
Evaluation of the Impact of the Peregrine Falcon (Falco peregrinus peregrinus) Reintroduction Process on Captive-Bred Population


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Microsatellite Genotyping
2.3. Statistical Analyses
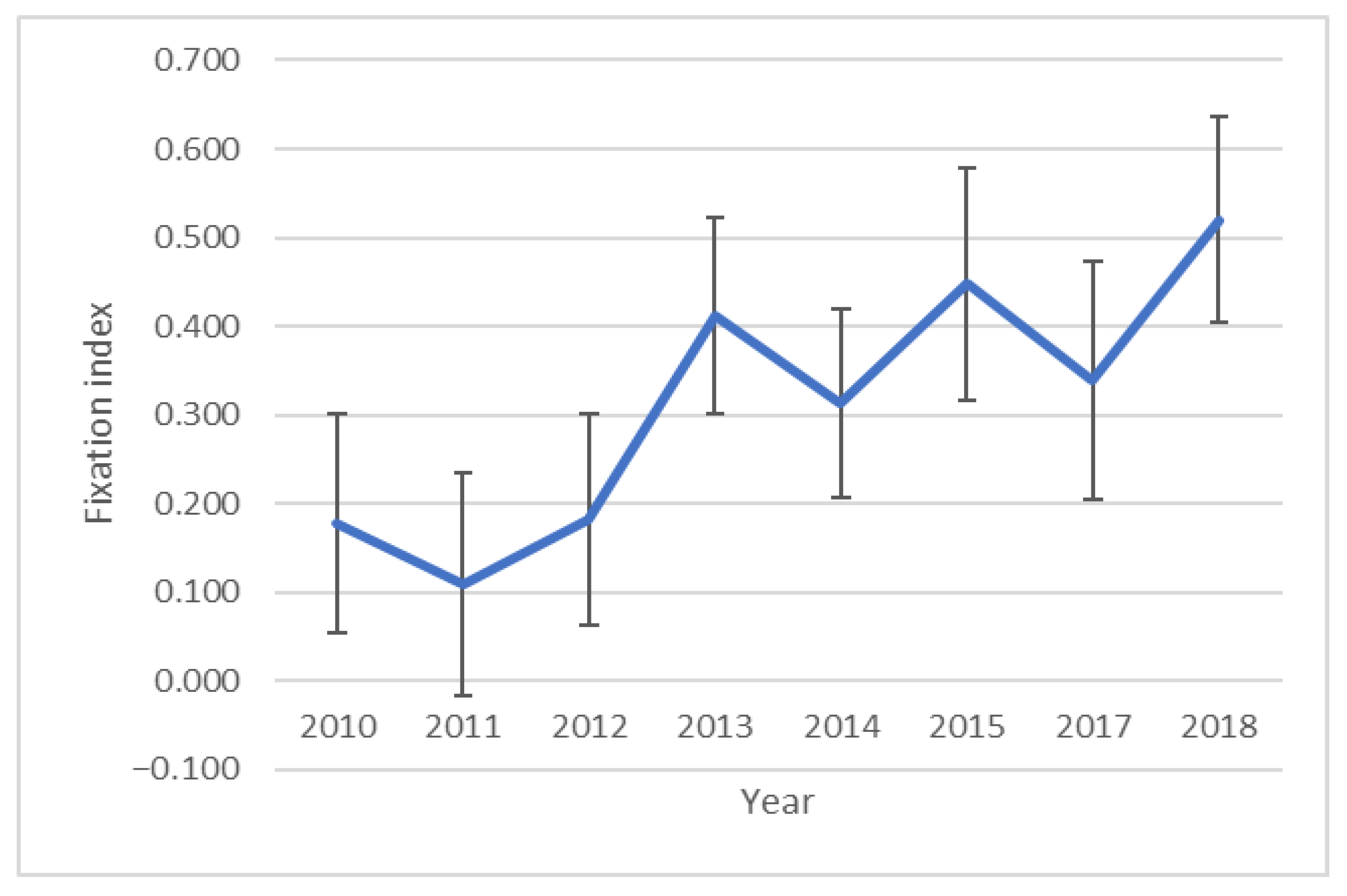
3. Results
3.1. All Samples
3.2. Individuals from the Six Main Providers
4. Discussion
4.1. Variability in the Population
4.2. Variability among Years
4.3. Possible Reasons for the Change in Genetic Structure
4.4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kolnegari, M.; Jamali, M.; Naserifard, M.; Ghous, K.; Hazrati, M.; Panter, C.T.; Dwyer, J.F. Falconry petroglyphs in Iran: New findings on the nexus between ancient humans and birds of prey. Eur. J. Wildl. Res. 2021, 67, 1–9. [Google Scholar] [CrossRef]
- Sielicki, S.; Sielicki, J. Restoration of peregrine falcon in Poland 1989–2007. In Peregrine Falcon Populations—Status and Perspectives in 21st Century, 1st ed.; Sielicki, J., Mizera, B., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2009; Volume 1, pp. 699–722. [Google Scholar]
- Bryndová, M.; Putnová, L.; Bartoňová, P.; Kaplanová, K.; Urban, T. Genetic differences between wild and captive populations of the peregrine falcon (Falco peregrinus) and the saker falcon (Falco cherrung) living in the Czech Republic. J. Agric. Sci. Technol. B 2012, 2, 642–651. [Google Scholar]
- Wegner, P.; Kleinstäuber, G.; Baum, F.; Schilling, F. Long-term investigation of the degree of exposure of German peregrine falcons (Falco peregrinus) to damaging chemicals from the environment. J. Ornithol. 2005, 146, 34–54. [Google Scholar] [CrossRef]
- Seddon, P.J.; Armstrong, D.P.; Maloney, R.F. Developing the science of reintroduction biology. Conserv. Biol. 2007, 21, 303–312. [Google Scholar] [CrossRef]
- Lindberg, P. The fall and the rise of the Swedish Peregrine Falcon population. In Peregrine Falcon Populations—Status and Perspectives in the 21st Century; Sielicki, J., Mizera, T., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2009; pp. 137–144. [Google Scholar]
- Monneret, R.-J. Evolution and current situation of the Peregrine Falcon in France. In Peregrine Falcon Populations—Status and Perspectives in the 21st Century; Sielicki, J., Mizera, T., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2009; pp. 169–174. [Google Scholar]
- Wegner, P.; Brücher, H.; Brücher, S.; Höller, T.; Jöbges, M.; Kladny, M.; Lindner, M.; Sell, G.; Speer, G.; Thomas, T.; et al. Development of an urban population of Peregrine Falcons in North Rhine-Westphalia, Germany: Population status, nest site selection and some biological results. In Peregrine Falcon Populations—Status and Perspectives in the 21st Century; Sielicki, J., Mizera, T., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2009; pp. 729–746. [Google Scholar]
- Heinrich, W. Peregrine Falcon recovery in the continental United States, 1974–1999, with notes on related programs of The Peregrine Fund. In Peregrine Falcon Populations—Status and Perspectives in the 21st Century; Sielicki, J., Mizera, T., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2009; pp. 429–442. [Google Scholar]
- Banks, A.N.; Crick, H.Q.P.; Coombes, R.; Benn, S.; Ratcliffe, D.A.; Humphreys, E.M. The breeding status of peregrine falcons Falco peregrinus in the UK and Isle of Man in 2002. Bird Study 2010, 57, 421–436. [Google Scholar] [CrossRef] [Green Version]
- Petrov, R.; Andonova, Y.; Gancheva, Y.; Klisurov, I. Implications of captive breeding for the reintroduction of the Saker falcon (Falco cherrug) in Bulgaria. Agric. Sci. Technol. 2021, 13, 300–306. [Google Scholar] [CrossRef]
- Sielicki, S.; Sielicki, J. Falconry and the restoration of the peregrine falcon in Poland in 1990–2015. In Falconary—Its Influence on Biodiversity and Cultural Heritage, 1st ed.; Szymak, U., Sianko, P., Eds.; Muzeum Podlaskie w Białymstoku: Białystok, Poland, 2016; Volume 1, pp. 41–47. [Google Scholar]
- Ivanovsky, V.; Vintchevsky, A. Status of the Peregrine Falcon in Belarus. In Peregrine Falcon Populations—Status and Perspectives in the 21st Century; Sielicki, J., Mizera, T., Eds.; Turul–Poznań University of Life Sciences Press: Warsaw, Poland; Poznań, Poland, 2008; pp. 117–120. [Google Scholar]
- Brambilla, M.; Rubolini, D.; Guidali, F. Factors affecting breeding habitat selection in a cliff-nesting peregrine Falco peregrinus population. J. Ornithol. 2006, 147, 428–435. [Google Scholar] [CrossRef]
- Ławicki, Ł.; Sielicki, S. Restoration of the tree-nesting population of the peregrine falcon Falco peregrinus in Pomerania. Ornis Pol. 2019, 60, 75–84. [Google Scholar]
- Nesje, M.; Røed, K.H.; Bell, D.A.; Lindberg, P.; Lifjeld, J.T. Microsatellite analysis of population structure and genetic variability in peregrine falcons (Falco peregrinus). Anim. Conserv. 2000, 3, 267–275. [Google Scholar] [CrossRef]
- Nittinger, F.; Gamauf, A.; Pinsker, W.; Wink, M.; Haring, E. Phylogeography and population structure of the saker falcon (Falco cherrug) and the influence of hybridization: Mitochondrial and microsatellite data. Mol. Ecol. 2007, 16, 1497–1517. [Google Scholar] [CrossRef]
- Jacobsen, F.; Nesje, M.; Bachmann, L.; Lifjeld, J.T. Significant genetic admixture after reintroduction of peregrine falcon (Falco peregrinus) in Southern Scandinavia. Conserv. Genet. 2008, 9, 581–591. [Google Scholar] [CrossRef]
- Ponnikas, S.; Ollila, T.; Kvist, L. Turnover and post-bottleneck genetic structure in a recovering population of Peregrine Falcons Falco peregrinus. Ibis 2017, 159, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Puchała, K.O.; Nowak-Życzyńska, Z.; Sielicki, S.; Olech, W. Assessment of the genetic potential of the peregrine falcon (Falco peregrinus peregrinus) population used in the reintroduction program in Poland. Genes 2021, 12, 666. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Marshall, T.C.; Slate, J.; Kruuk, L.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; von Holdt, B.M. Structure Harvester: A website and program for visualizing Structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software Structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mengoni, C.; Zuberogoitia, I.; Mucci, N.; Boano, G.; Urban, T.; Guzzo, E.; Sarà, M. Genetic variability in Peregrine Falcon populations of the Western Palaearctic region. Ornis Hung. 2018, 26, 12–26. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Allele Ranges | N | Na | Ne | Ho | He | F |
|---|---|---|---|---|---|---|---|
| NVHfp107 | 203–208 | 373 | 4 | 2.082 | 0.466 | 0.520 | 0.102 |
| NVHfp13 | 93–103 | 373 | 9 | 4.654 | 0.488 | 0.785 | 0.379 |
| NVHfp46_1 | 117–122 | 373 | 6 | 4.129 | 0.539 | 0.758 | 0.289 |
| NVHfp5 | 102–108 | 374 | 5 | 1.462 | 0.070 | 0.316 | 0.780 |
| NVHfp54 | 104–208 | 369 | 9 | 2.487 | 0.428 | 0.598 | 0.284 |
| NVHfp82_2 | 134–140 | 373 | 5 | 1.648 | 0.137 | 0.393 | 0.652 |
| NVHfp86_2 | 140–145 | 368 | 5 | 3.453 | 0.351 | 0.710 | 0.507 |
| NVHfp89 | 116–132 | 367 | 9 | 4.831 | 0.569 | 0.793 | 0.282 |
| NVHfp92_1 | 110–126 | 365 | 7 | 3.011 | 0.288 | 0.668 | 0.569 |
| Locus | Allele Ranges | N | Na | Ne | Ho | He | F |
|---|---|---|---|---|---|---|---|
| NVHfp107 | 203–208 | 170 | 4 | 1.951 | 0.465 | 0.488 | 0.047 |
| NVHfp13 | 96–103 | 169 | 8 | 4.161 | 0.349 | 0.760 | 0.540 |
| NVHfp46_1 | 117–122 | 170 | 6 | 3.886 | 0.494 | 0.743 | 0.335 |
| NVHfp5 | 102–108 | 170 | 4 | 1.709 | 0.082 | 0.415 | 0.801 |
| NVHfp54 | 104–115 | 170 | 6 | 2.425 | 0.441 | 0.588 | 0.249 |
| NVHfp82_2 | 134–140 | 169 | 5 | 2.509 | 0.195 | 0.602 | 0.675 |
| NVHfp86_2 | 140–145 | 168 | 5 | 3.716 | 0.315 | 0.731 | 0.568 |
| NVHfp89 | 116–132 | 167 | 9 | 5.598 | 0.575 | 0.821 | 0.300 |
| NVHfp92_1 | 110–124 | 168 | 6 | 3.4051 | 0.310 | 0.706 | 0.562 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puchała, K.O.; Nowak-Życzyńska, Z.; Sielicki, S.; Olech, W. Evaluation of the Impact of the Peregrine Falcon (Falco peregrinus peregrinus) Reintroduction Process on Captive-Bred Population. Genes 2022, 13, 1487. https://doi.org/10.3390/genes13081487
Puchała KO, Nowak-Życzyńska Z, Sielicki S, Olech W. Evaluation of the Impact of the Peregrine Falcon (Falco peregrinus peregrinus) Reintroduction Process on Captive-Bred Population. Genes. 2022; 13(8):1487. https://doi.org/10.3390/genes13081487
Chicago/Turabian StylePuchała, Karol O., Zuzanna Nowak-Życzyńska, Sławomir Sielicki, and Wanda Olech. 2022. "Evaluation of the Impact of the Peregrine Falcon (Falco peregrinus peregrinus) Reintroduction Process on Captive-Bred Population" Genes 13, no. 8: 1487. https://doi.org/10.3390/genes13081487