
Using Sex-Linked Markers via Genotyping-by-Sequencing to Identify XX/XY Sex Chromosomes in the Spiny Frog (Quasipaa boulengeri)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Sampling
2.2. Genotyping-by-Sequencing
2.3. Filtering and SNP Calling
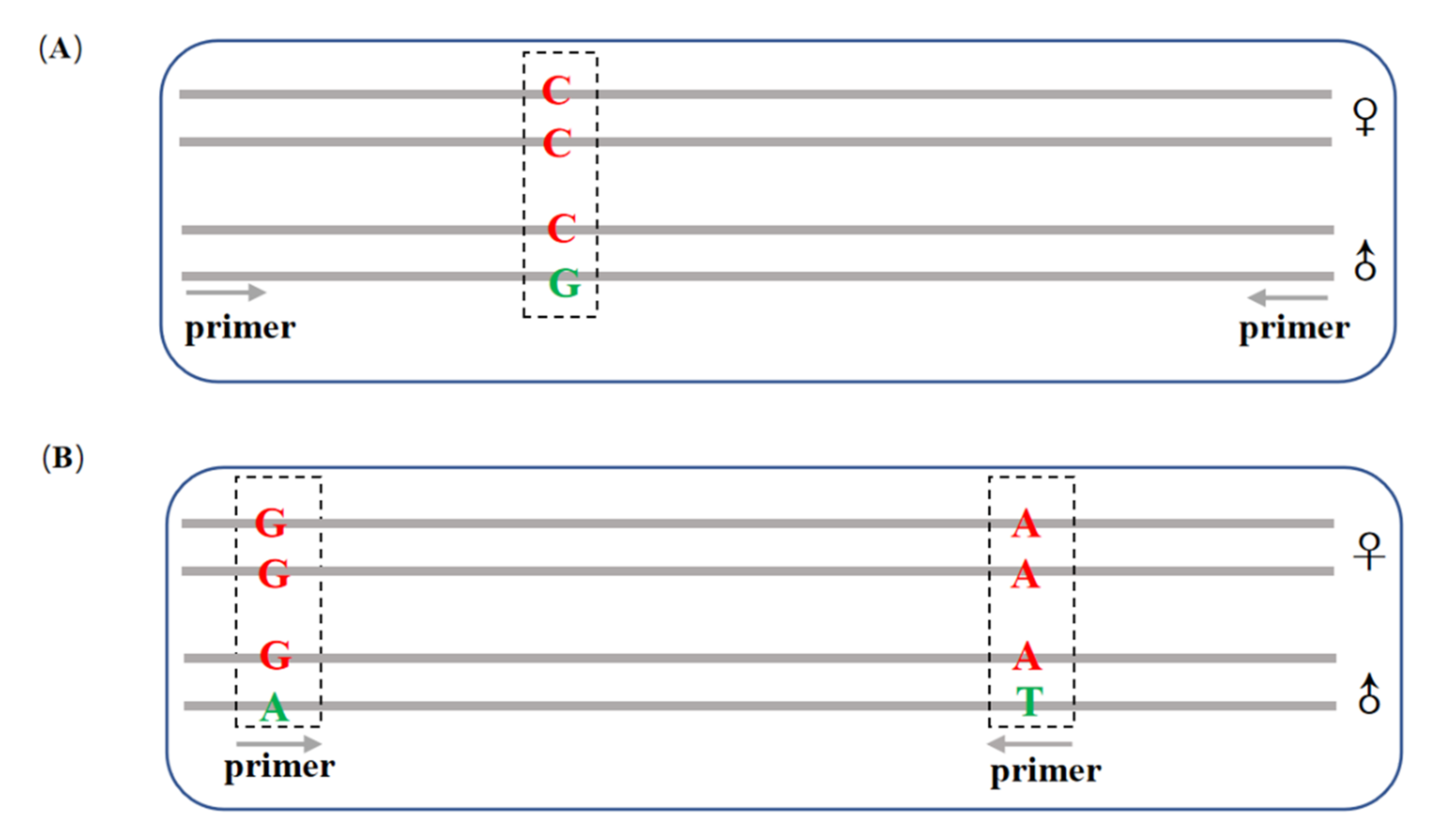
2.4. Screening Sex-Linked Markers
2.5. Confirmation of Sex-Specific Markers
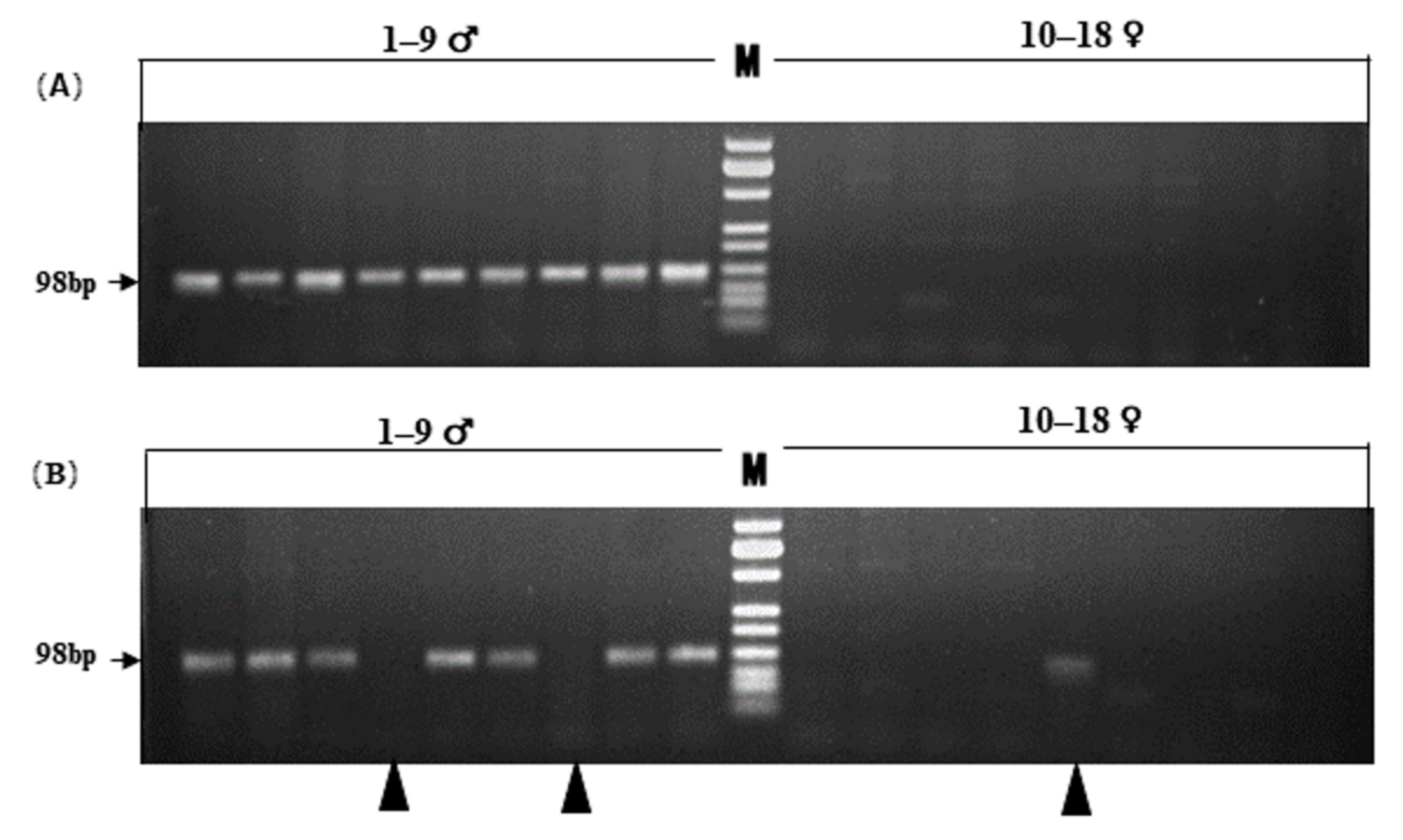
2.6. PCR Validation
2.7. Genes Associated with Sex Determination or Sex Differentiation on Chromosome 1
3. Results
3.1. GBS Data Analyses and Sex-Linked Marker Screening
3.2. Confirmation of the Putatively Sex-Linked Marker
3.3. Validation of the Sex-Linked SNP Markers
3.4. Potential Sex-Determining Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hillis, D.M.; Green, D.M. Evolutionary changes of heterogametic sex in the phylogenetic history of amphibians. J. Evol. Biol. 1990, 3, 49–64. [Google Scholar] [CrossRef]
- Miura, I. Sex Determination and Sex Chromosomes in Amphibia. Sex. Dev. 2017, 11, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M. Sex chromosome evolution in non-mammalian vertebrates. Curr. Opin. Genet. Dev. 2004, 14, 634–641. [Google Scholar] [CrossRef]
- Brelsford, A.; Lavanchy, G.; Sermier, R.; Rausch, A.; Perrin, N. Identifying homomorphic sex chromosomes from wild-caught adults with limited genomic resources. Mol. Ecol. Resour. 2017, 17, 752–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffries, D.L.; Lavanchy, G.; Sermier, R.; Sredl, M.J.; Miura, I.; Borzee, A.; Barrow, L.N.; Canestrelli, D.; Crochet, P.A.; Dufresnes, C.; et al. A rapid rate of sex-chromosome turnover and non-random transitions in true frogs. Nat. Commun. 2018, 9, 4088. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.R.; Skelly, D.K.; Ezaz, T. Sex-linked markers in the North American green frog (Rana clamitans) developed using DArTseq provide early insight into sex chromosome evolution. BMC Genom. 2016, 17, 844. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Xia, Y.; Yue, B.; Zeng, X. Assigning the Sex-Specific Markers via Genotyping-by-Sequencing onto the Y Chromosome for a Torrent Frog Amolops mantzorum. Genes 2020, 11, 727. [Google Scholar] [CrossRef]
- Hu, Q.; Chang, C.; Wang, Q.; Tian, H.; Qiao, Z.; Wang, L.; Meng, Y.; Xu, C.; Xiao, H. Genome-wide RAD sequencing to identify a sex-specific marker in Chinese giant salamander Andrias davidianus. BMC Genom. 2019, 20, 415. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Castoe, T.A.; Nielsen, S.V.; Banks, J.L.; Card, D.C.; Schield, D.R.; Schuett, G.W.; Booth, W. The Discovery of XY Sex Chromosomes in a Boa and Python. Curr. Biol. 2017, 27, 2148–2153.e2144. [Google Scholar] [CrossRef] [Green Version]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction Site-Associated DNA Sequencing (RAD-seq) Reveals an Extraordinary Number of Transitions among Gecko Sex-Determining Systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; McKenna, E.; Meyer, W.; Nielsen, S.V.; Pinto, B.J.; Scantlebury, D.P.; Higham, T.E. XX/XY Sex Chromosomes in the South American Dwarf Gecko (Gonatodes humeralis). J. Hered. 2018, 109, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.; Zarkower, D. Identification of sex-specific molecular markers using restriction site-associated DNA sequencing. Mol. Ecol. Resour. 2014, 14, 902–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, P.L.; Burridge, C.P.; Ezaz, T.; Wapstra, E. Conservation of Sex-Linked Markers among Conspecific Populations of a Viviparous Skink, Niveoscincus ocellatus, Exhibiting Genetic and Temperature-Dependent Sex Determination. Genome Biol. Evol. 2018, 10, 1079–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, L.; Xia, Y.; Zheng, Y.; Zeng, X. A de novo case of floating chromosomal polymorphisms by translocation in Quasipaa boulengeri (Anura, Dicroglossidae). PLoS ONE 2012, 7, e46163. [Google Scholar] [CrossRef] [Green Version]
- Yuan, S.; Xia, Y.; Zeng, X. A Sex-linked Microsatellite Marker Reveals Male Heterogamety in Quasipaa boulengeri(Anura: Dicroglossidae). Asian Herpetol. Res. 2017, 8, 184–189. [Google Scholar] [CrossRef]
- Yuan, X.; Xia, Y.; Zeng, X. Suppressed Recombination of Sex Chromosomes Is Not Caused by Chromosomal Reciprocal Translocation in Spiny Frog (Quasipaa boulengeri). Front. Genet. 2018, 9, 288. [Google Scholar] [CrossRef]
- Brelsford, A.; Stock, M.; Betto-Colliard, C.; Dubey, S.; Dufresnes, C.; Jourdan-Pineau, H.; Rodrigues, N.; Savary, R.; Sermier, R.; Perrin, N. Homologous sex chromosomes in three deeply divergent anuran species. Evolution 2013, 67, 2434–2440. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Xia, Y.; Zeng, X. Sex chromosomal dimorphisms narrated by X-chromosome translocation in a spiny frog (Quasipaa boulengeri). Front. Zool. 2018, 15, 47. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.; Hohenlohe, P.A.; Bassham, S.; Amores, A.; Cresko, W.A. Stacks: An analysis tool set for population genomics. Mol. Ecol. 2013, 22, 3124–3140. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Yuan, S.; Liu, Y.; Xia, Y.; Zeng, X. Microsatellites mapping for non-model species with chromosomal rearrangement: A case study in the frog Quasipaa boulengeri (Anura: Dicroglossidae). Genome 2017, 60, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Yuan, X.; Luo, W.; Yuan, S.; Zeng, X. The Origin and Evolution of Chromosomal Reciprocal Translocation in Quasipaa boulengeri (Anura, Dicroglossidae). Front. Genet. 2019, 10, 1364. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.B.; Xiong, Z.J.; Xiang, X.Y.; Liu, S.P.; Zhou, W.W.; Tu, X.L.; Zhong, L.; Wang, L.; Wu, D.D.; Zhang, B.L.; et al. Whole-genome sequence of the Tibetan frog Nanorana parker and the comparative evolution of tetrapod genomes. Proc. Natl. Acad. Sci. USA 2015, 112, 201501764. [Google Scholar] [CrossRef] [Green Version]
- Purcell, J.; Brelsford, A.; Wurm, Y.; Perrin, N.; Chapuisat, M. Convergent Genetic Architecture Underlies Social Organization in Ants. Curr. Biol. 2014, 24, 2728–2732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778. [Google Scholar] [CrossRef] [Green Version]
- Raymond, C.S.; Parker, E.D.; Kettlewell, J.R.; Brown, L.G.; Page, D.C.; Kusz, K.; Jaruzelska, J.; Reinberg, Y.; Flejter, W.L.; Bardwell, V.J.; et al. A Region of Human Chromosome 9p Required for Testis Development Contains Two Genes Related to Known Sexual Regulators. Hum. Mol. Genet. 1999, 8, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Roeszler, K.N.; Ohnesorg, T.; Cummins, D.M.; Farlie, P.G.; Doran, T.J.; Sinclair, A.H. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken. Nature 2009, 461, 267–271. [Google Scholar] [CrossRef]
- Yoshimoto, S.; Okada, E.; Umemoto, H.; Tamura, K.; Uno, Y.; Nishida-Umehara, C.; Matsuda, Y.; Takamatsu, N.; Shiba, T.; Ito, M. A W-linked DM-domain gene, DM-W, participates in primary ovary development in Xenopus laevis. Proc. Natl. Acad. Sci. USA 2008, 105, 2469. [Google Scholar] [CrossRef] [Green Version]
- Brelsford, A.; Dufresnes, C.; Perrin, N. High-density sex-specific linkage maps of a European tree frog (Hyla arborea) identify the sex chromosome without information on offspring sex. Heredity 2016, 116, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, N.; Studer, T.; Dufressnes, C.; Ma, W.-J.; Veltsos, P.; Perrin, N. Dmrt1 polymorphism and sex-chromosome differentiation in Rana temporaria. Mol. Ecol. 2017, 26, 4897–4905. [Google Scholar] [CrossRef] [PubMed]
- Miura, I. An evolutionary witness: The frog Rana rugosa underwent change of heterogametic sex from XY male to ZW female. Sex. Dev. 2007, 1, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Sumida, M.; Nishioka, M. Sex-linked genes and linkage maps in amphibians. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 126, 257–270. [Google Scholar] [CrossRef]
- Alho, J.S.; Matsuba, C.; MerilÄ, J. Sex reversal and primary sex ratios in the common frog (Rana temporaria). Mol. Ecol. 2010, 19, 1763–1773. [Google Scholar] [CrossRef]
- Hayes, T.B.; Khoury, V.; Narayan, A.; Nazir, M.; Park, A.; Brown, T.; Adame, L.; Chan, E.; Buchholz, D.; Stueve, T.; et al. Atrazine induces complete feminization and chemical castration in male African clawed frogs (Xenopus laevis). Proc. Natl. Acad. Sci. USA 2010, 107, 4612–4617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamschick, S.; Rozenblut-Kościsty, B.; Ogielska, M.; Lehmann, A.; Lymberakis, P.; Hoffmann, F.; Lutz, I.; Kloas, W.; Stöck, M. Sex reversal assessments reveal different vulnerability to endocrine disruption between deeply diverged anuran lineages. Sci. Rep. 2016, 6, 23825. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| GBS-Tags | Primer ID | Primer Sequences (5′-3′) | Site Type | Product Size (bp) |
|---|---|---|---|---|
| 712545 | QS1 | Forward: ACAAAGCTAGTGAAACATGATGGTC | SD | 183 |
| Reverse: CAAACACACAGGCATGGCAA | ||||
| 70112 | QS18 | Forward: TAGCTTACTTGCACATCA | SD | 283 |
| Reverse: AGCCCAAATCCCTCTTAG | ||||
| 35863 | QS30 | Forward: TATGTCAGATGGCTTCAGGACG | SD | 255 |
| Reverse: GCTCCGTGTGCTCCTTACA | ||||
| 35863 | QS29 | Forward: TAAAACGTTCACATACTATA | PA | 98 |
| Reverse: GTCCAGCCAGTCACTGGTTCA | ||||
| 465977 | QS43 | Forward: AGTAATAACAATCTACAAGCAT | SD | 258 |
| Reverse: ATGTAATGTCCCCAAGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Luo, W.; Xia, Y.; Zeng, X. Using Sex-Linked Markers via Genotyping-by-Sequencing to Identify XX/XY Sex Chromosomes in the Spiny Frog (Quasipaa boulengeri). Genes 2022, 13, 575. https://doi.org/10.3390/genes13040575
Yang X, Luo W, Xia Y, Zeng X. Using Sex-Linked Markers via Genotyping-by-Sequencing to Identify XX/XY Sex Chromosomes in the Spiny Frog (Quasipaa boulengeri). Genes. 2022; 13(4):575. https://doi.org/10.3390/genes13040575
Chicago/Turabian StyleYang, Xusheng, Wei Luo, Yun Xia, and Xiaomao Zeng. 2022. "Using Sex-Linked Markers via Genotyping-by-Sequencing to Identify XX/XY Sex Chromosomes in the Spiny Frog (Quasipaa boulengeri)" Genes 13, no. 4: 575. https://doi.org/10.3390/genes13040575