoot Signaling in Plants. Proc. Natl. Acad. Sci. USA 2014, 111, 6497–6502. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Wu, R.; Wee, C.W.; Xie, F.; Wei, X.; Chan, P.M.; Tham, C.; Duan, L.; Dinneny, J.R. A Spatio-Temporal Understanding of Growth Regulation During the Salt Stress Response in Arabidopsis. Plant Cell 2013, 25, 2132–2154. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Dai, S.; Wang, R.; Chen, S.; Li, N.; Zhou, X.; Lu, C.; Shen, X.; Zheng, X.; Hu, Z.; et al. Calcium Mediates Root K+/Na+ Homeostasis in Poplar Species Differing in Salt Tolerance. Tree Physiol. 2009, 29, 1175–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, D.M.; Oliveira, M.M.; Saibo, N.J.M. Regulation of Na+ and K+ Homeostasis in Plants: Towards Improved Salt Stress Tolerance in Crop Plants. Genet. Mol. Biol. 2017, 40, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive Oxygen Species Homeostasis and Signalling during Drought and Salinity Stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S.D. Chloroplast Function and Ion Regulation in Plants Growing on Saline Soils: Lessons from Halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhang, J.; Beckles, D.M. A Pivotal Role for Starch in the Reconfiguration of 14c-Partitioning and Allocation in Arabidopsis thaliana under Short-Term Abiotic Stress. Sci. Rep. 2018, 8, 9314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, S.; Wang, J.; Lin, J. Exogenous Abscisic Acid Alleviates Harmful Effect of Salt and Alkali Stresses on Wheat Seedlings. Int. J. Environ. Res. Public Health 2020, 17, 3770. [Google Scholar] [CrossRef]
- Guo, R.; Yang, Z.; Li, F.; Yan, C.; Zhong, X.; Liu, Q.; Xia, X.; Li, H.; Zhao, L. Comparative Metabolic Responses and Adaptive Strategies of Wheat (Triticum Aestivum) to Salt and Alkali Stress. BMC Plant Biol. 2015, 15, 170. [Google Scholar] [CrossRef] [Green Version]
- Gong, B.; Wen, D.; Bloszies, S.; Li, X.; Wei, M.; Yang, F.J.; Shi, Q.H.; Wang, X.F. Comparative Effects of Nacl and Nahco3 Stresses on Respiratory Metabolism, Antioxidant System, Nutritional Status, and Organic Acid Metabolism in Tomato Roots. Acta Physiol. Plant 2014, 36, 2167–2181. [Google Scholar] [CrossRef]
- Li, X.; Zheng, H.; Wu, W.; Liu, H.; Wang, J.; Jia, Y.; Li, J.; Yang, L.; Lei, L.; Zou, D.; et al. Qtl Mapping and Candidate Gene Analysis for Alkali Tolerance in Japonica Rice at the Bud Stage Based on Linkage Mapping and Genome-Wide Association Study. Rice 2020, 13, 48. [Google Scholar] [CrossRef]
- Li, N.; Zheng, H.; Cui, J.; Wang, J.; Liu, H.; Sun, J.; Liu, T.; Zhao, H.; Lai, Y.; Zou, D. Genome-Wide Association Study and Candidate Gene Analysis of Alkalinity Tolerance in Japonica Rice Germplasm at the Seedling Stage. Rice 2019, 12, 24. [Google Scholar] [CrossRef]
- He, Y.; Dong, Y.; Yang, X.; Guo, D.; Qian, X.; Yan, F.; Wang, Y.; Li, J.; Wang, Q. Functional Activation of a Novel R2r3-Myb Protein Gene, Gmmyb68, Confers Salt-Alkali Resistance in Soybean (Glycine Max L.). Genome 2020, 63, 13–26. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, X.; Xu, C.; Guo, D.; Niu, L.; Wang, Y.; Li, J.; Yan, F.; Wang, Q. Overexpression of a Novel Transcriptional Repressor Gmmyb3a Negatively Regulates Salt-Alkali Tolerance and Stress-Related Genes in Soybean. Biochem. Biophys. Res. Commun. 2018, 498, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Cai, H.; Luo, X.; Bai, X.; Deyholos, M.K.; Chen, Q.; Chen, C.; Ji, W.; Zhu, Y. Over-Expression of a Novel Jaz Family Gene from Glycine Soja, Increases Salt and Alkali Stress Tolerance. Biochem. Biophys. Res. Commun. 2012, 426, 273–279. [Google Scholar] [CrossRef]
- Guan, Q.J.; Wang, L.F.; Bu, Q.Y.; Wang, Z.Y. The Rice Gene Oszfp6 Functions in Multiple Stress Tolerance Responses in Yeast and Arabidopsis. Plant Physiol. Biochem. 2014, 82, 1–8. [Google Scholar] [CrossRef]
- Guan, Q.J.; Ma, H.Y.; Wang, Z.J.; Wang, Z.Y.; Bu, Q.Y.; Liu, S.K. A Rice Lsd1-Like-Type Zfp Gene Oslol5 Enhances Saline-Alkaline Tolerance in Transgenic Arabidopsis thaliana, Yeast and Rice. BMC Genom. 2016, 17, 142. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Yang, X.X.; Zhang, L.; Zhang, J.; Du, B.; Yao, L.; Li, X.T.; Guo, C. Alfalfa Mscbl4 Enhances Calcium Metabolism but Not Sodium Transport in Transgenic Tobacco under Salt and Saline-Alkali Stress. Plant Cell Rep. 2020, 39, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Chen, N.; Song, L.; Wang, D.; Cai, H.; Yao, L.; Li, X.; Guo, C. Alfalfa (Medicago sativa L.) Mscml46 Gene Encoding Calmodulin-Like Protein Confers Tolerance to Abiotic Stress in Tobacco. Plant Cell Rep. 2021, 40, 1907–1922. [Google Scholar] [CrossRef]
- Zheng, M.; Zhang, L.; Tang, M.; Liu, J.; Liu, H.; Yang, H.; Fan, S.; Terzaghi, W.; Wang, H.; Hua, W. Knockout of Two Bnamax1 Homologs by Crispr/Cas9-Targeted Mutagenesis Improves Plant Architecture and Increases Yield in Rapeseed (Brassica napus L.). Plant Biotechnol. J. 2020, 18, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, I.A.A.; Shalby, N.; El-Badri, A.M.; Batool, M.; Wang, C.; Wang, Z.; Salah, A.; Rady, M.M.; Jie, K.; Wang, B.; et al. Rna-Seq Analysis Revealed Key Genes Associated with Salt Tolerance in Rapeseed Germination through Carbohydrate Metabolism, Hormone, and Mapk Signaling Pathways. Ind. Crops Prod. 2022, 176, 114262. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, G.; Du, L.; Shang, X.; Cheng, C.; Yang, B.; Hu, Y.; Cai, C.; Guo, W. Genetic Regulation of Salt Stress Tolerance Revealed by Rna-Seq in Cotton Diploid Wild Species, Gossypium davidsonii. Sci. Rep. 2016, 6, 20582. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.S. The Water Culture Method for Growing Plants without Soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 32. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Nacl-Induced Senescence in Leaves of Rice ( Oryza Sativa L.) Cultivars Differing in Salinity Resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, F.; Xu, W.; Di, C.; Zhou, S.; Xue, Y.; Yu, J.; Su, Z. Increased Expression of Osspx1 Enhances Cold/Subfreezing Tolerance in Tobacco and Arabidopsis thaliana. Plant Biotechnol. J. 2009, 7, 550–561. [Google Scholar] [CrossRef]
- Kannangara, C.G.; Woolhouse, H.W. Changes in the Enzyme Activity of Soluble Protein Fractions in the Course of Foliar Senescence in Perilla frutescens (L.) Britt. New Phytol. 1968, 67, 533–542. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide Dismutase: Improved Assays and an Assay Applicable to Acrylamide Gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Zhao, X.; Wei, P.; Liu, Z.; Yu, B.; Shi, H. Soybean Na+/H+ Antiporter Gmssos1 Enhances Antioxidant Enzyme Activity and Reduces Na+ Accumulation in Arabidopsis and Yeast Cells under Salt Stress. Acta Physiol. Plant. 2016, 39, 19. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. Hisat: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of Exon Skipping Events Differentiate among Splicing Patterns in Sixteen Human Tissues. F1000Research 2013, 2, 188. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Genome Biol 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene Ontology Analysis for Rna-Seq: Accounting for Selection Bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. Kobas 2.0: A Web Server for Annotation and Identification of Enriched Pathways and Diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative Pcr and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Chen, K.C.; Cheng, T.S.; Lee, C.; Lin, S.H.; Tung, C.W. Chlorophyll Fluorescence Analysis in Diverse Rice Varieties Reveals the Positive Correlation between the Seedlings Salt Tolerance and Photosynthetic Efficiency. BMC Plant Biol. 2019, 19, 403. [Google Scholar] [CrossRef] [Green Version]
- Watson, S.J.; Sowden, R.G.; Jarvis, P. Abiotic Stress-Induced Chloroplast Proteome Remodelling: A Mechanistic Overview. J. Exp. Bot. 2018, 69, 2773–2781. [Google Scholar] [CrossRef]
- Lang, L.; Xu, A.; Ding, J.; Zhang, Y.; Zhao, N.; Tian, Z.; Liu, Y.; Wang, Y.; Liu, X.; Liang, F.; et al. Quantitative Trait Locus Mapping of Salt Tolerance and Identification of Salt-Tolerant Genes in Brassica napus L. Front. Plant Sci. 2017, 8, 1000. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.B.; Mohamed, S.; Stephen, B.; Harold, B.; Sabah, M. Cell Membrane Stability and Association Mapping for Drought and Heat Tolerance in a Worldwide Wheat Collection. Sustainability 2017, 9, 1606. [Google Scholar] [CrossRef] [Green Version]
- Thalmann, M.; Santelia, D. Starch as a Determinant of Plant Fitness under Abiotic Stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, I.A.A.; Shalby, N.; Bai, C.; Qin, M.; Agami, R.A.; Jie, K.; Wang, B.; Zhou, G. Stomatal and Photosynthetic Traits Are Associated with Investigating Sodium Chloride Tolerance of Brassica napus L. Cultivars. Plants 2020, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a Multifaceted Signalling Molecule in Plant Responses to Abiotic Stress: Understanding the Physiological Mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt Tolerance and Salinity Effects on Plants: A Review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Krzesłowska, M. The Cell Wall in Plant Cell Response to Trace Metals: Polysaccharide Remodeling and Its Role in Defense Strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Guo, Q.; Liu, P.; Huang, J.; Zhang, S.; Yang, G.; Wu, C.; Zheng, C.; Yan, K. Dual Roles of the Serine/Arginine-Rich Splicing Factor Sr45a in Promoting and Interacting with Nuclear Cap-Binding Complex to Modulate the Salt-Stress Response in Arabidopsis. New Phytol. 2021, 230, 641–655. [Google Scholar] [CrossRef]
- Hu, Q.; Chen, Y.; Zhao, Y.; Gu, J.; Ma, M.; Li, H.; Li, C.; Wang, Z.Y. Overexpression of Scl30a from Cassava (Manihot Esculenta) Negatively Regulates Salt Tolerance in Arabidopsis. Funct. Plant Biol. 2021, 48, 1213–1224. [Google Scholar] [CrossRef]
- Wang, W.; Pang, J.; Zhang, F.; Sun, L.; Yang, L.; Zhao, Y.; Yang, Y.; Wang, Y.; Siddique, K.H.M. Integrated Transcriptomics and Metabolomics Analysis to Characterize Alkali Stress Responses in Canola (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 605–620. [Google Scholar] [CrossRef]
- Abd El-Moneim, D.; Contreras, R.; Silva-Navas, J.; Gallego, F.J.; Figueiras, A.M.; Benito, C. On the Consequences of Aluminium Stress in Rye: Repression of Two Mitochondrial Malate Dehydrogenase Mrnas. Plant Biol. 2015, 17, 123–133. [Google Scholar] [CrossRef]
- Kim, S.C.; Guo, L.; Wang, X. Nuclear Moonlighting of Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenase Regulates Arabidopsis Response to Heat Stress. Nat. Commun. 2020, 11, 3439. [Google Scholar] [CrossRef]
- Liu, T.; Fang, H.; Liu, J.; Reid, S.; Hou, J.; Zhou, T.; Tian, Z.; Song, B.; Xie, C. Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenases Play Crucial Roles in Controlling Cold-Induced Sweetening and Apical Dominance of Potato (Solanum tuberosum L.) Tubers. Plant Cell Environ. 2017, 40, 3043–3054. [Google Scholar] [CrossRef]
- Schneider, M.; Knuesting, J.; Birkholz, O.; Heinisch, J.J.; Scheibe, R. Cytosolic Gapdh as a Redox-Dependent Regulator of Energy Metabolism. BMC Plant Biol. 2018, 18, 184. [Google Scholar] [CrossRef] [Green Version]
- Vescovi, M.; Zaffagnini, M.; Festa, M.; Trost, P.; Lo Schiavo, F.; Costa, A. Nuclear Accumulation of Cytosolic Glyceraldehyde-3-Phosphate Dehydrogenase in Cadmium-Stressed Arabidopsis Roots. Plant Physiol. 2013, 162, 333–346. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.; Singh, R.P.; Tai, G.C. Molecular Characterization and Expression Analysis of 1-Aminocyclopropane-1-Carboxylate Oxidase Homologs from Potato under Abiotic and Biotic Stresses. Genome 2002, 45, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; He, Z.; Patkar, R.; Zhu, Q.; Zhong, J.; Li, D.; Ronald, P.; Lamb, C.; Chattoo, B.B. Induction of H2O2 in Transgenic Rice Leads to Cell Death and Enhanced Resistance to Both Bacterial and Fungal Pathogens. Transgenic Res. 2003, 12, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Shortt, B.J.; Lawrence, E.B.; Levine, E.B.; Fitzsimmons, K.C.; Shah, D.M. Disease Resistance Conferred by Expression of a Gene Encoding H2O2-Generating Glucose Oxidase in Transgenic Potato Plants. Plant Cell 1995, 7, 1357–1368. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, D.; Miwa, T.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Ohsumi, C. Identification of Photorespiratory Glutamate:Glyoxylate Aminotransferase (Ggat) Gene in Arabidopsis. Plant J. 2003, 33, 975–987. [Google Scholar] [CrossRef]
- Yang, H.; Deng, L.; Liu, H.; Fan, S.; Hua, W.; Liu, J. Overexpression of Bnaaox1b Confers Tolerance to Osmotic and Salt Stress in Rapeseed. G3 2019, 9, 3501–3511. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yang, B.; Liu, W.Z.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.K.; Jiang, Y.Q. Identification and Characterization of Cbl and Cipk Gene Families in Canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 8. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Tao, S.; Zhu, Y.; Zhang, Q.; Li, P.; Wang, H.; Zhang, Y.; Bakirov, A.; Cao, H.; Qin, M.; et al. Identification of Alkaline Salt Tolerance Genes in Brassica napus L. by Transcriptome Analysis. Genes 2022, 13, 1493. https://doi.org/10.3390/genes13081493
Xu Y, Tao S, Zhu Y, Zhang Q, Li P, Wang H, Zhang Y, Bakirov A, Cao H, Qin M, et al. Identification of Alkaline Salt Tolerance Genes in Brassica napus L. by Transcriptome Analysis. Genes. 2022; 13(8):1493. https://doi.org/10.3390/genes13081493
Chicago/Turabian StyleXu, Yu, Shunxian Tao, Yunlin Zhu, Qi Zhang, Ping Li, Han Wang, Yan Zhang, Aldiyar Bakirov, Hanming Cao, Mengfan Qin, and et al. 2022. "Identification of Alkaline Salt Tolerance Genes in Brassica napus L. by Transcriptome Analysis" Genes 13, no. 8: 1493. https://doi.org/10.3390/genes13081493