
Toxoplasma gondii Seropositivity Interacts with Catechol-O-methyltransferase Val105/158Met Variation Increasing the Risk of Schizophrenia

 , , ,
, , ,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample and Measures
2.2. Microbiological Analyses
2.2.1. Detection of IgG Anti-T. gondii by ELISA
2.2.2. Detection of T. gondii DNA by Nested PCR
2.3. Genetic Analyses
2.4. Statistical Analyses
3. Results
3.1. Sociodemographic Sample Characteristics
3.2. Main Effect of the COMT Val105/158Met Polymorphism
3.3. Main Effect of T. gondii Infection
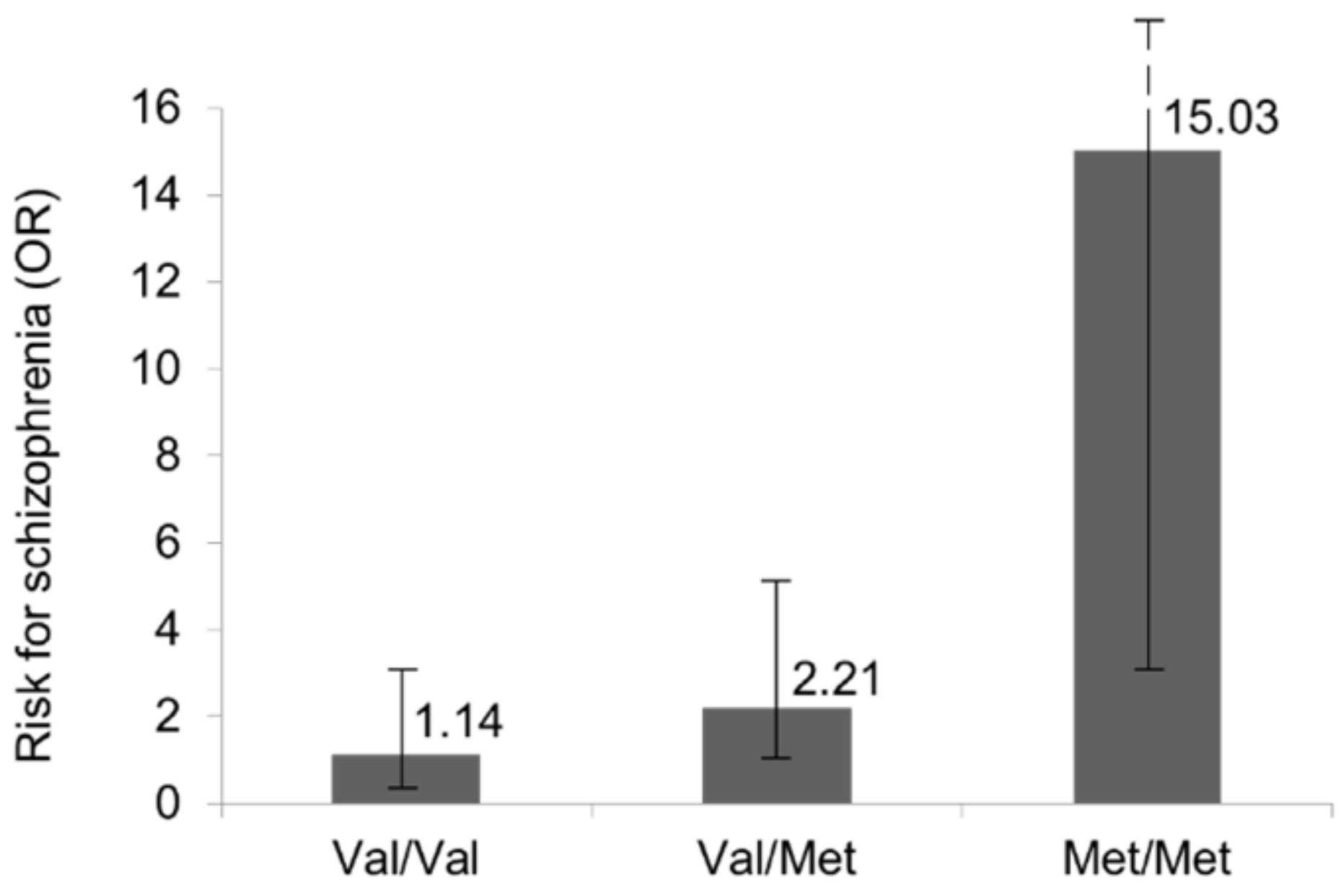
3.4. COMT–T. gondii Interaction
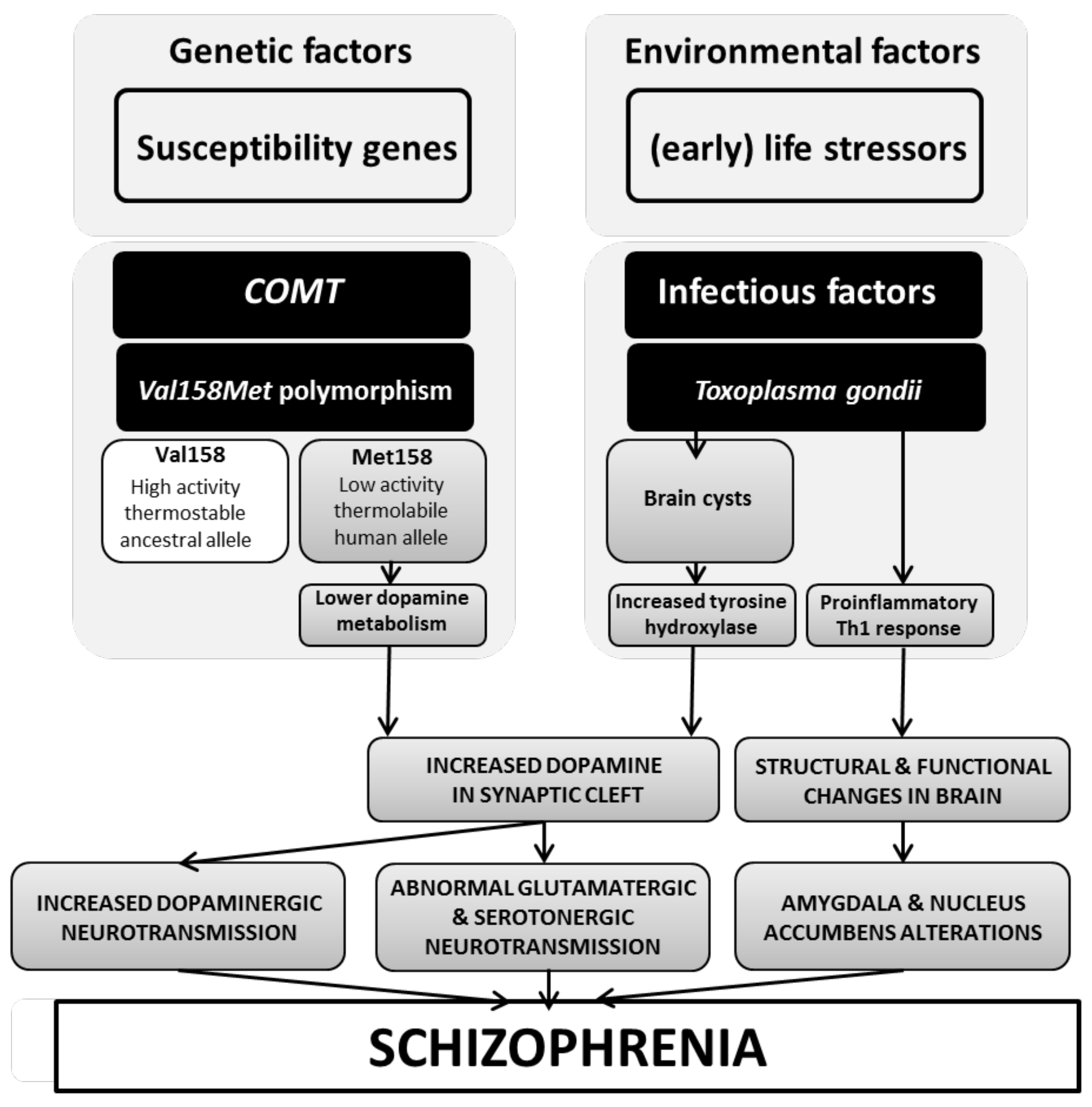
4. Discussion
Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- European Network of National Networks Studying Gene-Environment Interactions in Schizophrenia (EU-GEI); van Os, J.; Rutten, B.P.; Myin-Germeys, I.; Delespaul, P.; Viechtbauer, W.; van Zelst, C.; Bruggeman, R.; Reininghaus, U.; Morgan, C.; et al. Identifying gene-environment interactions in schizophrenia: Contemporary challenges for integrated, large-scale investigations. Schizophr. Bull. 2014, 40, 729–736. [Google Scholar] [PubMed] [Green Version]
- Howes, O.D.; Kapur, S. The dopamine hypothesis of schizophrenia: Version III--the final common pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubetskoy, V.; Pardiñas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.-Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Hosák, L. Role of the COMT gene Val105/158Met polymorphism in mental disorders: A review. Eur. Psychiatry J. Assoc. Eur. Psychiatr. 2007, 22, 276–281. [Google Scholar] [CrossRef]
- Arias, I.; Sorlozano, A.; Villegas, E.; Luna, J.D.D.; McKenney, K.; Cervilla, J.; Gutierrez, B.; Gutierrez, J. Infectious agents associated with schizophrenia: A meta-analysis. Schizophr. Res. 2012, 136, 128–136. [Google Scholar] [CrossRef]
- Shehata, A.I.; Hassanein, F.I.; Abdul-Ghani, R. Seroprevalence of Toxoplasma gondii infection among patients with non-schizophrenic neurodevelopmental disorders in Alexandria, Egypt. Acta Trop. 2016, 154, 155–159. [Google Scholar] [CrossRef]
- Stilo, S.A.; Murray, R.M. Non-Genetic Factors in Schizophrenia. Curr. Psychiatry Rep. 2019, 21, 100. [Google Scholar] [CrossRef] [Green Version]
- Wahbeh, M.H.; Avramopoulos, D. Gene-Environment Interactions in Schizophrenia: A Literature Review. Genes 2021, 12, 1850. [Google Scholar] [CrossRef]
- Boksa, P. Maternal infection during pregnancy and schizophrenia. J. Psychiatry Neurosci. JPN 2008, 33, 183–185. [Google Scholar]
- Torrey, E.F.; Yolken, R.H. Toxoplasma gondii and schizophrenia. Emerg. Infect. Dis. 2003, 9, 1375–1380. [Google Scholar] [CrossRef]
- Oncu-Oner, T.; Can, S. Meta-analysis of the relationship between Toxoplasma gondii and schizophrenia. Ann. Parasitol. 2022, 68, 103–110. [Google Scholar]
- Prandovszky, E.; Gaskell, E.; Martin, H.; Dubey, J.P.; Webster, J.P.; McConkey, G.A. The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS ONE. 2011, 6, e23866. [Google Scholar] [CrossRef]
- Wing, J.K.; Babor, T.; Brugha, T.; Burke, J.; Cooper, J.E.; Giel, R.; Jablenski, A.; Regier, D.; Sartorius, N. SCAN. Schedules for Clinical Assessment in Neuropsychiatry. Arch. Gen. Psychiatry 1990, 47, 589–593. [Google Scholar] [CrossRef]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Balker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59 (Suppl. 20), 22–33; [Google Scholar]
- Daniels, J.K.; Williams, N.M.; Williams, J.; Jones, L.A.; Cardno, A.G.; Murphy, K.C.; Spurlock, G.; Riley, B.; Scambler, P.; Asherson, P.; et al. No evidence for allelic association between schizophrenia and a polymorphism determining high or low catechol O-methyltransferase activity. Am. J. Psychiatry 1996, 153, 268–270. [Google Scholar]
- Gutiérrez, B.; Bertranpetit, J.; Guillamat, R.; Vallès, V.; Arranz, M.J.; Kerwin, R.; Fañanás, L. Association analysis of the catechol O-methyltransferase gene and bipolar affective disorder. Am. J. Psychiatry 1997, 154, 113–115. [Google Scholar]
- Bilder, R.M.; Volavka, J.; Lachman, H.M.; Grace, A.A. The Catechol-O-Methyltransferase Polymorphism: Relations to the Tonic–Phasic Dopamine Hypothesis and Neuropsychiatric Phenotypes. Neuropsychopharmacology 2004, 29, 1943–1961. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.M.; Pilowsky, L.S. Dopamine and antipsychotic drug action revisited. Br. J. Psychiatry J. Ment. Sci. 2002, 181, 271–275. [Google Scholar] [CrossRef] [Green Version]
- Rosa, A.; Peralta, V.; Cuesta, M.J.; Zarzuela, A.; Serrano, F.; Martínez-Larrea, A.; Fañanás, L. New evidence of association between COMT gene and prefrontal neurocognitive function in healthy individuals from sibling pairs discordant for psychosis. Am. J. Psychiatry 2004, 161, 1110–1112. [Google Scholar] [CrossRef]
- Munafò, M.R.; Bowes, L.; Clark, T.G.; Flint, J. Lack of association of the COMT (Val158Met) gene and schizophrenia: A meta-analysis of case-control studies. Mol. Psychiatry 2005, 10, 765–770. [Google Scholar] [CrossRef]
- Réthelyi, J.M.; Benkovits, J.; Bitter, I. Genes and environments in schizophrenia: The different pieces of a manifold puzzle. Neurosci. Biobehav. Rev. 2013, 37 Pt 1, 2424–2437. [Google Scholar] [CrossRef]
- Yolken, R.H.; Dickerson, F.B.; Fuller Torrey, E. Toxoplasma and schizophrenia. Parasite Immunol. 2009, 31, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Fernández, J.; Luna Del Castillo, J.d.D.; Mañanes-González, S.; Carrillo-Ávila, J.A.; Gutiérrez, B.; Cervilla, J.A.; Sorlozano-Puerto, A. Different presence of Chlamydia pneumoniae, herpes simplex virus type 1, human herpes virus 6, and Toxoplasma gondii in schizophrenia: Meta-analysis and analytical study. Neuropsychiatrc Dis. Treat. 2015, 11, 843–852. [Google Scholar] [CrossRef] [Green Version]
- Bird, E.D.; Spokes, E.G.; Iversen, L.L. Increased dopamine concentration in limbic areas of brain from patients dying with schizophrenia. Brain J. Neurol. 1979, 102, 347–360. [Google Scholar] [CrossRef]
- Weinberger, D.R. Implications of normal brain development for the pathogenesis of schizophrenia. Arch. Gen. Psychiatry 1987, 44, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Webster, J.P. Rats, cats, people and parasites: The impact of latent toxoplasmosis on behaviour. Microbes Infect. 2001, 3, 1037–1045. [Google Scholar] [CrossRef]
- Xiao, J.; Prandovszky, E.; Kannan, G.; Pletnikov, M.V.; Dickerson, F.; Severance, E.G.; Yolken, R.H. Toxoplasma gondii: Biological Parameters of the Connection to Schizophrenia. Schizophr. Bull. 2018, 44, 983–992. [Google Scholar] [CrossRef] [Green Version]
- Schizophrenia Working Group of the Psychiatric Genomics Consortium. Biological insights from 108 schizophrenia-associated genetic loci. Nature 2014, 511, 421–427. [Google Scholar] [CrossRef] [Green Version]
- Avramopoulos, D.; Pearce, B.D.; McGrath, J.; Wolyniec, P.; Wang, R.; Eckart, N.; Hatzimanolis, A.; Goes, F.S.; Nestadt, G.; Mulle, J.; et al. Infection and Inflammation in Schizophrenia and Bipolar Disorder: A Genome Wide Study for Interactions with Genetic Variation. Potash JB, editor. PLoS ONE. 2015, 10, e0116696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Mouhawess, A.; Hammoud, A.; Zoghbi, M.; Hallit, S.; Haddad, C.; El Haddad, K.; el Khoury, S.; Tannous, J.; Obeid, S.; Halabi, M.A.; et al. Relationship between Toxoplasma gondii seropositivity and schizophrenia in the Lebanese population: Potential implication of genetic polymorphism of MMP-9. BMC Psychiatry 2020, 20, 264. [Google Scholar]
- Horacek, J.; Flegr, J.; Tintera, J.; Verebova, K.; Spaniel, F.; Novak, T.; Brunovsky, M.; Bubenikova-Valesova, V.; Holub, D.; Palenicek, T.; et al. Latent toxoplasmosis reduces gray matter density in schizophrenia but not in controls: Voxel-based-morphometry (VBM) study. World J. Biol. Psychiatry 2012, 13, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Mamah, D.; Wang, L.; Barch, D.; de Erausquin, G.A.; Gado, M.; Csernansky, J.G. Structural analysis of the basal ganglia in schizophrenia. Schizophr. Res. 2007, 89, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Shergill, S.S.; Bullmore, E.; Simmons, A.; Murray, R.; McGuire, P. Functional anatomy of auditory verbal imagery in schizophrenic patients with auditory hallucinations. Am. J. Psychiatry 2000, 157, 1691–1693. [Google Scholar] [CrossRef]
- Lawrie, S.M.; Whalley, H.C.; Job, D.E.; Johnstone, E.C. Structural and functional abnormalities of the amygdala in schizophrenia. Ann. N. Y. Acad. Sci. 2003, 985, 445–460. [Google Scholar] [CrossRef]
- Lotta, T.; Vidgren, J.; Tilgmann, C.; Ulmanen, I.; Melén, K.; Julkunen, I.; Taskinen, J. Kinetics of human soluble and membrane-bound catechol O-methyltransferase: A revised mechanism and description of the thermolabile variant of the enzyme. Biochemistry 1995, 34, 4202–4210. [Google Scholar] [CrossRef]
- Torrey, E.F.; Yolken, R.H. Schizophrenia as a pseudogenetic disease: A call for more gene-environmental studies. Psychiatry Res. 2019, 278, 146–150. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristics | Patients N (%) | Controls N (%) | p-Value |
|---|---|---|---|
| Mean age (n = 283) | 33.29 (SD = 8.58) | 39.08 (SD = 10.83) | t = 4.94, p-value < 0.001 |
| Sex (n = 283) | |||
| Male | 95 (67.4) | 67 (47.2) | χ2 = 12.76, df = 1, p-value < 0.001 |
| Female | 46 (32.6) | 75 (52.8) | |
| Education (n = 271) | |||
| Elementary education | 21 (16.3) | 15 (10.6) | χ2 = 7.33, df = 3, p-value < 0.062 |
| Junior school | 59 (45.7) | 60 (42.3) | |
| High school | 44 (34.1) | 50 (35.2) | |
| University degree | 5 (3.9) | 17 (12) | |
| Marital status (n = 270) | |||
| Single/never married | 97 (75.2) | 41 (29.1) | χ2 = 61.03, df = 3, p-value < 0.001 |
| Living with a partner | 21 (16.3) | 84 (59.6) | |
| Divorced/separated | 11 (8.5) | 15 (10.6) | |
| Widowed | 0 (0) | 1 (0.7) |
| Allelic and Genotypic Frequencies | Controls N (%) | Patients N (%) |
|---|---|---|
| Allelic frequencies | ||
| G | 167 (58.80) | 155 (54.96) |
| A | 117 (41.20) | 127 (45.04) |
| χ2 = 0.85; df = 1; p-value = 0.35 | ||
| Genotypic frequencies | ||
| G/G | 46 (32.4) | 40 (28.4) |
| G/A | 75 (52.8) | 75 (53.2) |
| A/A | 21 (14.8) | 26 (18.4) |
| χ2 = 0.95; df = 2; p-value = 0.62 | ||
| Infection by T. gondii | Controls (N (%)) | Patients (N (%)) |
|---|---|---|
| Yes | 29 (20.42) | 55 (39) |
| No | 113 (79.58) | 86 (61) |
| OR = 2.50; 95% CI: 1.47–4.23; p-value = 0.0006 | ||
| Infection by T. gondii | Group | Val/Val | Val/Met | Met/Met |
|---|---|---|---|---|
| No | Controls | 35 (31%) | 59 (52.2%) | 19 (16.8%) |
| Cases | 30 (34.9%) | 46 (53.5%) | 10 (11.6%) | |
| Yes | Controls | 11 (37.9%) | 16 (55.2%) | 2 (6.9%) |
| Cases | 10 (18.2%) | 29 (52.7%) | 16 (29.1%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rovira, P.; Gutiérrez, B.; Sorlózano-Puerto, A.; Gutiérrez-Fernández, J.; Molina, E.; Rivera, M.; Martínez-Leal, R.; Ibanez-Casas, I.; Martín-Laguna, M.V.; Rosa, A.; et al. Toxoplasma gondii Seropositivity Interacts with Catechol-O-methyltransferase Val105/158Met Variation Increasing the Risk of Schizophrenia. Genes 2022, 13, 1088. https://doi.org/10.3390/genes13061088
Rovira P, Gutiérrez B, Sorlózano-Puerto A, Gutiérrez-Fernández J, Molina E, Rivera M, Martínez-Leal R, Ibanez-Casas I, Martín-Laguna MV, Rosa A, et al. Toxoplasma gondii Seropositivity Interacts with Catechol-O-methyltransferase Val105/158Met Variation Increasing the Risk of Schizophrenia. Genes. 2022; 13(6):1088. https://doi.org/10.3390/genes13061088
Chicago/Turabian StyleRovira, Paula, Blanca Gutiérrez, Antonio Sorlózano-Puerto, José Gutiérrez-Fernández, Esther Molina, Margarita Rivera, Rafael Martínez-Leal, Inmaculada Ibanez-Casas, María Victoria Martín-Laguna, Araceli Rosa, and et al. 2022. "Toxoplasma gondii Seropositivity Interacts with Catechol-O-methyltransferase Val105/158Met Variation Increasing the Risk of Schizophrenia" Genes 13, no. 6: 1088. https://doi.org/10.3390/genes13061088