
Connexin Expression in Pituitary Adenomas and the Effects of Overexpression of Connexin 43 in Pituitary Tumor Cell Lines

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical-Pathological Features
2.2. RNA Extraction
2.3. Quantitative Reverse Transcriptase-Polymerase Chain Reaction
2.4. Immunohistochemistry Analysis
2.5. Cell Culture
2.6. Cx43 Transient Transfection
2.7. Cell Count Assay
2.8. Cell Immunofluorescence
2.9. Cell Cycle
2.10. TUNEL Assay
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. mRNA Expression of Connexin 26, 32, and 43 in Pituitary Neuroendocrine Adenomas
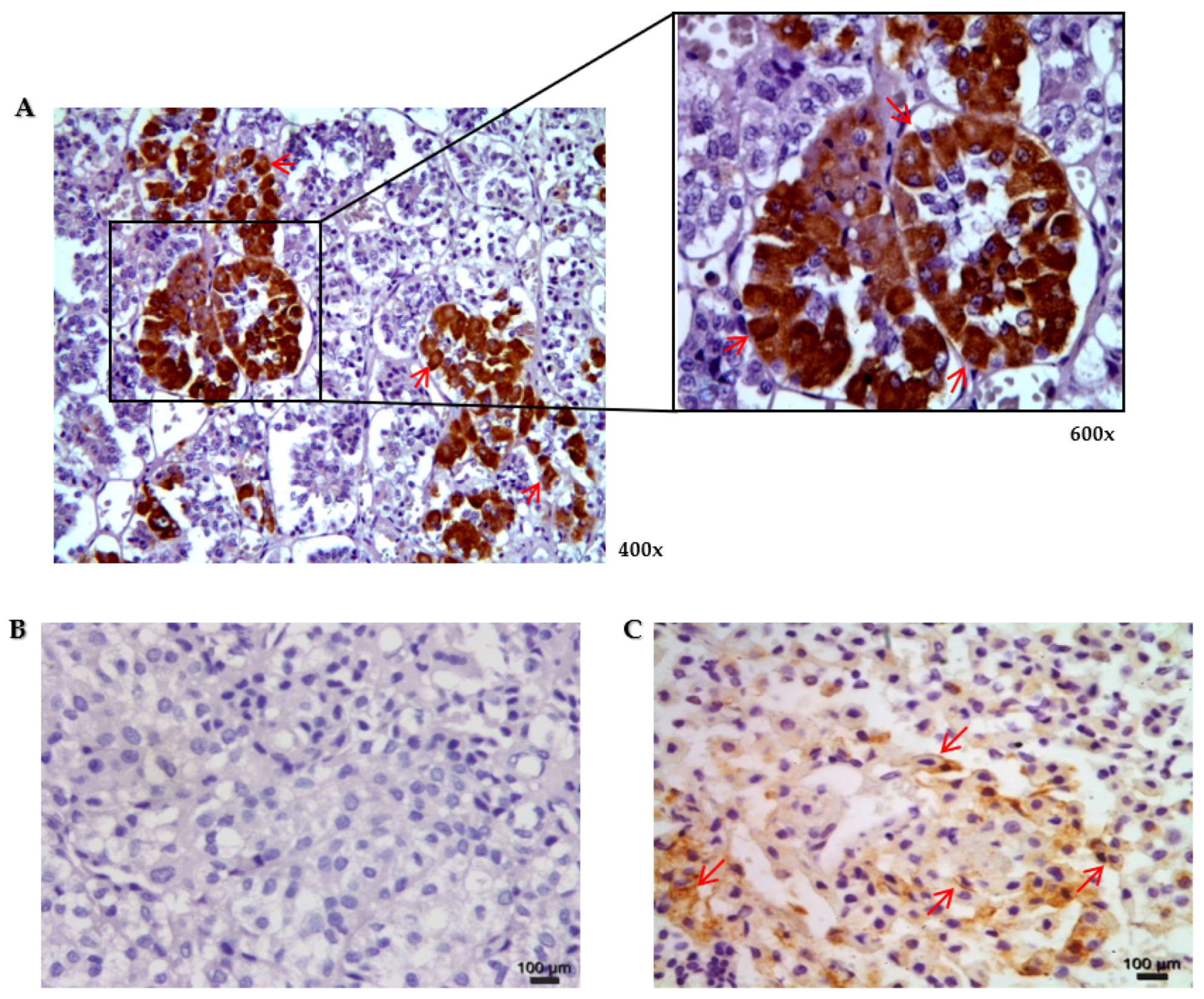
3.2. Protein Expression of Connexin 43 in Normal Human Pituitary and Pituitary Neuroendocrine Adenomas
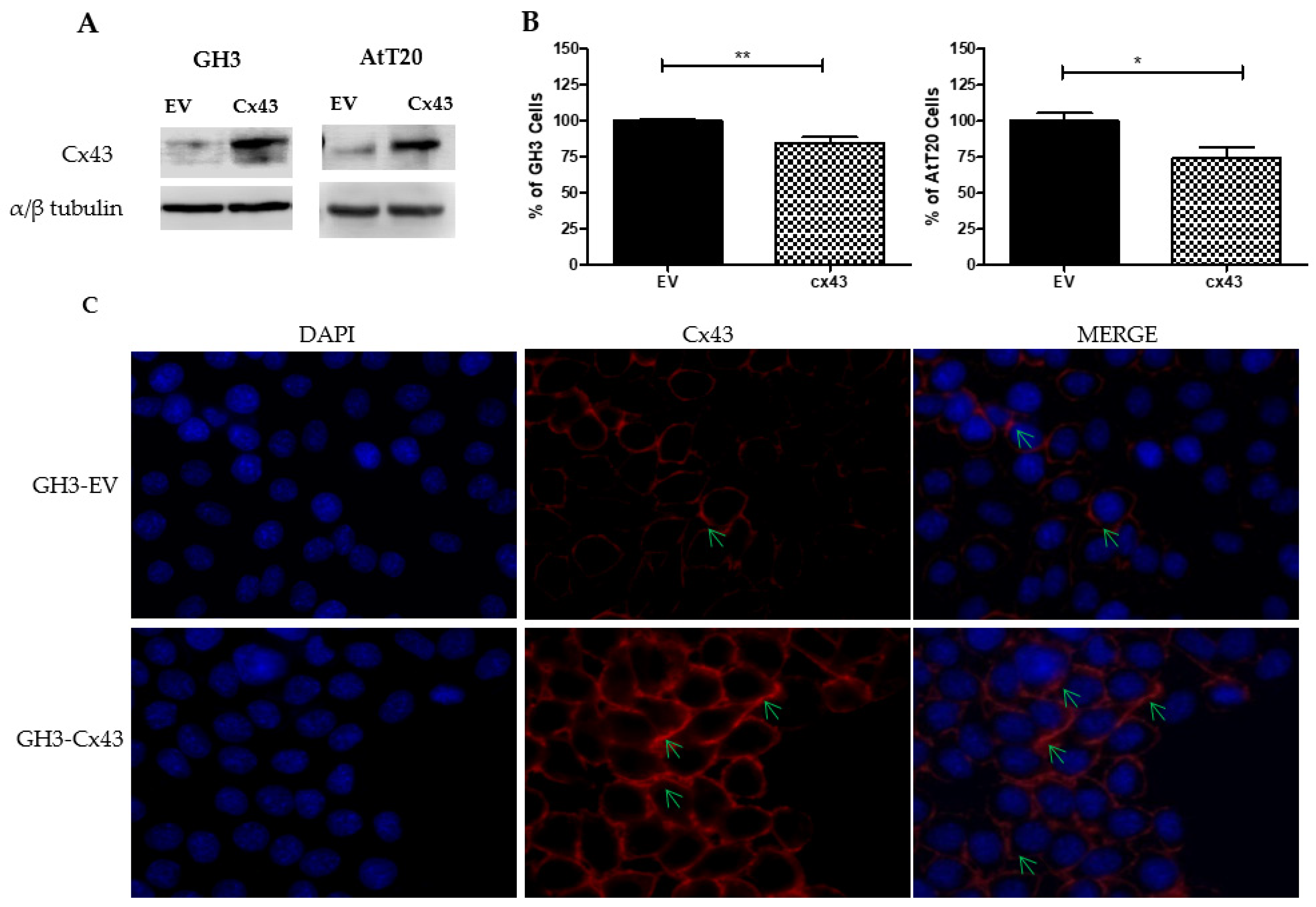
3.3. Overexpression of Cx43 Decreases Cell Number in Pituitary Tumor Cell Lines
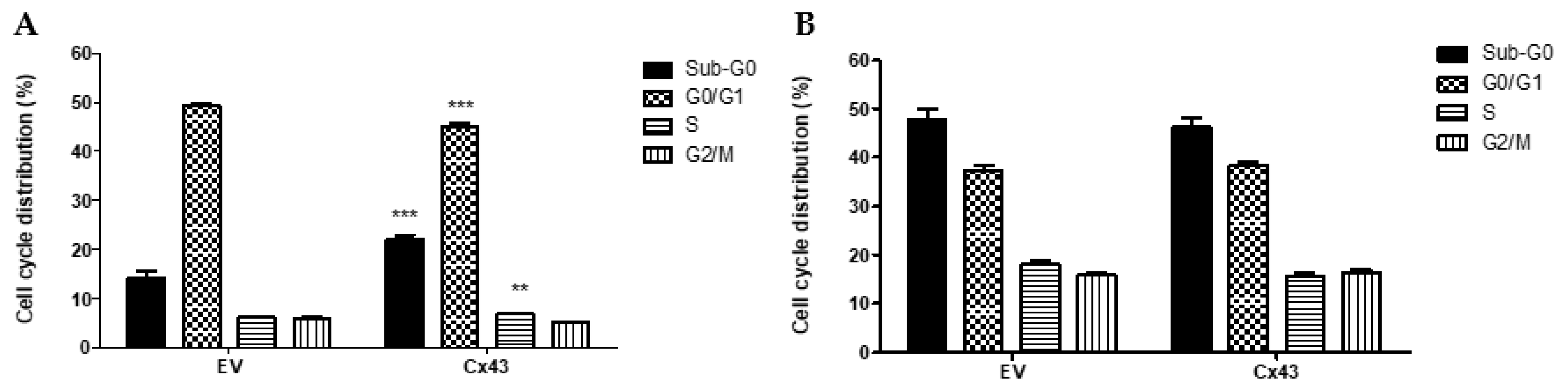
3.4. Overexpression of Cx43 Affects the Cell Cycle in Pituitary Tumor Cell Lines
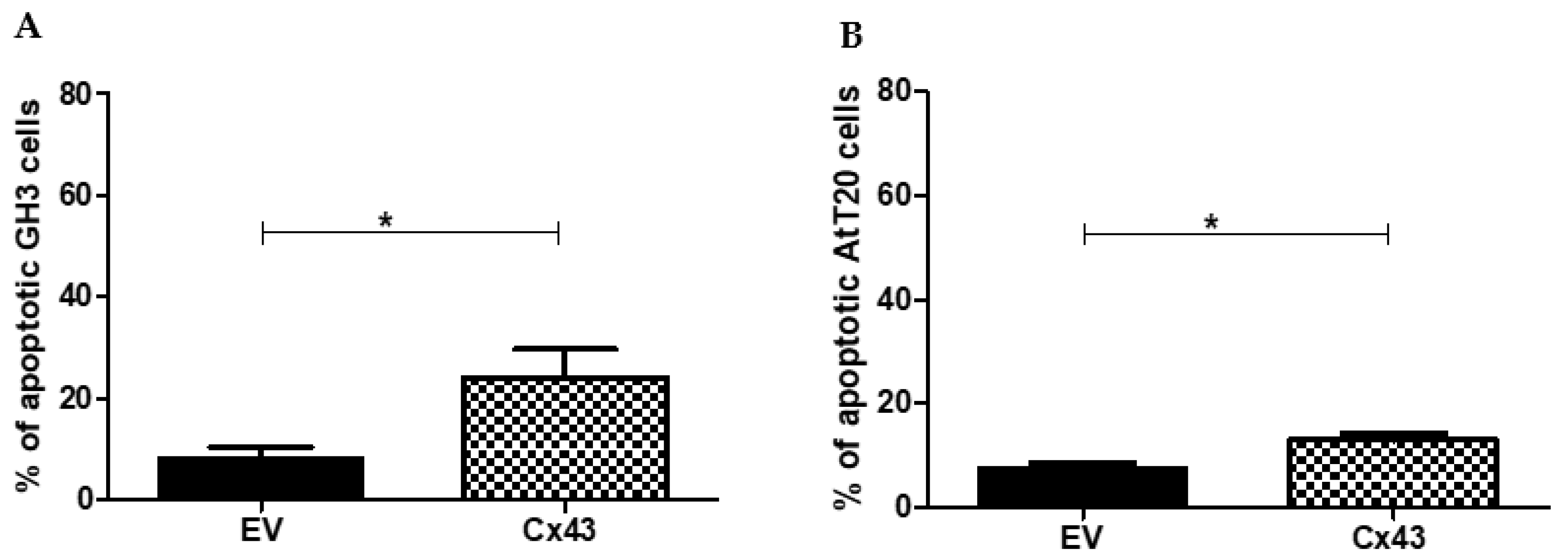
3.5. Overexpression of Cx43 Increases the Number of Apoptotic Cells in Pituitary Tumor Cell Lines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamasaki, H.; Mensnil, M.; Omori, Y.; Mironov, N.; Krutovskikh, V. Intercellular communication and carcinogenesis. Mutat. Rev. 1995, 333, 181–188. [Google Scholar] [CrossRef]
- Aasen, T.; Mesnil, M.; Naus, C.C.; Lampe, P.D.; Laird, D.W. Gap junctions and cancer: Communicating for 50 years. Nat. Rev. Cancer. 2016, 16, 775–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, N.M.; Gilula, N.B. The gap junctions communication channels. Cell 1996, 84, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Brücher, B.L.; Jamall, I.S. Cell-cell communication in the tumor microenvironment, carcinogenesis, and anticancer treatment. Cell Physiol. Biochem. 2014, 34, 213–243. [Google Scholar] [CrossRef]
- Mesnil, M.; Crespin, S.; Avanzo, J.L.; Zaidan-Dagli, M.L. Defective gap junctional intercellular communication in the carcinogenic process. Biochim. Biophys. Acta. 2005, 1719, 125–145. [Google Scholar] [CrossRef] [Green Version]
- Kar, R.; Batra, N.; Riquelme, M.A.; Jiang, J.X. Biological role of connexin intercellular channels and hemichannels. Arch. Biochem. Biophys. 2012, 524, 2–15. [Google Scholar] [CrossRef] [Green Version]
- Hervé, J.C.; Derangeon, M. Gap-junction-mediated cell-to-cell communication. Cell Tissue Res. 2013, 352, 21–31. [Google Scholar] [CrossRef]
- Carette, D.; Gilleron, J.; Chevallier, D.; Segretain, D.; Pointis, G. Connexin a check-point component of cell apoptosis in normal and physiological conditions. Biochimie 2014, 101, 1–9. [Google Scholar] [CrossRef]
- Le Tissier, P.R.; Hodson, D.J.; Lafont, C.; Fontanaud, P.; Schaeffer, M.; Mollard, P. Anterior pituitary cell networks. Front. Neuroendocrinol. 2012, 33, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Hodson, D.J.; Legros, C.; Desarme’nien, M.G.; Gue’rineau, N.C. Roles of connexins and pannexins in (neuro)endocrine physiology. Cell. Mol. Life Sci. 2015, 72, 2911–2928. [Google Scholar] [CrossRef]
- Laird, D.W.; Lampe, P.D. Therapeutic strategies targeting connexins. Nat. Rev. Drug Discov. 2018, 17, 905–921. [Google Scholar] [CrossRef] [PubMed]
- Vinken, M.; Decrock, E.; De Vuyst, E.; Ponsaerts, R.; D’hondt, C.; Bultynck, G.; Ceelen, L.; Vanhaecke, T.; Leybaert, L.; Rogiers, V. Connexins: Sensors and regulators of cell cycling. Biochim. Biophys. Acta 2011, 1815, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Bonacquisti, E.E.; Nguyen, J. Connexin 43(Cx43) in câncer: Implications for therapeutic approaches via gap junctions. Cancer Lett. 2019, 442, 439–444. [Google Scholar] [CrossRef]
- Talhouk, R.S.; Fares, M.B.; Rahme, G.J.; Hariri, H.H.; Rayess, T.; Dbouk, H.A.; Bazzoun, D.; El-Labban, D.; El-Sabban, M.E. Context dependent reversion of tumor phenotype by connexin-43 expression in MDA-MB231 cells and MCF-7: Role of β-Catenin/connexin43 association. Exp. Cell Res. 2013, 319, 3065–3080. [Google Scholar] [CrossRef]
- Mesnil, M.; Aasen, T.; Boucher, J.; Chepied, A.; Cronier, L.; Defamie, N.; Kameritsch, P.; Laird, D.W.; Lampe, P.D.; Lathia, J.D.; et al. An Update on minding the gap in cancer. Biochim. Biophys. Acta 2018, 1860, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Aasen, T.; Leithe, E.; Graham, S.V.; Kameritsch, P.; Mayán, M.D.; Mesnil, M.; Pogoda, K.; Tabernero, A. Connexins in cancer: Bridging the gap to the clinic. Oncogene 2019, 38, 4429–4451. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.P.; Fan, Y.; Hossain, M.Z.; Peng, A.; Zeng, Z.L.; Boynton, A.L. Reversion of the neoplastic phenotype of human glioblastoma cells by connexin 43 (Cx43). Cancer Res. 1998, 58, 5089–5096. [Google Scholar]
- Brockmeyer, P.; Hemmerlein, B.; Jung, K.; Falco, F.; Brodmann, T.; Gruber, R.M.; Schliephake HKramer, F.J. Connexin subtype expression during oral carcinogenesis: A pilot study in patients with oral squamous cell carcinoma. Mol. Clin. Oncol. 2016, 4, 298–302. [Google Scholar] [CrossRef] [Green Version]
- Ableser, M.J.; Penuela, S.; Lee, J.; Shao, Q.; Laird, D.W. Connexin43 reduces melanoma growth within a keratinocyte microenvironment and during tumorigenesis in vivo. J. Biol. Chem. 2014, 289, 1592–1603. [Google Scholar] [CrossRef] [Green Version]
- Tittarelli, A.; Guerrero, I.; Tempio, F.; Gleisner, M.A.; Avalos, I.; Sabanegh, S.; Ortı´z, C.; Michea, L.; Lopez, M.N.; Mendoza-Naranjo, A.; et al. Overexpression of connexin 43 reduces melanoma proliferative and metastatic capacity. Br. J. Cancer 2015, 113, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.P.; Hossain, M.Z.; Huang, R.; Gano, J.; Fan, Y.; Boynton, A. Connexin 43 (Cx43) enhances chemotherapy-induced apoptosis in human glioblastoma cells. Int. J Cancer 2001, 92, 130–138. [Google Scholar] [CrossRef]
- Alonso, F.; Domingos-Pereira, S.; Le Gal, L.; Derré, L.; Meda, P.; Jichlinski, P.; Nardelli-Haefliger, D.; Haefliger, J.A. Targeting endothelial connexin40 inhibits tumor growth by reducing angiogenesis and improving vessel perfusion. Oncotarget 2016, 7, 14015–14028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morand, I.; Fonlupt, P.; Guerrier, A.; Trouillas, J.; Calle, A.; Remy, C.; Rousset, B.; Munari-Silem, Y. Cell-to-cell communication in the anterior pituitary: Evidence for gap junction-mediated exchanges between endocrine cells and folliculostellate cells. Endocrinology 1996, 137, 3356–3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheithauer, B.W.; Gaffey, T.A.; Lloyd, R.V.; Sebo, T.J.; Kovacs, K.T.; Horvath, E.; Yapicier, O.; Young, W.F.; Meyer, F.B.; Kuroki, T.; et al. Pathobiology of pituitary adenomas and carcinomas. Neurosurgery 2006, 59, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Portovedo, S.; Gaido, N.; Nunes, B.A.; Nascimento, G.; Rocha, A.; Magalhães, M.; Nascimento, G.C.; Carvalho, D.P.; Takiya, C.; Faria, M.S.; et al. Differential Expression of HMGA1 and HMGA2 in pituitary neuroendocrine tumors. Mol. Cell. Endocrinol. 2019, 490, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Sav, A.; Rotondo, F.; Syro, L.V.; Di Ieva, A. Cusimano MD and Kovacs K. Invasive, atypical and aggressive pituitary adenomas and carcinomas. Endocrinol. Metab. Clin. 2015, 44, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Trouillas, J.; Roy, P.; Sturm, N.; Dantony, E.; Cortet-Rudelli, C.; Viennet, G.; Bonneville, J.F.; Assaker, R.; Auger, C.; Brue, T.; et al. A new prognostic clinicopathological classification of pituitary adenomas: A multicentric case-control study of 410 patients with 8 years post-operative follow-up. Acta. Neuropathol. 2013, 126, 123–135. [Google Scholar] [CrossRef]
- Trouillas, J.; Jaffrain-Rea, M.L.; Vasiljevic, A.; Raverot, G.; Roncaroli, F.; Villa, C. How to Classify Pituitary Neuroendocrine Tumors (PitNETs)s. Cancers 2020, 12, 514. [Google Scholar] [CrossRef] [Green Version]
- Di Ieva, A.; Rotondo, F.; Syro, L.V.; Cusimano, M.D.; Kovacs, K. Aggressive pituitary adenomas-diagnosis and emerging treatments. Nat. Rev. Endocrinol. 2014, 10, 423–435. [Google Scholar] [CrossRef]
- Furth, J.; Gadsen, E.L.; Upion, A.C. ACTH secreting transplantable pituitary tumors. Biol. Div. Natl. Lab. 1956, 84, 253–254. [Google Scholar] [CrossRef]
- Buonassisi, V.; Sato, T.G.; Cohen, A.I. Hormone-producing cultures of adrenal and pituitary tumor origin. Proc. Natl. Acad. Sci. USA 1962, 48, 1184–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashjian, A.H.; Yasumura, Y.; Levine, L.; Sato, G.H.; Parker, M.L. Establishment of Clonal Strains of Rat Pituitary Tumor Cells That Secrete Growth Hormone. Endocrinology 1968, 82, 342. [Google Scholar] [CrossRef] [PubMed]
- Daita, G.; Yonemasu, Y. Dural invasion and proliferative potential of pituitary adenomas. Neurol. Med. Chir. 1996, 36, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Meda, P.; Pepper, M.S.; Traub, O.; Willecke, K.; Gros, D.; Beyer, E.; Nicholson, B.; Paul, D.; Orci, L. Differential expression of gap junction connexins in endocrine and exocrine glands. Endocrinology 1993, 133, 2371–2378. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Hossain, M.Z.; Hertzberg, E.L.; Uemura, H.; Murphy, L.J.; Nagy, J.I. Connexin43 in rat pituitary: Localization at pituicyte and stellate cell gap junctions and within gonadotrophs. Histochemistry 1993, 100, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Guerineau, N.C.; Bonnefont, X.; Stoeckel, L.; Mollard, P. Synchronized spontaneous Ca2+transients in acute anterior pituitary slices. J. Biol. Chem. 1998, 273, 10389–10395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fauquier, T.; Guerineau, N.C.; McKinney, R.A.; Bauer, K.; Mollard, P. Folliculostellate cell network: A route for long-distance communication in the anterior pituitary. Proc. Natl. Acad. Sci. USA 2001, 98, 8891–8896. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, H. Aberrant expression and function of gap junctions during carcinogenesis. Environ. Health Perspect. 1991, 93, 191–197. [Google Scholar] [CrossRef]
- Xing, Y.; Xiao, Y.; Zeng, F.; Zhao, J.; Xiao, C.; Xiong PFeng, W. Altered expression of connexin-43 and impaired capacity of gap junctional intercellular communication in prostate cancer cells. J. Huazhong Univ. Sci. Technol. Med. Sci. 2007, 27, 291–294. [Google Scholar] [CrossRef]
- Vinken, M.; Decrock, E.; Leybaert, L.; Bultynck, G.; Himpens, B.; Vanhaecke, T.; Rogiers, V. Non-channel functions of connexins in cell growth and cell death. Biochim. Biophys. Acta 2012, 1818, 2002–2008. [Google Scholar] [CrossRef]
- Zhang, Y.; Kaneda, M.; Morita, I. The gap junction-independent tumor-suppressing effect of connexin 43. J. Biol. Chem. 2003, 278, 44852–44856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ionta, M.; Ferreira, R.A.; Pfister, S.C.; Machado-Santelli, G.M. Exogenous Cx43 expression decrease cell proliferation rate in rat hepatocarcinoma cells independently of functional gap junction. Cancer Cell Int. 2009, 13, 9:22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horiguchi, K.; Fujiwara, K.; Kouki, T.; Kikuchi, M.; Yashiro, T. Immunohistochemistry of connexin 43 throughout anterior pituitary gland in a transgenic rat with green fluorescent protein-expressing folliculo-stellate cells. Anat. Sci. Int. 2008, 83, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Ai, X.L.; Chi, Q.; Qiu, Y.; Li, H.Y.; Li, D.J.; Wang, J.X.; Wang, Z.Y. Gap junction protein connexin43 deregulation contributes to bladder carcinogenesis via targeting MAPK pathway. Mol. Cell. Biochem. 2017, 428, 109–118. [Google Scholar] [CrossRef]
- Cronier, L.; Crespin, S.; Strale, P.O.; Defamie, N.; Mesnil, M. Gap junctions and cancer: New functions for an old story. Antioxid. Redox Signal. 2009, 11, 323–338. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Nakayama, K.; Morita, I. A novel route for connexin 43 to inhibit cell proliferation: Negative regulation of S-phase kinase-associated protein (Skp2). Cancer Res. 2003, 63, 1623–1630. [Google Scholar]
- Wang, M.; Berthoud, V.M.; Beyer, E.C. Connexin43 increases the sensitivity of prostate cancer cells to TNFα-induced apoptosis. J. Cell Sci. 2007, 120, 320–329. [Google Scholar] [CrossRef] [Green Version]
- Iacobas, D.A.; UrbanMaldonado, M.; Iacobas, S.; Scemes, E.; Spray, D.C. Array analysis of gene expression in connexin43 null astrocytes. Physiol. Genom. 2003, 15, 177190. [Google Scholar] [CrossRef] [Green Version]
- Teleki, I.; Krenacs, T.; Szasz, M.A.; Kulka, J.; Wichmann, B.; Leo, C.; Papassotiropoulos, B.; Riemenschnitter, C.; Moch, H.; Varga, Z. The potential prognostic value of connexin 26 and 46 expression in neoadjuvant-treated breast cancer. BMC Cancer 2013, 13, 50. [Google Scholar] [CrossRef] [Green Version]
- Puzzo, L.; Caltabiano, R.; Parenti, R.; Trapasso, S.; Allegra, E. Connexin 43 (Cx43) Expression in Laryngeal Squamous Cell Carcinomas: Preliminary Data on Its Possible Prognostic Role. Head Neck Pathol. 2016, 10, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Sirnes, S.; Bruun, J.; Kolberg, M.; Kjenseth, A.; Lind, G.E.; Svindland, A.; Brech, A.; Nesbakken, A.; Lothe, R.A.; Leithe, E.; et al. Connexin43 acts as a colorectal cancer tumor suppressor and predicts disease outcome. Int. J. Cancer 2012, 131, 570–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Zhou, A.; Zhang, R.; Meng, Q. Effects of silencing connexin43 on expression of pituitary tumor-transforming gene in prolactinomas. Neurol. Res. 2015, 37, 153–158. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical-Pathological Features | PitNETs (n) | ||
|---|---|---|---|
| NF-PitNETs * | Somatotropinomas | Corticotropinomas | |
| Number of cases | 38 | 27 | 13 |
| Age (years) at surgery (n, %) | |||
| ≤45 | 15 (39) | 19 (70) | 9 (70) |
| >45 | 23 (61) | 8 (30) | 4 (30) |
| Gender (n, %) | |||
| Female | 15 (39) | 14 (50) | 12 (92) |
| Male | 23 (61) | 13 (50) | 1 (8) |
| Tumor size (n, %) | 30 | 19 | 7 |
| Microadenomas | 0 (0) | 2 (11) | 3 (43) |
| Macroadenomas | 15 (50) | 12 (63) | 4 (57) |
| Giant adenoma | 15 (50) | 5 (26) | 0 (0) |
| Invasiveness (n, %) | 28 | 19 | 6 |
| Noninvasive | 9 (32) | 4 (21) | 0 (0) |
| Invasive | 19 (68) | 15 (79) | 6 (100) |
| N. of Cases | Connexin 26 | Connexin 32 | Connexin 43 | |
|---|---|---|---|---|
| NF-PitNET * | 31 | |||
| Median (min-max) | 0.24 (0–24.51) | 0.15 (0–23.15) | 0.19 (0–4.49) | |
| Somatotropinomas | 20 | |||
| Median (min-max) | 1.27 (0–14.91) | 0.59 (0–13.47) | 0.38(0–3.06) | |
| Corticotropinomas | 9 | |||
| Median (min-max) | 0.5 (0–14.62) | 0.61 (0–11.17) | 1.03 (4.63) | |
| Total | 60 |
| Connexin 43 Expression | ||||
|---|---|---|---|---|
| Tumor Type | N. of Cases | Negative | Low | Moderate/High |
| NF-PitNETs * | 30 | 26 | 2 | 2 |
| Somatotropinomas | 24 | 21 | 3 | 0 |
| Corticotropinomas | 11 | 3 | 3 | 5 |
| Total | 65 | 50 | 8 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nunes, B.; Pópulo, H.; Lopes, J.M.; Reis, M.; Nascimento, G.; Nascimento, A.G.; Fernandes, J.; Faria, M.; de Carvalho, D.P.; Soares, P.; et al. Connexin Expression in Pituitary Adenomas and the Effects of Overexpression of Connexin 43 in Pituitary Tumor Cell Lines. Genes 2022, 13, 674. https://doi.org/10.3390/genes13040674
Nunes B, Pópulo H, Lopes JM, Reis M, Nascimento G, Nascimento AG, Fernandes J, Faria M, de Carvalho DP, Soares P, et al. Connexin Expression in Pituitary Adenomas and the Effects of Overexpression of Connexin 43 in Pituitary Tumor Cell Lines. Genes. 2022; 13(4):674. https://doi.org/10.3390/genes13040674
Chicago/Turabian StyleNunes, Bruno, Helena Pópulo, José Manuel Lopes, Marta Reis, Gilvan Nascimento, Ana Giselia Nascimento, Janaína Fernandes, Manuel Faria, Denise Pires de Carvalho, Paula Soares, and et al. 2022. "Connexin Expression in Pituitary Adenomas and the Effects of Overexpression of Connexin 43 in Pituitary Tumor Cell Lines" Genes 13, no. 4: 674. https://doi.org/10.3390/genes13040674