
The Effect of EGR1 on the Proliferation of Dermal Papilla Cells

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Cloning of the EGR1 CDS Region
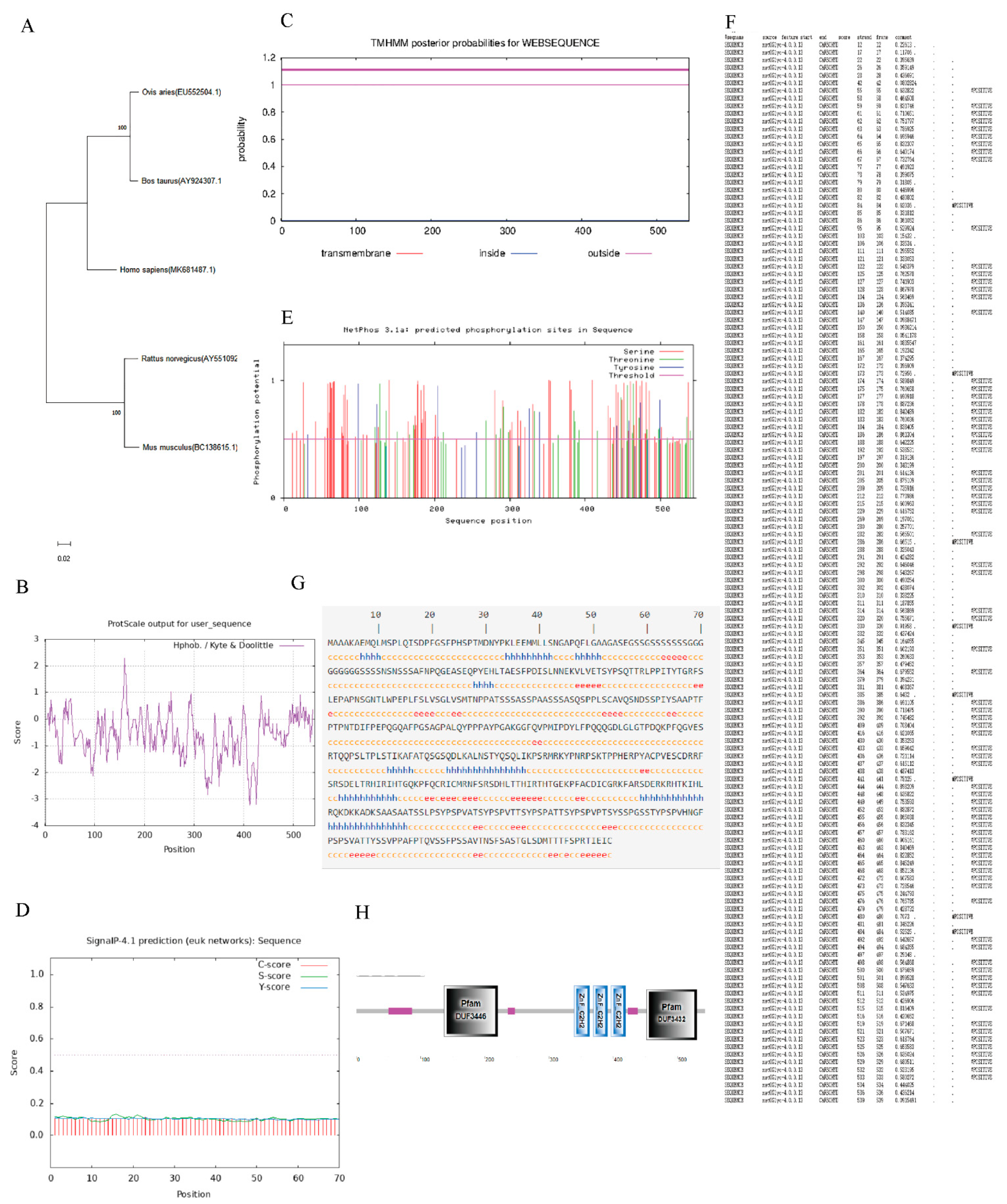
2.3. Bioinformatics Analysis of EGR1
2.4. Overexpression Vector Construction and siRNA Synthesis
2.5. Cell Culture
2.6. Cell Proliferation
2.7. RT-qPCR
2.8. Western Blot
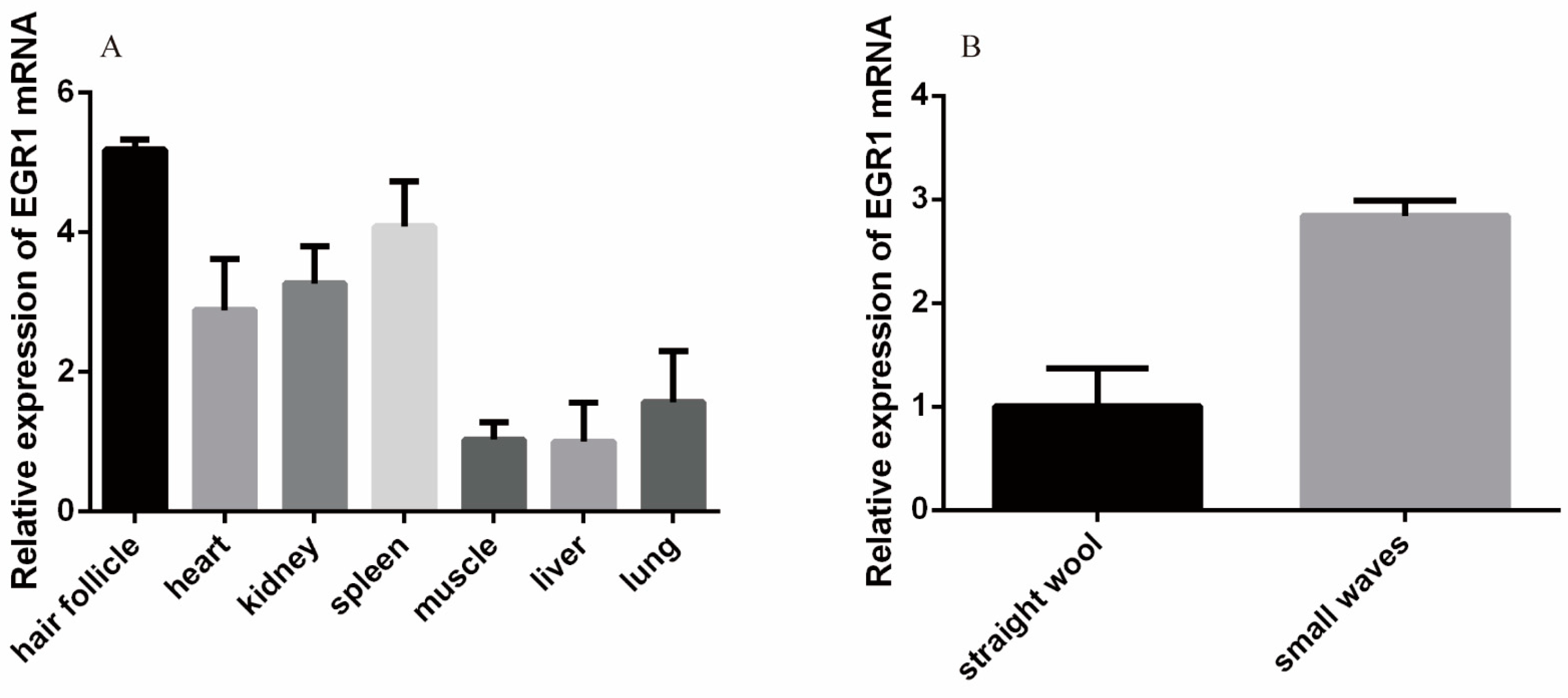
2.9. Analysis of EGR1 Expression Profiles in Different Tissues
2.10. Statistical Analysis
3. Results
3.1. Cloning of the EGR1 CDS Region
3.2. Bioinformatics Analysis of EGR1
3.3. Analysis of EGR1 Expression Profiles in Different Tissues
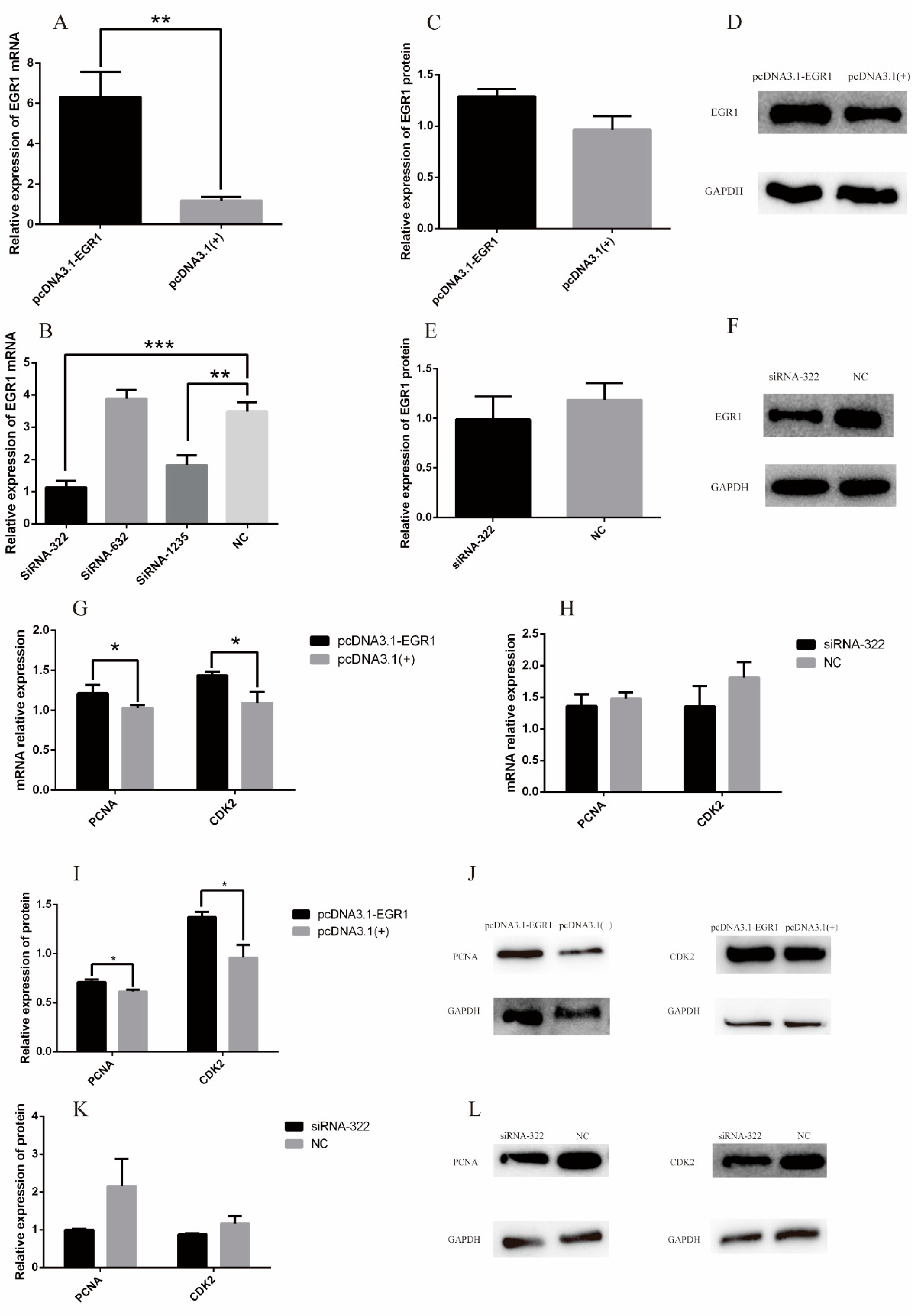
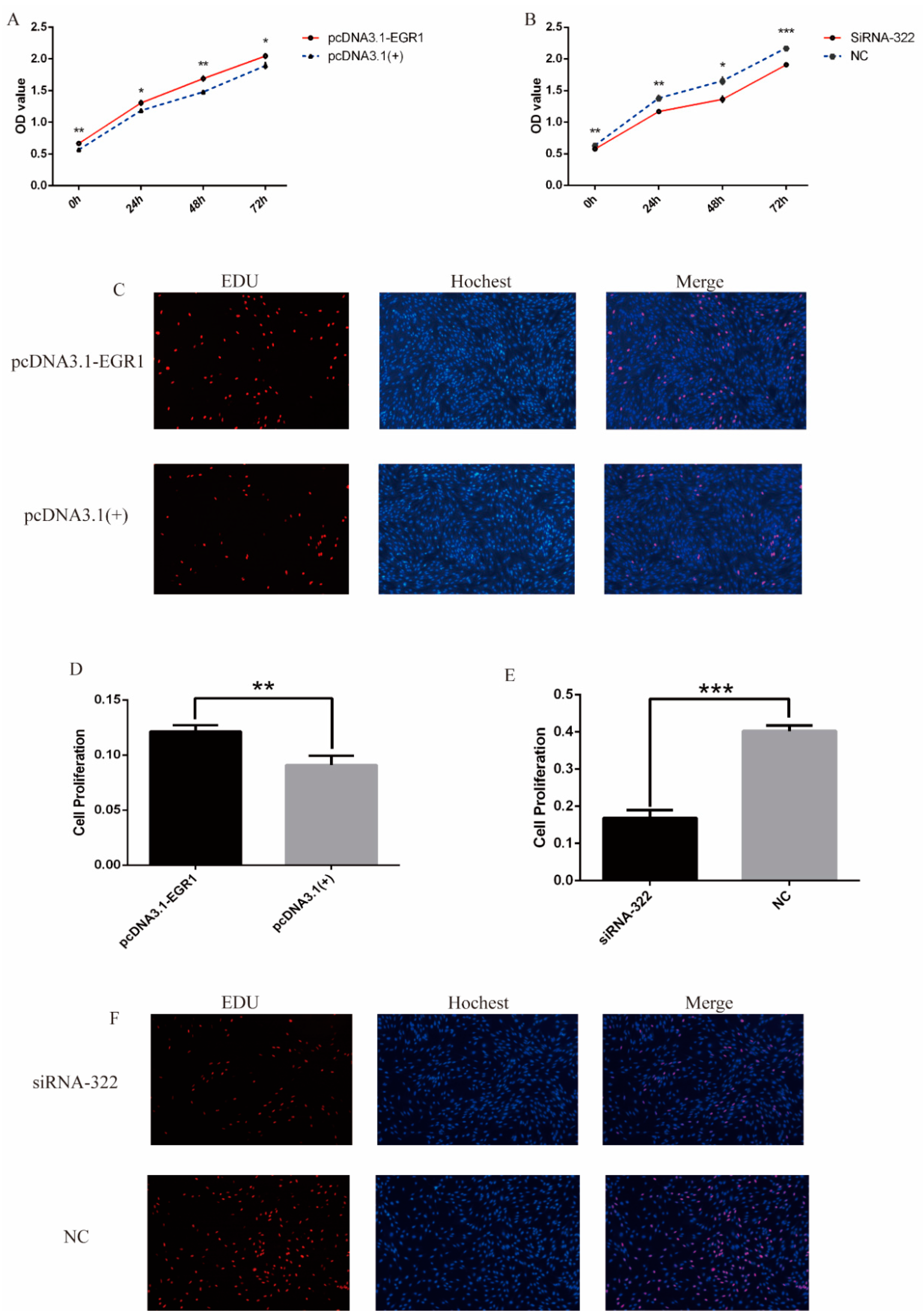
3.4. EGR1 Promoted Proliferation of DPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lv, X.; Chen, W.; Sun, W.; Hussain, Z.; Chen, L.; Wang, S.; Wang, J. Expression profile analysis to identify circular RNA expression signatures in hair follicle of Hu sheep lambskin. Genomics 2020, 112, 4454–4462. [Google Scholar] [CrossRef] [PubMed]
- Hardy, M.H. The secret life of the hair follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef]
- Botchkarev, V.A. Bone morphogenetic proteins and their antagonists in skin and hair follicle biology. J. Investig. Dermatol. 2003, 120, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Reddy, S.; Andl, T.; Bagasra, A.; Lu, M.M.; Epstein, D.J.; Morrisey, E.E.; Millar, S.E. Characterization of Wnt gene expression in developing and postnatal hair follicles and identification of Wnt5a as a target of Sonic hedgehog in hair follicle morphogenesis. Mech. Develop. 2001, 107, 69–82. [Google Scholar] [CrossRef]
- Nissimov, J.N.; Das Chaudhuri, A.B. Hair curvature: A natural dialectic and review. Biol. Rev. Camb. Philos. Soc. 2014, 89, 723–766. [Google Scholar] [CrossRef]
- Thiel, G.; Cibelli, G. Regulation of life and death by the zinc finger transcription factor Egr-1. J. Cell Physiol. 2002, 193, 287–292. [Google Scholar] [CrossRef]
- Unoki, M.; Nakamura, Y. EGR2 induces apoptosis in various cancer cell lines by direct transactivation of BNIP3L and BAK. Oncogene 2003, 22, 2172–2185. [Google Scholar] [CrossRef] [Green Version]
- Vaish, V.; Piplani, H.; Rana, C.; Vaiphei, K.; Sanyal, S.N. NSAIDs may regulate EGR-1-mediated induction of reactive oxygen species and non-steroidal anti-inflammatory drug-induced gene (NAG)-1 to initiate intrinsic pathway of apoptosis for the chemoprevention of colorectal cancer. Mol. Cell Biochem. 2013, 378, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jeong, I.Y.; Lim, Y.; Lee, Y.H.; Shin, S.Y. Estrogen receptor beta stimulates Egr-1 transcription via MEK1/Erk/Elk-1 cascade in C6 glioma cells. BMB Rep. 2011, 44, 452–457. [Google Scholar] [CrossRef]
- Li, L.; Ameri, A.H.; Wang, S.; Jansson, K.H.; Casey, O.M.; Yang, Q.; Beshiri, M.L.; Fang, L.; Lake, R.G.; Agarwal, S.; et al. EGR1 regulates angiogenic and osteoclastogenic factors in prostate cancer and promotes metastasis. Oncogene 2019, 38, 6241–6255. [Google Scholar] [CrossRef] [PubMed]
- Myung, E.; Park, Y.L.; Kim, N.; Chung, C.Y.; Park, H.B.; Park, H.C.; Myung, D.S.; Kim, J.S.; Cho, S.B.; Lee, W.S.; et al. Expression of early growth response-1 in human gastric cancer and its relationship with tumor cell behaviors and prognosis. Pathol. Res. Pract. 2013, 209, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Calogero, A.; Arcella, A.; De Gregorio, G.; Porcellini, A.; Mercola, D.; Liu, C.T.; Lombari, V.; Zani, M.; Giannini, G.; Gagliardi, F.M.; et al. The early growth response gene EGR-1 behaves as a suppressor gene that is down-regulated independent of ARF/Mdm2 but not p53 alterations in fresh human gliomas. Clin. Cancer Res. 2001, 7, 2788–2796. [Google Scholar] [PubMed]
- Kumbrink, J.; Gerlinger, M.; Johnson, J.P. Egr-1 induces the expression of its corepressor nab2 by activation of the nab2 promoter thereby establishing a negative feedback loop. J. Biol. Chem. 2005, 280, 42785–42793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, X.; Sun, W.; Zou, S.; Chen, L.; Mwacharo, J.M.; Wang, J. Characteristics of the BMP7 Promoter in Hu Sheep. Animals 2019, 9, 874. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Gao, W.; Jin, C.; Wang, L.; Wang, Y.; Chen, W.; Zou, S.; Huang, S.; Li, Z.; Wang, J.; et al. Preliminary study on microR-148a and microR-10a in dermal papilla cells of Hu sheep. BMC Genet. 2019, 20, 70. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Reynolds, A.J.; Lawrence, C.; Cserhalmi-Friedman, P.B.; Christiano, A.M.; Jahoda, C.A.B. Trans-gender induction of hair follicles. Nature 1999, 402, 33–34. [Google Scholar] [CrossRef]
- Driskell, R.R.; Giangreco, A.; Jensen, K.B.; Mulder, K.W.; Watt, F.M. Sox2-positive dermal papilla cells specify hair follicle type in mammalian epidermis. Development 2009, 136, 2815–2823. [Google Scholar] [CrossRef] [Green Version]
- Chi, W.; Wu, E.; Morgan, B.A. Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline. Development 2013, 140, 1676–1683. [Google Scholar] [CrossRef] [Green Version]
- Alonso, L.; Fuchs, E. The hair cycle. J. Cell Sci. 2006, 119, 391–393. [Google Scholar] [CrossRef]
- Jahoda, C.A.B.; Oliver, R.F.; Reynolds, A.J.; Forrester, J.C.; Gillespie, J.W.; Cserhalmi-Friedman, P.B.; Christiano, A.M.; Horne, K.A. Trans-species hair growth induction by human hair follicle dermal papillae. Exp. Dermatol. 2001, 10, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Clavel, C.; Rendl, M.; Watt, F.M. Hair follicle dermal papilla cells at a glance. J. Cell Sci. 2011, 124, 1179–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierard-Franchimont, C.; Paquet, P.; Quatresooz, P.; Pierard, G.E. Mechanobiology and cell tensegrity: The root of ethnic hair curling? J. Cosmet. Dermatol. 2011, 10, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.K.; Islam, S.M.R.; Saha, T.; Nishat, A.; Biswas, P.K.; Gil, M.; Nkenyereye, L.; El-Sappagh, S.; Islam, M.S.; Cho, S.G. Prognostic role of EGR1 in breast cancer: A systematic review. BMB Rep. 2021, 54, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Tallafuss, A.; Stednitz, S.J.; Voeun, M.; Levichev, A.; Larsch, J.; Eisen, J.; Washbourne, P. Egr1 Is Necessary for Forebrain Dopaminergic Signaling during Social Behavior. Eneuro 2022, 9, ENEURO.0035-22.2022. [Google Scholar] [CrossRef]
- Sheng, S.Y.; Zou, M.N.; Yang, Y.L.; Guan, M.P.; Ren, S.J.; Wang, X.Y.; Wang, L.; Xue, Y.M. miR-23a-3p regulates the inflammatory response and fibrosis in diabetic kidney disease by targeting early growth response 1. In Vitro Cell Dev. Biol.-Anim. 2021, 57, 763–774. [Google Scholar] [CrossRef]
- Patwardhan, S.; Gashler, A.; Siegel, M.G.; Chang, L.C.; Joseph, L.J.; Shows, T.B.; Le Beau, M.M.; Sukhatme, V.P. EGR3, a novel member of the Egr family of genes encoding immediate-early transcription factors. Oncogene 1991, 6, 917–928. [Google Scholar]
- Gupta, M.P.; Gupta, M.; Zak, R.; Sukhatme, V.P. Egr-1, a serum-inducible zinc finger protein, regulates transcription of the rat cardiac alpha-myosin heavy chain gene. J. Biol. Chem. 1991, 266, 12813–12816. [Google Scholar] [CrossRef]
- Lee, S.L.; Tourtellotte, L.C.; Wesselschmidt, R.L.; Milbrandt, J. Growth and differentiation proceeds normally in cells deficient in the immediate early gene NGFI-A. J. Biol. Chem. 1995, 270, 9971–9977. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, A.M.; Wilce, P.A. Egr transcription factors in the nervous system. Neurochem. Int. 1997, 31, 477–510. [Google Scholar] [CrossRef]
- Topilko, P.; Schneider-Maunoury, S.; Levi, G.; Trembleau, A.; Gourdji, D.; Driancourt, M.A.; Rao, C.V.; Charnay, P. Multiple pituitary and ovarian defects in Krox-24 (NGFI-A, Egr-1)-targeted mice. Mol. Endocrinol. 1998, 12, 107–122. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Sun, H.F.; Wang, X.F.; Hou, N.; Zhao, L.Y.; Tong, D.D.; He, K.; Yang, Y.; Song, T.S.; Yang, J.; et al. EGR1 mediates miR-203a suppress the hepatocellular carcinoma cells progression by targeting HOXD3 through EGFR signaling pathway. Oncotarget 2016, 7, 45302–45316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, X.; He, L.; Wang, R.; Shen, J.; Wang, Z.; Shen, Y.; Fan, L.; Shao, J.; Qi, H. miR-377-3p-Mediated EGR1 Downregulation Promotes B[a]P-Induced Lung Tumorigenesis by Wnt/Beta-Catenin Transduction. Front. Oncol. 2021, 11, 699004. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.Y.; Liu, K.T.; Xiang, Z.F.; Yu, X.; Wang, P.; Ma, Q. MiR-125b-2-3p associates with prognosis of ccRCC through promoting tumor metastasis via targeting EGR1. Am. J. Transl. Res. 2020, 12, 5575–5585. [Google Scholar]
- Yan, L.; Wang, Y.; Liang, J.; Liu, Z.; Sun, X.; Cai, K. MiR-301b promotes the proliferation, mobility, and epithelial-to-mesenchymal transition of bladder cancer cells by targeting EGR1. Biochem. Cell Biol. 2017, 95, 571–577. [Google Scholar] [CrossRef]
- Adly, M.A.; Assaf, H.A.; Hussein, M.R. Expression of bone morphogenetic protein-7 in human scalp skin and hair follicles. Br. J. Dermatol. 2006, 154, 551–554. [Google Scholar] [CrossRef]
- Schwachtgen, J.L.; Houston, P.; Campbell, C.; Sukhatme, V.; Braddock, M. Fluid shear stress activation of egr-1 transcription in cultured human endothelial and epithelial cells is mediated via the extracellular signal-related kinase 1/2 mitogen-activated protein kinase pathway. J. Clin. Investig. 1998, 101, 2540–2549. [Google Scholar] [CrossRef] [Green Version]
- Burnatowska-Hledin, M.A.; Kossoris, J.B.; Van Dort, C.J.; Shearer, R.L.; Zhao, P.; Murrey, D.A.; Abbott, J.L.; Kan, C.E.; Barney, C.C. T47D breast cancer cell growth is inhibited by expression of VACM-1, a cul-5 gene. Biochem. Bioph. Res. 2004, 319, 817–825. [Google Scholar] [CrossRef]
- Lv, X.Y.; Sun, W.; Yin, J.F.; Ni, R.; Su, R.; Wang, Q.Z.; Gao, W.; Bao, J.J.; Yu, J.R.; Wang, L.H.; et al. An Integrated Analysis of MicroRNA and mRNA Expression Profiles to Identify RNA Expression Signatures in Lambskin Hair Follicles in Hu Sheep. PLoS ONE 2016, 11, e0157463. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′→3′) |
|---|---|
| siRNA-322 | CCUGACAUCUCUCUGAAUATT UAUUCAGAGAGAUGUCAGGTT |
| siRNA-632 | CCACACCUAACACUGACAUTT AUGUCAGUGUUAGGUGUGGTT |
| siRNA-1235 | GCAAGAGGCAUACCAAGAUTT AUCUUGGUAUGCCUCUUGCTT |
| Gene ID | Sequences (5′→30′) | Product Length/bp | Accession No. |
|---|---|---|---|
| EGR1 | F: TTCAACCCTCAGGGGGAGG R: CGCTGACCAGACTGAAGAGG | 223 | EU552504.1 |
| PCNA | F: TCTGCAAGTGGAGAACTTGGAA R: AGGAGACAGTGGAGTGGCTT | 162 | XM_004014340.5 |
| CDK2 | F: TGGGCCAGGCAGGATTTTAG R: GTCGAAGGTGAGGTACTGGC | 166 | FJ422550.1 |
| GAPDH | F: TCTCAAGGGCATTCTAGGCTAC R: GCCGAATTCATTGTCGTACCAG | 151 | NM_001190390.1 |
| Species | GenBank No. | Nucleotide (%) | Amino Acid (%) |
|---|---|---|---|
| Bos taurus | AY924307.1 | 97.06 | 98.53 |
| Homo sapiens | MK681487.1 | 92.03 | 95.88 |
| Rattus norvegicus | AY551092.1 | 84.85 | 85.69 |
| Mus musculus | BC138615.1 | 84.70 | 86.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Wang, S.; Cao, X.; Yuan, Z.; Getachew, T.; Mwacharo, J.M.; Haile, A.; Lv, X.; Sun, W. The Effect of EGR1 on the Proliferation of Dermal Papilla Cells. Genes 2022, 13, 1242. https://doi.org/10.3390/genes13071242
Xu Y, Wang S, Cao X, Yuan Z, Getachew T, Mwacharo JM, Haile A, Lv X, Sun W. The Effect of EGR1 on the Proliferation of Dermal Papilla Cells. Genes. 2022; 13(7):1242. https://doi.org/10.3390/genes13071242
Chicago/Turabian StyleXu, Yeling, Shanhe Wang, Xiukai Cao, Zehu Yuan, Tesfaye Getachew, Joram M. Mwacharo, Aynalem Haile, Xiaoyang Lv, and Wei Sun. 2022. "The Effect of EGR1 on the Proliferation of Dermal Papilla Cells" Genes 13, no. 7: 1242. https://doi.org/10.3390/genes13071242