
Immune Transcriptome and Secretome Differ between Human CD71+ Erythroid Cells from Adult Bone Marrow and Fetal Liver Parenchyma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Study Interventions
2.3. Participant Safety
2.4. Cell Isolation
2.5. Magnetic Separation
2.6. Viability Staining
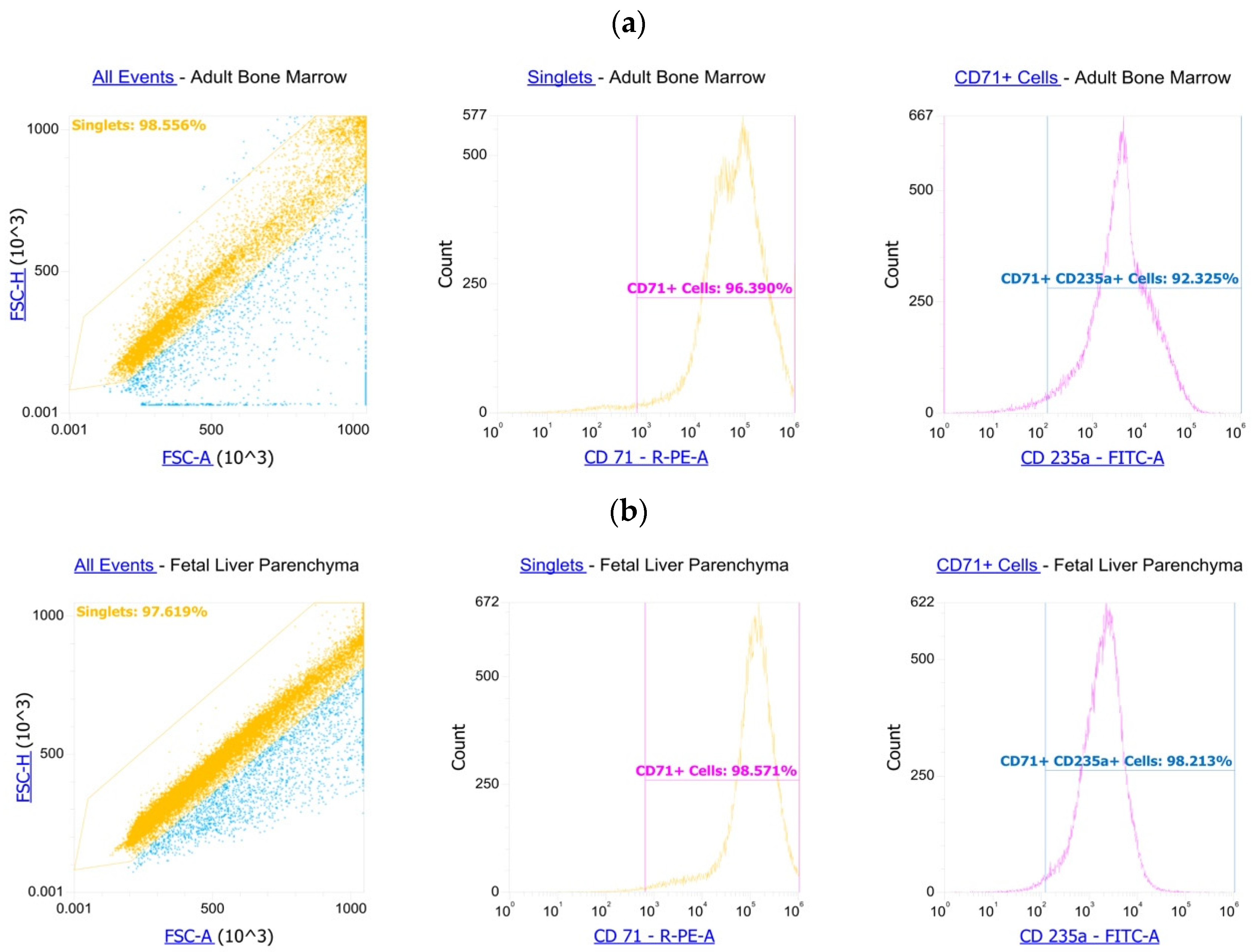
2.7. Flow Cytometry
2.8. Cell Culturing
2.9. Total RNA Extraction
2.10. Cell Culture Medium Harvesting
2.11. Immune Transcriptome Profiling
2.12. Cytokine Quantification in Culture Medium
2.13. Data Analysis
3. Results
3.1. Cytokine Gene Expression Profiles of the CECs
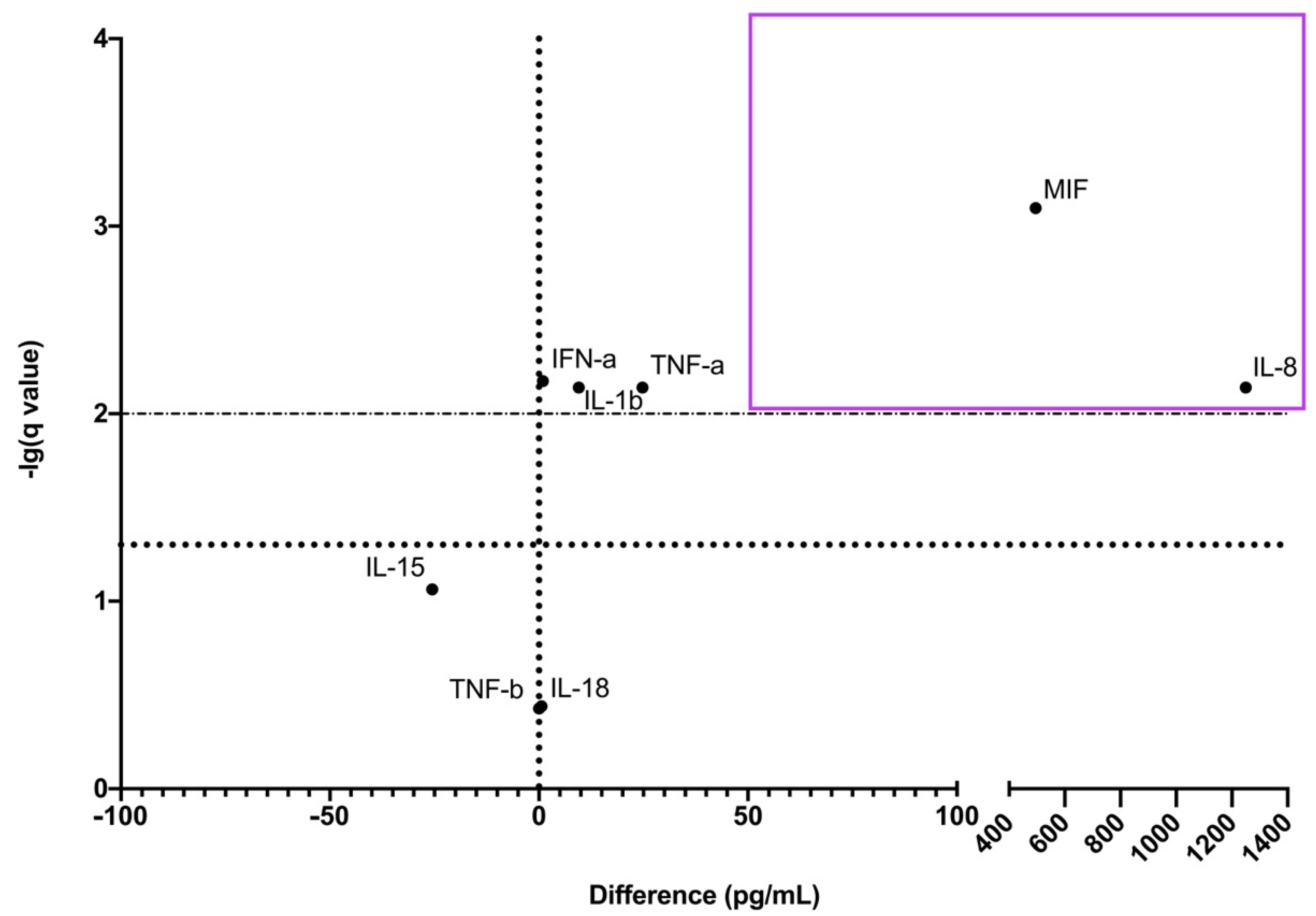
3.2. Cytokine Protein Profiles in CECs’ Culture Medium
3.3. CECs Have TLR Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elahi, S.; Vega-López, M.A.; Herman-Miguel, V.; Ramírez-Estudillo, C.; Mancilla-Ramírez, J.; Motyka, B.; West, L.; Oyegbami, O. CD71+ erythroid cells in human neonates exhibit immunosuppressive properties and compromise immune response against systemic infection in neonatal mice. Front. Immunol. 2020, 3076, 597433. [Google Scholar] [CrossRef] [PubMed]
- Grzywa, T.M.; Nowis, D.; Golab, J. The role of CD71+ erythroid cells in the regulation of the immune response. Pharmacol. Ther. 2021, 228, 107927. [Google Scholar] [CrossRef] [PubMed]
- Elahi, S.; Ertelt, J.M.; Kinder, J.M.; Jiang, T.T.; Zhang, X.; Xin, L.; Chaturvedi, V.; Strong, B.S.; Qualls, J.E.; Steinbrecher, K.A.; et al. Immunosuppressive CD71+ erythroid cells compromise neonatal host defence against infection. Nature 2013, 504, 158–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunsmore, G.; Bozorgmehr, N.; Delyea, C.; Koleva, P.; Namdar, A.; Elahi, S. Erythroid Suppressor Cells Compromise Neonatal Immune Response against Bordetella pertussis. J. Immunol. 2017, 199, 2081–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delyea, C.; Bozorgmehr, N.; Koleva, P.; Dunsmore, G.; Shahbaz, S.; Huang, V.; Elahi, S. CD71+ erythroid suppressor cells promote fetomaternal tolerance through arginase-2 and PDL-1. J. Immunol. 2018, 200, 4044–4058. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, S.; Bozorgmehr, N.; Koleva, P.; Namdar, A.; Jovel, J.; Fava, R.A.; Elahi, S. CD71+ VISTA+ erythroid cells promote the development and function of regulatory T cells through TGF-β. PLoS Biol. 2018, 16, e2006649. [Google Scholar] [CrossRef] [Green Version]
- Marsee, D.K.; Pinkus, G.S.; Yu, H. CD71 (transferrin receptor) an effective marker for erythroid precursors in bone marrow biopsy specimens. Am. J. Clin. Pathol. 2010, 134, 429–435. [Google Scholar] [CrossRef]
- Black, A.P.; Bhayani, H.; Ryder, C.A.; Gardner-Medwin, J.M.; Southwood, T.R. T-cell activation without proliferation in juvenile idiopathic arthritis. Arthritis Res. Ther. 2002, 4, 177. [Google Scholar] [CrossRef]
- Sennikov, S.V.; Eremina, L.V.; Samarin, D.M.; Avdeev, I.V.; Kozlov, V.A. Cytokine gene expression in erythroid cells. Eur. Cytokine Netw. 1996, 7, 771–774. [Google Scholar]
- Sennikov, S.V.; Injelevskaya, T.V.; Krysov, S.V.; Silkov, A.N.; Kovinev, I.B.; Dyachkova, N.J.; Zenkov, A.N.; Loseva, M.I.; Kozlov, V.A. Production of hemo-and immunoregulatory cytokines by erythroblast antigen+ and glycophorin A+ cells from human bone marrow. BMC Cell Biol. 2004, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Sennikov, S.V.; Krysov, S.V.; Injelevskaya, T.V.; Silkov, A.N.; Kozlov, V.A. Production of cytokines by immature erythroid cells derived from human embryonic liver. Eur. Cytokine Netw. 2001, 12, 274–279. [Google Scholar]
- Denisova, V.V.; Kulagin, A.D.; Lisukov, I.A.; Kryuchkova, I.V.; Sizikova, S.A.; Sennikov, S.V.; Kozlov, V.A. Cytokine-producing activity of bone marrow erythrokaryocytes and its regulation under normal conditions. Bull. Exp. Biol. Med. 2007, 143, 218–221. [Google Scholar] [CrossRef]
- Seledtsov, V.I.; Seledtsova, G.V.; Samarin, D.M.; Taraban, V.Y.; Sennikov, S.V.; Kozlov, V.A. Characterization of erythroid cell-derived natural suppressor activity. Immunobiology 1998, 198, 361–374. [Google Scholar] [CrossRef]
- Kingsley, P.D.; Malik, J.; Fantauzzo, K.A.; Palis, J. Yolk sac–derived primitive erythroblasts enucleate during mammalian embryogenesis. Blood 2004, 104, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, M.H.; Isern, J.; Fraser, S.T. The embryonic origins of erythropoiesis in mammals. Blood J. Am. Soc. Hematol. 2012, 119, 4828–4837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broxmeyer, H.E.; Douglas, G.W.; Hangoc, G.; Cooper, S.; Bard, J.; English, D.; Arny, M.; Thomas, L.; Boyse, E.A. Human umbilical cord blood as a potential source of transplantable hematopoietic stem/progenitor cells. Proc. Natl. Acad. Sci. USA 1989, 86, 3828–3832. [Google Scholar] [CrossRef] [Green Version]
- Moras, M.; Lefevre, S.D.; Ostuni, M.A. From erythroblasts to mature red blood cells: Organelle clearance in mammals. Front. Physiol. 2017, 8, 1076. [Google Scholar] [CrossRef]
- Coyne, J.D. Extramedullary haemopoiesis. J. Clin. Pathol. 2005, 58, 448. [Google Scholar]
- Shahbaz, S.; Xu, L.; Osman, M.; Sligl, W.; Shields, J.; Joyce, M.; Tyrrell, D.L.; Oyegbami, O.; Elahi, S. Erythroid precursors and progenitors suppress adaptive immunity and get invaded by SARS-CoV-2. Stem Cell Rep. 2021, 16, 1165–1181. [Google Scholar] [CrossRef]
- Zhao, G.; Jiang, D.; Cai, W.; Chen, X.; Dong, W.; Chen, L.; Hong, G.; Wu, B.; Yao, Y.; Lu, Z.Q. CD71+ erythroid cells expansion in adult sepsis: Potential causes and role in prognosis and nosocomial infection prediction. Front. Immunol. 2022, 13, 830025. [Google Scholar] [CrossRef]
- Van Eeden, S.E.; Terashima, T. Interleukin 8 (IL-8) and the release of leukocytes from the bone marrow. Leuk. Lymphoma 2000, 37, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.B.; García-Gómez, A.; Garayoa, M.; Corchete, L.A.; Hernández, J.M.; San Miguel, J.; Gutierrez, N.C. Effects of IL-8 up-regulation on cell survival and osteoclastogenesis in multiple myeloma. Am. J. Pathol. 2016, 186, 2171–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Ryu, C.H.; Jun, J.A.; Kim, S.M.; Jeong, C.H.; Jeun, S.S. IL-8 enhances the angiogenic potential of human bone marrow mesenchymal stem cells by increasing vascular endothelial growth factor. Cell Biol. Int. 2014, 38, 1050–1059. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wang, Q.; Qian, J.; Li, Y.; Lu, Y.; Bi, E.; Zhang, M.; Li, T.; Yang, M.; Wu, Y.; et al. Macrophage Migration Inhibitory Factor Regulates Multiple Myeloma Bone Marrow Homing. Blood 2014, 124, 2015. [Google Scholar] [CrossRef]
- Takahashi, N.; Nishihira, J.; Sato, Y.; Kondo, M.; Ogawa, H.; Ohshima, T.; Une, Y.; Todo, S. Involvement of macrophage migration inhibitory factor (MIF) in the mechanism of tumor cell growth. Mol. Med. 1998, 4, 707–714. [Google Scholar] [CrossRef]
- Abu El-Asrar, A.M.; Ahmad, A.; Siddiquei, M.M.; De Zutter, A.; Allegaert, E.; Gikandi, P.W.; De Hertogh, G.; Van Damme, J.; Opdenakker, G.; Struyf, S. The proinflammatory and proangiogenic macrophage migration inhibitory factor is a potential regulator in proliferative diabetic retinopathy. Front. Immunol. 2019, 10, 2752. [Google Scholar] [CrossRef] [Green Version]
- Liew, F.Y.; McInnes, I.B. Role of interleukin 15 and interleukin 18 in inflammatory response. Ann. Rheum. Dis. 2002, 61 (Suppl. 2), ii100–ii102. [Google Scholar] [CrossRef]
- Calmon-Hamaty, F.; Combe, B.; Hahne, M.; Morel, J. Lymphotoxin α stimulates proliferation and pro-inflammatory cytokine secretion of rheumatoid arthritis synovial fibroblasts. Cytokine 2011, 53, 207–214. [Google Scholar] [CrossRef]
- Ashkar, A.A.; Rosenthal, K.L. Toll-like receptor 9, CpG DNA and innate immunity. Curr. Mol. Med. 2002, 2, 545–556. [Google Scholar] [CrossRef]
- Takeda, K.; Takeuchi, O.; Akira, S. Recognition of lipopeptides by Toll-like receptors. J. Endotoxin Res. 2002, 8, 459–463. [Google Scholar] [CrossRef]
- Girardin, S.E.; Boneca, I.G.; Viala, J.; Chamaillard, M.; Labigne, A.; Thomas, G.; Philpott, D.J.; Sansonetti, P.J. Nod2 is a general sensor of peptidoglycan through muramyl dipeptide (MDP) detection. J. Biol. Chem. 2003, 278, 8869–8872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, M.F.; Wiens, G.D.; Purcell, M.K.; Palti, Y. Characterization of Toll-like receptor 3 gene in rainbow trout (Oncorhynchus mykiss). Immunogenetics 2005, 57, 510–519. [Google Scholar] [CrossRef] [PubMed]
- Morera, D.; Roher, N.; Ribas, L.; Balasch, J.C.; Doñate, C.; Callol, A.; Boltaña, S.; Roberts, S.; Goetz, G.; Goetz, F.W.; et al. RNA-Seq reveals an integrated immune response in nucleated erythrocytes. PLoS ONE 2011, 6, e26998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St Paul, M.; Paolucci, S.; Barjesteh, N.; Wood, R.D.; Sharif, S. Chicken erythrocytes respond to Toll-like receptor ligands by up-regulating cytokine transcripts. Res. Vet. Sci. 2013, 95, 87–91. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ABM | FL | ||||
|---|---|---|---|---|---|
| Cytokine | Presence | № of Samples Positive | Sex | Presence | № of Samples Positive |
| IFNA1 | + | 4/4 | F, M | + | 4/4 |
| IL1B | + | 4/4 | F, M | + | 3/4 |
| IL8 | + | 4/4 | F, M | + | 3/4 |
| IL15 | + | 2/4 | F, M | + | 4/4 |
| IL18 | + | 2/4 | F, M | + | 3/4 |
| IL32 | + | 4/4 | F, M | + | 4/4 |
| MIF | + | 4/4 | F, M | + | 0/4 |
| LTA | − | 0/4 | − | + | 3/4 |
| TNF | − | 0/4 | − | + | 1/4 |
| TGFB1 | + | 2/4 | F | − | 0/4 |
| Adult Bone Marrow CECs | Fetal Liver Parenchyma CECs | |||||||
|---|---|---|---|---|---|---|---|---|
| Cytokine | Mean | SD | № of Samples Positive | mRNA Presence | Mean | SD | № of Samples Positive | mRNA Presence |
| pg/mL | pg/mL | n | +/− | pg/mL | pg/mL | n | +/− | |
| IFN-a | 20.665 | 3.971 | 6/6 | + | 11.157 | 6.311 | 5/6 | + |
| IL-1b | 1.102 | 0.631 | 5/6 | + | 0.25 | 0.102 | 1/6 | + |
| IL-8 | 293.99 | 224.772 | 6/6 | + | 0 | 0 | 0/6 | + |
| IL-15 | 0 | 0 | 0/6 | + | 25.566 | 41.769 | 6/6 | + |
| IL-18 | 0.4 | 0.98 | 1/6 | + | 0.372 | 0.368 | 4/6 | + |
| MIF | 639.667 | 83.991 | 6/6 | + | 14.1 | 5.814 | 6/6 | + |
| TNF-a | 0 | 0 | 0/6 | − | 3.5 | 0.588 | 6/6 | + |
| TNF-b | 0 | 0 | 0/6 | − | 4.683 | 3.463 | 6/6 | + |
| ABM | FL | ||||
|---|---|---|---|---|---|
| Pattern-Recognition Receptor | Presence | № of Samples Positive | Sex | Presence | № of Samples Positive |
| TLR1 | − | 0/4 | F, M | + | 4/4 |
| TLR2 | + | 4/4 | F, M | + | 4/4 |
| TLR9 | + | 4/4 | F, M | + | 4/4 |
| NOD2 | − | 0/4 | F, M | + | 3/4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perik-Zavodskii, R.; Perik-Zavodskaya, O.; Shevchenko, Y.; Denisova, V.; Nazarov, K.; Obleuhova, I.; Zaitsev, K.; Sennikov, S. Immune Transcriptome and Secretome Differ between Human CD71+ Erythroid Cells from Adult Bone Marrow and Fetal Liver Parenchyma. Genes 2022, 13, 1333. https://doi.org/10.3390/genes13081333
Perik-Zavodskii R, Perik-Zavodskaya O, Shevchenko Y, Denisova V, Nazarov K, Obleuhova I, Zaitsev K, Sennikov S. Immune Transcriptome and Secretome Differ between Human CD71+ Erythroid Cells from Adult Bone Marrow and Fetal Liver Parenchyma. Genes. 2022; 13(8):1333. https://doi.org/10.3390/genes13081333
Chicago/Turabian StylePerik-Zavodskii, Roman, Olga Perik-Zavodskaya, Yulia Shevchenko, Vera Denisova, Kirill Nazarov, Irina Obleuhova, Konstantin Zaitsev, and Sergey Sennikov. 2022. "Immune Transcriptome and Secretome Differ between Human CD71+ Erythroid Cells from Adult Bone Marrow and Fetal Liver Parenchyma" Genes 13, no. 8: 1333. https://doi.org/10.3390/genes13081333