
Identification of the Core Promoter and Variants Regulating Chicken CCKAR Expression



Abstract
:1. Introduction
2. Materials and Methods
2.1. Re-Sequencing the CCKAR Locus
2.2. Identification of Conserved Element Variants
2.3. Construction of Reporter Gene Plasmids
2.4. Cell Culture, Transfection and Dual-Luciferase Reporter Assay
2.5. Predicting Disrupting Effects of Variants on TF Binding Motifs
2.6. Statistical Analysis
3. Results
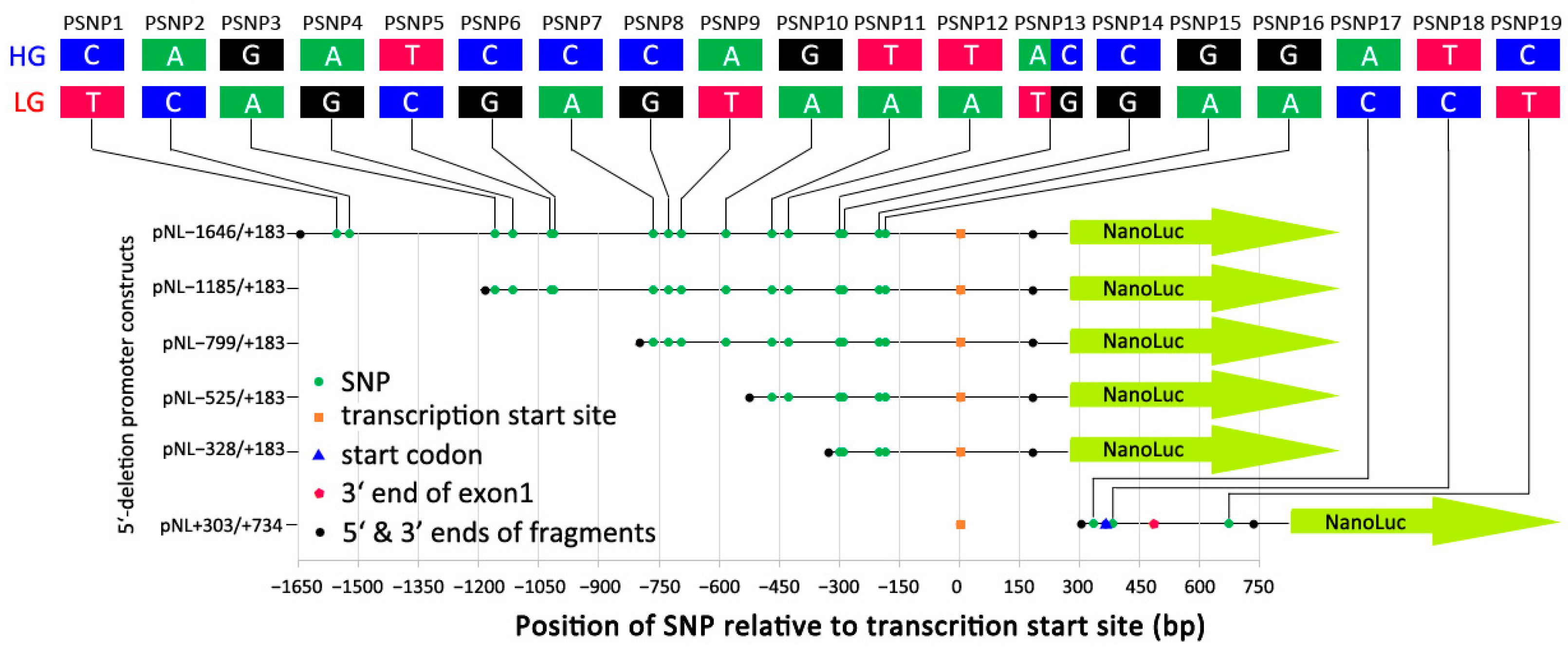
3.1. Identification of Variants in the 21 kb Region Flanking CCKAR
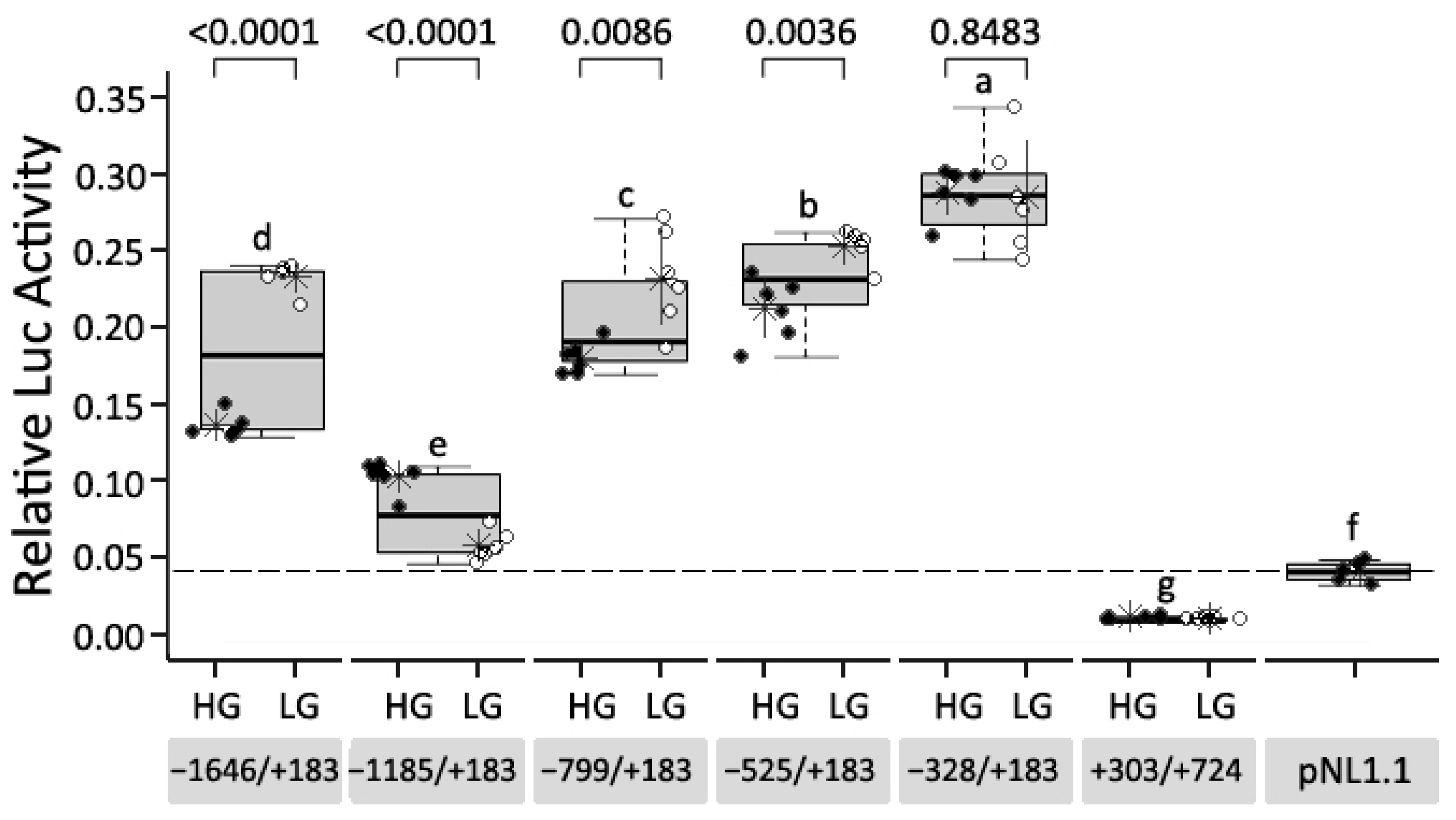
3.2. Defining the Core Promoter of Chicken CCKAR
3.3. Effects of Sequence Variants in the Promoter Region of CCKAR on Promoter Activity
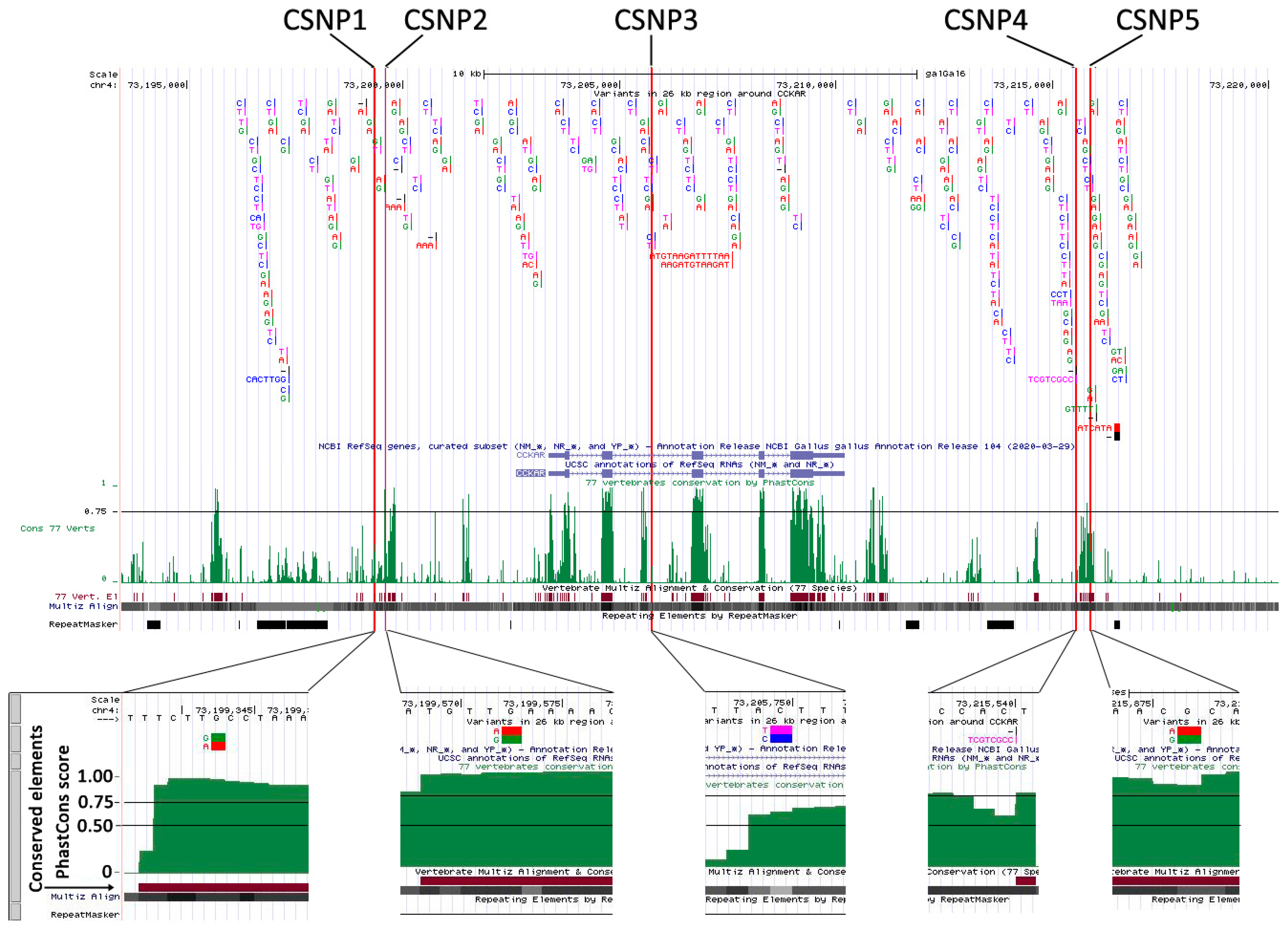
3.4. Discovery of Conserved Sequence Variants Affecting Promoter Activity
4. Discussion
4.1. Core Promoter of Chicken CCKAR Gene
4.2. Expression Regulation Effects of Sequence Variants in the Promoter Region of CCKAR
4.3. Effects of Conserved Element Variants on Promoter Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahima, R.S.; Antwi, D.A. Brain regulation of appetite and satiety. Endocrin. Metab. Clin. 2008, 37, 811–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynne, K.; Stanley, S.; McGowan, B.; Bloom, S. Appetite control. J. Endocrinol. 2005, 184, 291–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wren, A.M.; Bloom, S.R. Gut hormones and appetite control. Gastroenterology 2007, 132, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Austin, J.; Marks, D. Hormonal regulators of appetite. Int. J. Pediatr. Endocrinol. 2009, 2009, 141753. [Google Scholar] [CrossRef] [PubMed]
- Owyang, C.; Heldsinger, A. Vagal control of satiety and hormonal regulation of appetite. J. Neurogastroenterol. Motil. 2011, 17, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seroussi, E.; Cinnamon, Y.; Yosefi, S.; Genin, O.; Smith, J.G.; Rafati, N.; Bornelöv, S.; Andersson, L.; Friedman-Einat, M. Identification of the Long-Sought Leptin in Chicken and Duck, Expression Pattern of the Highly GC-Rich Avian leptin Fits an Autocrine/Paracrine Rather than Endocrine Function. Endocrinology 2016, 157, 737–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boswell, T.; Dunn, I.C. Regulation of Agouti-Related Protein and Pro-Opiomelanocortin Gene Expression in the Avian Arcuate Nucleus. Front. Endocrinol. 2017, 8, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seroussi, E.; Knytl, M.; Pitel, F.; Elleder, D.; Krylov, V.; Leroux, S.; Morisson, M.; Yosefi, S.; Miyara, S.; Ganesan, S.; et al. Avian Expression Patterns and Genomic Mapping Implicate Leptin in Digestion and TNF in Immunity; Suggesting That Their Interacting Adipokine Role Has Been Acquired Only in Mammals. Int. J. Mol. Sci. 2019, 20, 4489. [Google Scholar] [CrossRef] [Green Version]
- Guilloteau, P.; Le Meuth-Metzinger, V.; Morisset, J.; Zabielski, R. Gastrin; cholecystokinin and gastrointestinal tract functions in mammals. Nutr. Res. Rev. 2006, 19, 254–283. [Google Scholar] [CrossRef] [Green Version]
- Rehfeld, J.F. Cholecystokinin-From Local Gut Hormone to Ubiquitous Messenger. Front. Endocrinol. 2017, 8, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, T.H.; Bi, S. Hyperphagia and obesity in OLETF rats lacking CCK-1 receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, L.J.; Holicky, E.L.; Ulrich, C.D.; Wieben, E.D. Abnormal processing of the human cholecystokinin receptor gene in association with gallstones and obesity. Gastroenterology 1995, 109, 1375–1380. [Google Scholar] [CrossRef]
- Covasa, M.; Forbes, J.M. Effects of the CCK receptor antagonist MK-329 on food intake in broiler chickens. Pharmacol. Biochem. Behav. 1994, 48, 479–486. [Google Scholar] [CrossRef]
- Ebenezer, I.S.; de la Riva, C.; Baldwin, B.A. Effects of the CCK receptor antagonist MK-329 on food intake in pigs. Physiol. Behav. 1990, 47, 145–148. [Google Scholar] [CrossRef]
- Hewson, G.; Leighton, G.E.; Hill, R.G.; Hughes, J. The cholecystokinin receptor antagonist L364;718 increases food intake in the rat by attenuation of the action of endogenous cholecystokinin. Br. J. Pharmacol. 1988, 93, 79–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, I.C.; Meddle, S.L.; Wilson, P.W.; Wardle, C.A.; Law, A.S.; Bishop, V.R.; Hindar, C.; Robertson, G.W.; Burt, D.W.; Ellison, S.J.; et al. Decreased expression of the satiety signal receptor CCKAR is responsible for increased growth and body weight during the domestication of chickens. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E909–E921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rikimaru, K.; Takeda, H.; Uemoto, Y.; Komatsu, M.; Takahashi, D.; Suzuki, K.; Takahashi, H. Effect of a single-nucleotide polymorphism in the cholecystokinin type A receptor gene on growth traits in the Hinai-dori chicken breed. J. Poult. Sci. 2013, 50, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Li, X.; Luo, W.; Xu, Z.; Ji, C.; Zhang, Y.; Nie, Q.; Zhang, D.; Zhang, X. Feed conversion ratio; residual feed intake and cholecystokinin type A receptor gene polymorphisms are associated with feed intake and average daily gain in a Chinese local chicken population. J. Anim. Sci. Biotechnol. 2018, 9, 50. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, W. Research Note, Fine mapping of sequence variants associated with body weight of Lueyang black-boned chicken in the CCKAR gene. Poult. Sci. 2021, 100, 101448. [Google Scholar] [CrossRef] [PubMed]
- Blanchette, M.; Kent, W.J.; Riemer, C.; Elnitski, L.; Smit, A.F.; Roskin, K.M.; Baertsch, R.; Rosenbloom, K.; Clawson, H.; Green, E.D.; et al. Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res. 2004, 14, 708–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J.; Churchill, G.A. A Hidden Markov Model approach to variation among sites in rate of evolution. Mol. Biol. Evol. 1996, 13, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and inference of eukaryotic transcription factor sequence specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Berhanu Lemma, R.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Khan, N.M.; et al. JASPAR 2022, the 9th release of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef]
- Galbraith, J.D.; Kortschak, R.D.; Suh, A.; Adelson, D.L. Genome Stability Is in the Eye of the Beholder: CR1 Retrotransposon Activity Varies Significantly across Avian Diversity. Genome Biol. Evol. 2021, 13, evab259. [Google Scholar] [CrossRef]
- Takata, Y.; Takeda, S.; Kawanami, T.; Takiguchi, S.; Yoshida, Y.; Miyasaka, K.; Funakoshi, A. Promoter analysis of human cholecystokinin type-A receptor gene. J. Gastroenterol. 2002, 37, 815–820. [Google Scholar] [CrossRef]
- Thomas, B.E.; Thekkumkara, T.J. Glucose mediates transcriptional repression of the human angiotensin type-1 receptor gene: Role for a novel cis-acting element. Mol. Biol. Cell 2004, 15, 4347–4355. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; McFarland, K.A.; Jin, S.; Tong, G.; Duan, B.; Yang, A.; Hughes, T.R.; Liu, J.; Dove, S.L.; Navarre, W.W.; et al. A Novel AT-Rich DNA Recognition Mechanism for Bacterial Xenogeneic Silencer MvaT. PLoS Pathog. 2015, 11, e1004967. [Google Scholar] [CrossRef]
- Courey, A.J.; Jia, S. Transcriptional repression: The long and the short of it. Genes Dev. 2001, 15, 2786–2796. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Li, W.; Hou, N.; Huang, N. A Review of FoxO1-Regulated Metabolic Diseases and Related Drug Discoveries. Cells 2020, 9, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.; Ou, L.; Wang, W.; Guo, D.Y. Gastrin; Cholecystokinin; Signaling; and Biological Activities in Cellular Processes. Front. Endocrinol. 2020, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Siepel, A.; Bejerano, G.; Pedersen, J.S.; Hinrichs, A.S.; Hou, M.; Rosenbloom, K.; Clawson, H.; Spieth, J.; Hillier, L.W.; Richards, S.; et al. Evolutionarily conserved elements in vertebrate, insect, worm, and yeast genomes. Genome Res. 2005, 15, 1034–1050. [Google Scholar] [CrossRef] [Green Version]
- Kerje, S.; Carlborg, O.; Jacobsson, L.; Schütz, K.; Hartmann, C.; Jensen, P.; Andersson, L. The twofold difference in adult size between the red junglefowl and White Leghorn chickens is largely explained by a limited number of QTLs. Anim Genet. 2003, 34, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Rana, K.; Fam, B.C.; Clarke, M.V.; Pang, T.P.; Zajac, J.D.; MacLean, H.E. Increased adiposity in DNA binding-dependent androgen receptor knockout male mice associated with decreased voluntary activity and not insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E767–E778. [Google Scholar] [CrossRef]
- Miller, L.J.; Desai, A.J. Metabolic Actions of the Type 1 Cholecystokinin Receptor, Its Potential as a Therapeutic Target. Trends Endocrinol. Metab. 2016, 27, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Nakata, M.; Okamoto, S.; Takano, E.; Yada, T.; Minokoshi, Y.; Hirata, Y.; Nakajima, K.; Iskandar, K.; Hayashi, Y.; et al. PDK1-Foxo1 in agouti-related peptide neurons regulates energy homeostasis by modulating food intake and energy expenditure. PLoS ONE 2011, 6, e18324. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, K.; Cao, Y.; Hayashi, Y.; Nakata, M.; Takano, E.; Yada, T.; Zhang, C.; Ogawa, W.; Oki, M. PDK-1/FoxO1 pathway in POMC neurons regulates Pomc expression and food intake. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E787–E798. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | Positions Relative to CCKAR 1 | Primer Sequence (5′-3′) 2 | Size (bp) |
|---|---|---|---|
| Serial 5′-deleted promoter fragments | −1646/+183 | F: GGGGGTACCTAGAAAGGCAGGTATGTGCT R: GTTCACTCGAGTATGCCACACGAGGCAGTTT | 1829 |
| −1185/+1832 | F: GGGGGTACCAAGCCGAATTAGGCAGCTAA R: GTTCACTCGAGTATGCCACACGAGGCAGTTT | 1369 | |
| −799/+183 | F: GGGGGTACCGAAATCAGAACTTTTCAATC R: GTTCACTCGAGTATGCCACACGAGGCAGTTT | 983 | |
| −525/+183 | F: GGGGGTACCCTTTTAAACAGTCAAGGCTG R: GTTCACTCGAGTATGCCACACGAGGCAGTTT | 709 | |
| −328/+183 | F: GGGGGTACCGGCTTTGGATCAGGGACATG R: GTTCACTCGAGTATGCCACACGAGGCAGTTT | 512 | |
| Intragenic fragment | +303/+734 | F: GGGGGTACCATGGGCAGATAGTTACAAAC R: GTTCACTCGAGAAGCTGTTTACATTTTGTAC | 432 |
| CSNP-contained fragments | Upstream 4.0 kb (CSNP1) | F: GGGGGTACCCCTGCAGAGGTTCACTATGCT R:GTTCACTCGAGACCATTATTTGGGATGTTGGGA | 141 |
| Upstream 3.8 kb (CSNP2) | F: GGGGGTACCTGATCAGCCTGAGAGAGAGTGA R: GTTCACTCGAGACTCAGCTCCCCTTTTGGAG | 210 | |
| Intron 2 (CSNP3) | F: GGGGGTACCTCACAATTTGTAAGGTTATA R: GTTCACTCGAGTCTTAAAATTCAAGAGTAAG | 191 | |
| Downstream 5.3 kb (CSNP4) | F: GGGGGTACCTCACCAACAGCCCACTACAC R: GTTCACTCGAGGGAGCTCAGACGCAACATGA | 189 | |
| Downstream 5.6 kb (CSNP5) | F: GGGGGTACCCTGCTATCTGCTGGCGTTGT R: GTTCACTCGAGGCCCTTCCAACCGCTATCTA | 124 |
| Name | Description 1 | Position Relative to CCKAR | LG Allele 2 | HG Allele 2 | Conserved Alleles among Bird Species |
|---|---|---|---|---|---|
| CSNP1 | g.73199343G > A | Upstream 3.8 kb | G | A | G |
| CSNP2 | g.73199573A > G | Upstream 4.0 kb | A | G | A |
| CSNP3 | g.73205750T > C | Intron 2 | T | C | T |
| CSNP4 | g.73215540_73215541insTCGTCGGCC | Downstream 5.3 kb | - | TCGTCGGCC | TCGTCGGCC |
| CSNP5 | g.73215877A > G | Downstream 5.6 kb | A | G | A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Reid, A.M.A.; Wilson, P.W.; Dunn, I.C. Identification of the Core Promoter and Variants Regulating Chicken CCKAR Expression. Genes 2022, 13, 1083. https://doi.org/10.3390/genes13061083
Wang Z, Reid AMA, Wilson PW, Dunn IC. Identification of the Core Promoter and Variants Regulating Chicken CCKAR Expression. Genes. 2022; 13(6):1083. https://doi.org/10.3390/genes13061083
Chicago/Turabian StyleWang, Zhepeng, Angus M. A. Reid, Peter W. Wilson, and Ian C. Dunn. 2022. "Identification of the Core Promoter and Variants Regulating Chicken CCKAR Expression" Genes 13, no. 6: 1083. https://doi.org/10.3390/genes13061083