
Circum-Saharan Prehistory through the Lens of mtDNA Diversity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
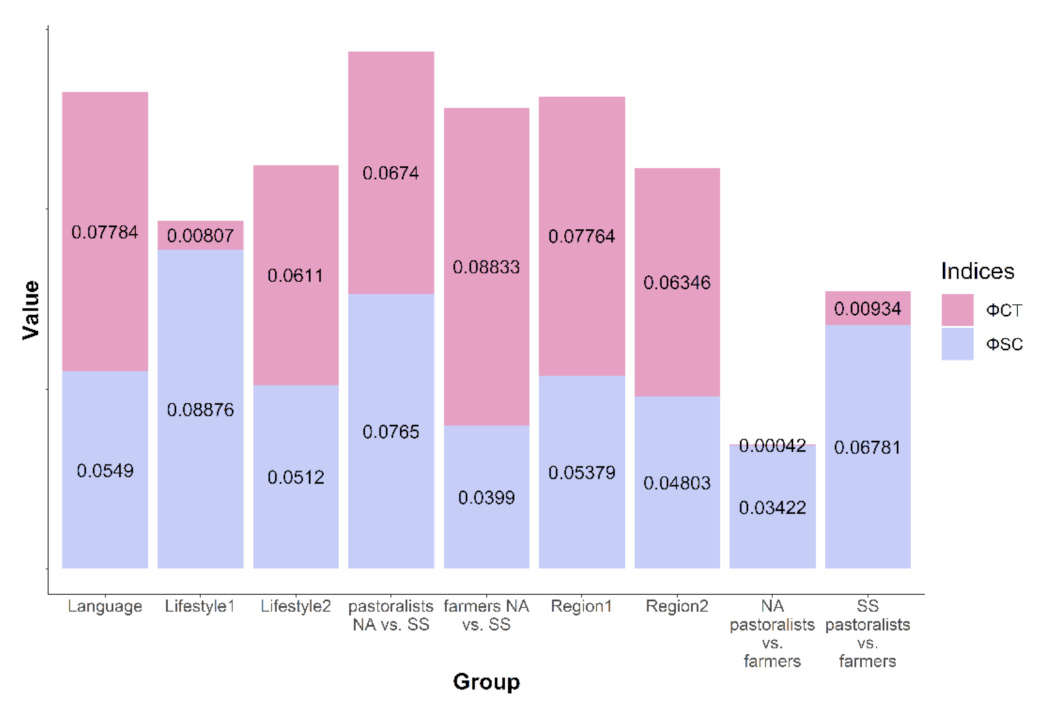
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bae, C.J.; Douka, K.; Petraglia, M.D. On the Origin of Modern Humans: Asian Perspectives. Science 2017, 358, eaai9067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvati, K.; Röding, C.; Bosman, A.M.; Karakostis, F.A.; Grün, R.; Stringer, C.; Karkanas, P.; Thompson, N.C.; Koutoulidis, V.; Moulopoulos, L.A.; et al. Apidima Cave Fossils Provide Earliest Evidence of Homo Sapiens in Eurasia. Nature 2019, 571, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Malaspinas, A.-S.; Westaway, M.C.; Muller, C.; Sousa, V.C.; Lao, O.; Alves, I.; Bergström, A.; Athanasiadis, G.; Cheng, J.Y.; Crawford, J.E.; et al. A Genomic History of Aboriginal Australia. Nature 2016, 538, 207–214. [Google Scholar] [CrossRef]
- Fernandes, V.; Alshamali, F.; Alves, M.; Costa, M.D.; Pereira, J.B.; Silva, N.M.; Cherni, L.; Harich, N.; Cerny, V.; Soares, P.; et al. The Arabian Cradle: Mitochondrial Relicts of the First Steps along the Southern Route out of Africa. Am. J. Hum. Genet. 2012, 90, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auton, A.; Abecasis, G.R.; Altshuler, D.M.; Durbin, R.M.; Abecasis, G.R.; Bentley, D.R.; Chakravarti, A.; Clark, A.G.; Donnelly, P.; Eichler, E.E.; et al. A Global Reference for Human Genetic Variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Behar, D.M.; Metspalu, E.; Kivisild, T.; Rosset, S.; Tzur, S.; Hadid, Y.; Yudkovsky, G.; Rosengarten, D.; Pereira, L.; Amorim, A.; et al. Counting the Founders: The Matrilineal Genetic Ancestry of the Jewish Diaspora. PLoS ONE 2008, 3, e2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kivisild, T. Maternal Ancestry and Population History from Whole Mitochondrial Genomes. Investig. Genet. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Černý, V.; Pereira, L.; Musilová, E.; Kujanová, M.; Vašíková, A.; Blasi, P.; Garofalo, L.; Soares, P.; Diallo, I.; Brdička, R.; et al. Genetic Structure of Pastoral and Farmer Populations in the African Sahel. Mol. Biol. Evol. 2011, 28, 2491–2500. [Google Scholar] [CrossRef] [Green Version]
- Čížková, M.; Munclinger, P.; Diallo, M.Y.; Kulichová, I.; Mokhtar, M.G.; Dème, A.; Pereira, L.; Černý, V. Genetic Structure of the Western and Eastern African Sahel/Savannah Belt and the Role of Nomadic Pastoralists as Inferred from the Variation of D-Loop Mitochondrial DNA Sequences. Hum. Biol. 2017, 89, 281–302. [Google Scholar] [CrossRef] [PubMed]
- Nováčková, J.; Čížková, M.; Mokhtar, M.G.; Duda, P.; Stenzl, V.; Tříska, P.; Hofmanová, Z.; Černý, V. Subsistence Strategy Was the Main Factor Driving Population Differentiation in the Bidirectional Corridor of the African Sahel. Am. J. Phys. Anthr. 2020, 171, 496–508. [Google Scholar] [CrossRef]
- Černý, V.; Salas, A.; Hájek, M.; Zaloudková, M.; Brdicka, R. A Bidirectional Corridor in the Sahel-Sudan Belt and the Distinctive Features of the Chad Basin Populations: A History Revealed by the Mitochondrial DNA Genome. Ann. Hum. Genet. 2007, 71, 433–452. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Černý, V.; Cerezo, M.; Silva, N.M.; Hájek, M.; Vasíková, A.; Kujanová, M.; Brdicka, R.; Salas, A. Linking the Sub-Saharan and West Eurasian Gene Pools: Maternal and Paternal Heritage of the Tuareg Nomads from the African Sahel. Eur. J. Hum. Genet. 2010, 18, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podgorná, E.; Soares, P.; Pereira, L.; Černý, V. The Genetic Impact of the Lake Chad Basin Population in North Africa as Documented by Mitochondrial Diversity and Internal Variation of the L3e5 Haplogroup. Ann. Hum. Genet. 2013, 77, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Blench, R. The Westward Wanderings of Cushitic Pastoralists: Explorations in the Prehistory of Cental Africa. In L’Homme et l’animal dans le bassin du lac Tchad; Editions IRD: Paris, France, 1999; pp. 39–80. [Google Scholar]
- Černý, V.; Fernandes, V.; Costa, M.D.; Hájek, M.; Mulligan, C.J.; Pereira, L. Migration of Chadic Speaking Pastoralists within Africa Based on Population Structure of Chad Basin and Phylogeography of Mitochondrial L3f Haplogroup. BMC Evol. Biol. 2009, 9, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerezo, M.; Černý, V.; Carracedo, Á.; Salas, A. New Insights into the Lake Chad Basin Population Structure Revealed by High-Throughput Genotyping of Mitochondrial DNA Coding SNPs. PLoS ONE 2011, 6, e18682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achilli, A.; Rengo, C.; Battaglia, V.; Pala, M.; Olivieri, A.; Fornarino, S.; Magri, C.; Scozzari, R.; Babudri, N.; Santachiara-Benerecetti, A.S.; et al. Saami and Berbers--an Unexpected Mitochondrial DNA Link. Am. J. Hum. Genet. 2005, 76, 883–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulichová, I.; Fernandes, V.; Deme, A.; Nováčková, J.; Stenzl, V.; Novelletto, A.; Pereira, L.; Černý, V. Internal Diversification of Non-Sub-Saharan Haplogroups in Sahelian Populations and the Spread of Pastoralism beyond the Sahara. Am. J. Phys. Anthropol. 2017, 164, 424–434. [Google Scholar] [CrossRef]
- Gifford-Gonzalez, D.; Hanotte, O. Domesticating Animals in Africa: Implications of Genetic and Archaeological Findings. J. World Prehistory 2011, 24, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Ozainne, S. Un Néolithique Ouest-Africain: Cadre Chrono-Culturel, Économique et Environnemental de l’Holocène Récent En Pays Dogon (Mali); Africa Magna Verlag: Frankfurt am Main, Germany, 2013; Volume 8, ISBN 3-937248-33-1. [Google Scholar]
- Ozainne, S.; Lespez, L.; Garnier, A.; Ballouche, A.; Neumann, K.; Pays, O.; Huysecom, E. A Question of Timing: Spatio-Temporal Structure and Mechanisms of Early Agriculture Expansion in West Africa. J. Archaeol. Sci. 2014, 50, 359–368. [Google Scholar] [CrossRef]
- Sikora, M.; Carpenter, M.L.; Moreno-Estrada, A.; Henn, B.M.; Underhill, P.A.; Sánchez-Quinto, F.; Zara, I.; Pitzalis, M.; Sidore, C.; Busonero, F.; et al. Population Genomic Analysis of Ancient and Modern Genomes Yields New Insights into the Genetic Ancestry of the Tyrolean Iceman and the Genetic Structure of Europe. PLoS Genet. 2014, 10, e1004353. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.W.K.; Marcus, J.H.; Sidore, C.; Biddanda, A.; Al-Asadi, H.; Zoledziewska, M.; Pitzalis, M.; Busonero, F.; Maschio, A.; Pistis, G.; et al. Genomic History of the Sardinian Population. Nat. Genet. 2018, 50, 1426–1434. [Google Scholar] [CrossRef] [PubMed]
- Vicente, M.; Priehodová, E.; Diallo, I.; Podgorná, E.; Poloni, E.S.; Černý, V.; Schlebusch, C.M. Population History and Genetic Adaptation of the Fulani Nomads: Inferences from Genome-Wide Data and the Lactase Persistence Trait. BMC Genom. 2019, 20, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priehodová, E.; Austerlitz, F.; Čížková, M.; Nováčková, J.; Ricaut, F.-X.; Hofmanová, Z.; Schlebusch, C.M.; Černý, V. Sahelian Pastoralism from the Perspective of Variants Associated with Lactase Persistence. Am. J. Phys. Anthr. 2020, 173, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Černý, V.; Fortes-Lima, C.; Tříska, P. Demographic History and Admixture Dynamics in African Sahelian Populations. Hum. Mol. Genet. 2021, 30, R29–R36. [Google Scholar] [CrossRef] [PubMed]
- Triska, P.; Soares, P.; Patin, E.; Fernandes, V.; Cerny, V.; Pereira, L. Extensive Admixture and Selective Pressure Across the Sahel Belt. Genome Biol. Evol. 2015, 7, 3484–3495. [Google Scholar] [CrossRef] [PubMed]
- Priehodová, E.; Austerlitz, F.; Čížková, M.; Mokhtar, M.G.; Poloni, E.S.; Černý, V. The Historical Spread of Arabian Pastoralists to the Eastern African Sahel Evidenced by the Lactase Persistence −13,915*G Allele and Mitochondrial DNA. Am. J. Hum. Biol. 2017, 29, e22950. [Google Scholar] [CrossRef] [PubMed]
- Levy, T.E.; Holl, A.F.C. Migrations, Ethnogenesis, and Settlement Dynamics: Israelites in Iron Age Canaan and Shuwa-Arabs in the Chad Basin. J. Anthropol. Archaeol. 2002, 21, 83–118. [Google Scholar] [CrossRef]
- Pereira, L.; Mutesa, L.; Tindana, P.; Ramsay, M. African Genetic Diversity and Adaptation Inform a Precision Medicine Agenda. Nat. Rev. Genet. 2021, 22, 284–306. [Google Scholar] [CrossRef]
- Mulindwa, J.; Noyes, H.; Ilboudo, H.; Pagani, L.; Nyangiri, O.; Kimuda, M.P.; Ahouty, B.; Asina, O.F.; Ofon, E.; Kamoto, K.; et al. High Levels of Genetic Diversity within Nilo-Saharan Populations: Implications for Human Adaptation. Am. J. Hum. Genet. 2020, 107, 473–486. [Google Scholar] [CrossRef]
- Choudhury, A.; Aron, S.; Botigué, L.R.; Sengupta, D.; Botha, G.; Bensellak, T.; Wells, G.; Kumuthini, J.; Shriner, D.; Fakim, Y.J.; et al. High-Depth African Genomes Inform Human Migration and Health. Nature 2020, 586, 741–748. [Google Scholar] [CrossRef]
- Vai, S.; Sarno, S.; Lari, M.; Luiselli, D.; Manzi, G.; Gallinaro, M.; Mataich, S.; Hübner, A.; Modi, A.; Pilli, E.; et al. Ancestral Mitochondrial N Lineage from the Neolithic ‘Green’ Sahara. Sci. Rep. 2019, 9, 3530. [Google Scholar] [CrossRef] [PubMed]
- Van Oven, M.; Kayser, M. Updated Comprehensive Phylogenetic Tree of Global Human Mitochondrial DNA Variation. Hum. Mutat. 2009, 30, E386–E394. [Google Scholar] [CrossRef] [PubMed]
- Gonder, M.K.; Mortensen, H.M.; Reed, F.A.; de Sousa, A.; Tishkoff, S.A. Whole-MtDNA Genome Sequence Analysis of Ancient African Lineages. Mol. Biol. Evol. 2007, 24, 757–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendall, K.E.; Sykes, B.C. Length Heteroplasmy in the First Hypervariable Segment of the Human MtDNA Control Region. Am. J. Hum. Genet. 1995, 57, 248. [Google Scholar]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical Tests of Neutrality of Mutations against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Ramos-Onsins, S.E.; Rozas, J. Statistical Properties of New Neutrality Tests against Population Growth. Mol. Biol. Evol. 2002, 19, 2092–2100. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population Growth Makes Waves in the Distribution of Pairwise Genetic Differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef]
- Reynolds, J.; Weir, B.S.; Cockerham, C.C. Estimation of the Coancestry Coefficient: Basis for a Short-Term Genetic Distance. Genetics 1983, 105, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M. A Correction to the Exact Test Based on the Ewens Sampling Distribution. Genet. Res. 1996, 68, 259–260. [Google Scholar] [CrossRef] [Green Version]
- Poloni, E.S.; Naciri, Y.; Bucho, R.; Niba, R.; Kervaire, B.; Excoffier, L.; Langaney, A.; Sanchez-Mazas, A. Genetic Evidence for Complexity in Ethnic Differentiation and History in East Africa. Ann. Hum. Genet. 2009, 73, 582–600. [Google Scholar] [CrossRef] [Green Version]
- Jari Oksanen, F.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Peter, R.; Minchin, R.B.; O’Hara, G.; Simpson, L.; et al. Vegan: Community Ecology Package. R Package Version; R Package: Vienna, Austria, 2020; pp. 2–5. [Google Scholar]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Beerli, P. How to Use MIGRATE or Why Are Markov Chain Monte Carlo Programs Difficult to Use? In Population Genetics for Animal Conservation; Conservation Biology; Rizzoli, A., Vernesi, C., Bertorelle, G., Hauffe, H.C., Bruford, M.W., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 42–79. ISBN 978-0-521-86630-9. [Google Scholar]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. Circlize Implements and Enhances Circular Visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henn, B.M.; Botigué, L.R.; Gravel, S.; Wang, W.; Brisbin, A.; Byrnes, J.K.; Fadhlaoui-Zid, K.; Zalloua, P.A.; Moreno-Estrada, A.; Bertranpetit, J.; et al. Genomic Ancestry of North Africans Supports Back-to-Africa Migrations. PLoS Genet. 2012, 8, e1002397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, R.M.; Kubacka, I.; Chinnery, P.F.; Lightowlers, R.N.; Turnbull, D.M.; Howell, N. Reanalysis and Revision of the Cambridge Reference Sequence for Human Mitochondrial DNA. Nat. Genet. 1999, 23, 147. [Google Scholar] [CrossRef] [PubMed]
- Forster, P.; Harding, R.; Torroni, A.; Bandelt, H.J. Origin and Evolution of Native American MtDNA Variation: A Reappraisal. Am. J. Hum. Genet. 1996, 59, 935–945. [Google Scholar]
- Soares, P.; Ermini, L.; Thomson, N.; Mormina, M.; Rito, T.; Röhl, A.; Salas, A.; Oppenheimer, S.; Macaulay, V.; Richards, M.B. Correcting for Purifying Selection: An Improved Human Mitochondrial Molecular Clock. Am. J. Hum. Genet. 2009, 84, 740–759. [Google Scholar] [CrossRef] [Green Version]
- Saillard, J.; Forster, P.; Lynnerup, N.; Bandelt, H.-J.; Nørby, S. MtDNA Variation among Greenland Eskimos: The Edge of the Beringian Expansion. Am. J. Hum. Genet. 2000, 67, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. PAML: A Program Package for Phylogenetic Analysis by Maximum Likelihood. Comput. Appl. Biosci. 1997, 13, 555–556. [Google Scholar] [CrossRef]
- Černý, V.; Hájek, M.; Bromová, M.; Čmejla, R.; Diallo, I.; Brdička, R. MtDNA of Fulani nomads and their genetic relationships to neighboring sedentary populations. Hum. Biol. 2006, 78, 9–27. [Google Scholar] [CrossRef]
- van de Loosdrecht, M.; Bouzouggar, A.; Humphrey, L.; Posth, C.; Barton, N.; Aximu-Petri, A.; Nickel, B.; Nagel, S.; Talbi, E.H.; El Hajraoui, M.A.; et al. Pleistocene North African Genomes Link Near Eastern and Sub-Saharan African Human Populations. Science 2018, 360, 548–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.C.; Alshamali, F.; Montinaro, F.; Cavadas, B.; Torroni, A.; Pereira, L.; Raveane, A.; Fernandes, V. Projecting Ancient Ancestry in Modern-Day Arabians and Iranians: A Key Role of the Past Exposed Arabo-Persian Gulf on Human Migrations. Genome Biol. Evol. 2021, 13, evab194. [Google Scholar] [CrossRef]
- Arredi, B.; Poloni, E.S.; Paracchini, S.; Zerjal, T.; Fathallah, D.M.; Makrelouf, M.; Pascali, V.L.; Novelletto, A.; Tyler-Smith, C. A Predominantly Neolithic Origin for Y-Chromosomal DNA Variation in North Africa. Am. J. Hum. Genet. 2004, 75, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Kujanová, M.; Pereira, L.; Fernandes, V.; Pereira, J.B.; Cerný, V. Near Eastern Neolithic Genetic Input in a Small Oasis of the Egyptian Western Desert. Am. J. Phys. Anthr. 2009, 140, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Černý, V.; Čížková, M.; Poloni, E.S.; Al-Meeri, A.; Mulligan, C.J. Comprehensive View of the Population History of Arabia as Inferred by MtDNA Variation. Am. J. Phys. Anthr. 2016, 159, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Currat, M.; Excoffier, L.; Penny, D. Modern Humans Did Not Admix with Neanderthals during Their Range Expansion into Europe. PLoS Biol. 2004, 2, e421. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, J.; Lopes, A.M.; Comas, D.; Amorim, A.; Arenas, M. Evaluating the Neolithic Expansion at Both Shores of the Mediterranean Sea. Mol. Biol. Evol. 2017, 34, 3232–3242. [Google Scholar] [CrossRef] [PubMed]
- Fregel, R.; Méndez, F.L.; Bokbot, Y.; Martín-Socas, D.; Camalich-Massieu, M.D.; Santana, J.; Morales, J.; Ávila-Arcos, M.C.; Underhill, P.A.; Shapiro, B.; et al. Ancient Genomes from North Africa Evidence Prehistoric Migrations to the Maghreb from Both the Levant and Europe. Proc. Natl. Acad. Sci. USA 2018, 115, 6774–6779. [Google Scholar] [CrossRef] [Green Version]
- Hernández, C.L.; Pita, G.; Cavadas, B.; López, S.; Sánchez-Martínez, L.J.; Dugoujon, J.-M.; Novelletto, A.; Cuesta, P.; Pereira, L.; Calderón, R. Human Genomic Diversity Where the Mediterranean Joins the Atlantic. Mol. Biol. Evol. 2020, 37, 1041–1055. [Google Scholar] [CrossRef]
- Bekada, A. Caractéristaion Anthropogénétique d’un Échantillon de la Population Algérienne: Analyse des Marqueurs Parentaux; Université d’Oran 1: Es Senia, Algeria, 2015. [Google Scholar]
- Adams, W.Y. Nubia: Corridor to Africa; Princeton University Press: Princeton, NJ, USA, 1977; ISBN 978-0-691-09370-3. [Google Scholar]
- Fox, C.L. MtDNA Analysis in Ancient Nubians Supports the Existence of Gene Flow between Sub-Sahara and North Africa in the Nile Valley. Ann. Hum. Biol. 1997, 24, 217–227. [Google Scholar] [CrossRef]
- Krings, M.; Salem, A.E.; Bauer, K.; Geisert, H.; Malek, A.K.; Chaix, L.; Simon, C.; Welsby, D.; Di Rienzo, A.; Utermann, G.; et al. MtDNA Analysis of Nile River Valley Populations: A Genetic Corridor or a Barrier to Migration? Am. J. Hum. Genet. 1999, 64, 1166–1176. [Google Scholar] [CrossRef] [Green Version]
- Segal, R. Islam’s Black Slaves: The History of Africa’s Other Black Diaspora; Atlantic Books: London, UK, 2002; ISBN 978-1-903809-81-5. [Google Scholar]
- Turner, M.D.; Schlecht, E. Livestock Mobility in Sub-Saharan Africa: A Critical Review. Pastoralism 2019, 9, 13. [Google Scholar] [CrossRef]
- Jesse, F.; Keding, B.; Lenssen-Erz, T.; Pöllath, N. I Hope Your Cattle Are Well’: Archaeological Evidence for Early Cattle-Centred Behaviour in the Eastern Sahara of Sudan and Chad. In Pastoralism in Africa: Past, Present and Future; Berghahn Books: New York, NY, USA, 2013; pp. 66–103. [Google Scholar]
- Linseele, V. From First Stock Keepers to Specialised Pastoralists in the West African Savannah. In Pastoralism in Africa: Past, Present and Future; Bollig, M., Schnegg, M., Wotzka, H.-P., Eds.; Berghan Books: New York, NY, USA, 2013; pp. 145–170. [Google Scholar]
- Ozainne, S.; Mayor, A.; Huysecom, E. Chronology of Human Occupation during the Holocene in West Africa. The Dogon country radiocarbon record. In Winds of change; Rudolf Habelt Verlag: Bonn, Germany, 2017; pp. 211–227. [Google Scholar]
- Wright, D.K. Humans as Agents in the Termination of the African Humid Period. Front. Earth Sci. 2017, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Manning, K.; Pelling, R.; Higham, T.; Schwenniger, J.-L.; Fuller, D.Q. 4500-Year Old Domesticated Pearl Millet (Pennisetum Glaucum) from the Tilemsi Valley, Mali: New Insights into an Alternative Cereal Domestication Pathway. J. Archaeol. Sci. 2011, 38, 312–322. [Google Scholar] [CrossRef]
- Burgarella, C.; Cubry, P.; Kane, N.A.; Varshney, R.K.; Mariac, C.; Liu, X.; Shi, C.; Thudi, M.; Couderc, M.; Xu, X.; et al. A Western Sahara Centre of Domestication Inferred from Pearl Millet Genomes. Nat. Ecol. Evol. 2018, 2, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Cubry, P.; Tranchant-Dubreuil, C.; Thuillet, A.-C.; Monat, C.; Ndjiondjop, M.-N.; Labadie, K.; Cruaud, C.; Engelen, S.; Scarcelli, N.; Rhoné, B.; et al. The Rise and Fall of African Rice Cultivation Revealed by Analysis of 246 New Genomes. Curr. Biol. 2018, 28, 2274–2282.e6. [Google Scholar] [CrossRef] [Green Version]
- Scarcelli, N.; Cubry, P.; Akakpo, R.; Thuillet, A.-C.; Obidiegwu, J.; Baco, M.N.; Otoo, E.; Sonké, B.; Dansi, A.; Djedatin, G.; et al. Yam Genomics Supports West Africa as a Major Cradle of Crop Domestication. Sci. Adv. 2019, 5, eaaw1947. [Google Scholar] [CrossRef] [Green Version]
- Winchell, F.; Stevens, C.J.; Murphy, C.; Champion, L.; Fuller, D.Q. Evidence for Sorghum Domestication in Fourth Millennium BC Eastern Sudan: Spikelet Morphology from Ceramic Impressions of the Butana Group. Curr. Anthropol. 2017, 58, 673–683. [Google Scholar] [CrossRef] [Green Version]
- Cervigni, R.; Morris, M. Confronting Drought in Africa’s Drylands: Opportunities for Enhancing Resilience; World Bank Publications: Herndon, VA, USA, 2016; ISBN 978-1-4648-0818-0. [Google Scholar]
- Kleisner, K.; Pokorný, Š.; Čížková, M.; Froment, A.; Černý, V. Nomadic Pastoralists and Sedentary Farmers of the Sahel/Savannah Belt of Africa in the Light of Geometric Morphometrics Based on Facial Portraits. Am. J. Phys. Anthropol. 2019, 169, 632–645. [Google Scholar] [CrossRef]
- Naino Jika, A.K.; Dussert, Y.; Raimond, C.; Garine, E.; Luxereau, A.; Takvorian, N.; Djermakoye, R.S.; Adam, T.; Robert, T. Unexpected Pattern of Pearl Millet Genetic Diversity among Ethno-Linguistic Groups in the Lake Chad Basin. Heredity 2017, 118, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musilová, E.; Fernandes, V.; Silva, N.M.; Soares, P.; Alshamali, F.; Harich, N.; Cherni, L.; Gaaied, A.B.; Al-Meeri, A.; Pereira, L.; et al. Population history of the Red Sea—Genetic exchanges between the Arabian Peninsula and East Africa signaled in the mitochondrial DNA HV1 haplogroup. Am. J. Phys. Anthropol. 2011, 145, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Cruciani, F.; Trombetta, B.; Sellitto, D.; Massaia, A.; Destro-Bisol, G.; Watson, E.; Beraud Colomb, E.; Dugoujon, J.-M.; Moral, P.; Scozzari, R. Human Y Chromosome Haplogroup R-V88: A Paternal Genetic Record of Early Mid Holocene Trans-Saharan Connections and the Spread of Chadic Languages. Eur. J. Hum. Genet. 2010, 18, 800–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shriner, D.; Rotimi, C.N. Genetic History of Chad. Am. J. Phys. Anthropol. 2018, 167, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, J.K.; Patterson, N.; Loh, P.-R.; Lipson, M.; Berger, B.; Stoneking, M.; Pakendorf, B.; Reich, D. Ancient West Eurasian Ancestry in Southern and Eastern Africa. Proc. Natl. Acad. Sci. USA 2014, 111, 2632–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kruskal–Wallis Test (p-Values) | ||||||
|---|---|---|---|---|---|---|
| Haplotype Diversity | Nucleotide Diversity | Mean Number of Pairwise Differences | Fu’s Fs (p) | Hri (p) | SSD (p) | |
| NA vs. SS | 0.380 | 0.000 | 0.000 | 0.563 | 0.068 | 0.208 |
| FM vs. PS | 0.004 | 0.856 | 0.630 | 0.133 | 0.271 | 0.249 |
| AA vs. NC vs. NS | 0.293 | 0.011 | 0.001 | 0.682 | 0.033 | 0.147 |
| eNA vs. eSS vs. wNA vs. wSS | 0.453 | 0.000 | 0.000 | 0.940 | 0.266 | 0.231 |
| NAFM vs. NAPS vs. SSFM vs. SSPS | 0.000 | 0.000 | 0.000 | 0.507 | 0.482 | 0.874 |
| Maximum Parsimony | Maximum Likelihood | |||||
|---|---|---|---|---|---|---|
| Total Clock | Synonymous Clock | All SNPs | ||||
| Haplogroup | Age | 95% CI | Age | 95% CI | Age | 95% CI |
| N1a1a | 24,005 | 16,482–31,778 | 20,727 | 10,873–30,580 | 21,754 | 21,304–22,182 |
| N1a1a3 | 13,900 | 10,071–17,808 | 14,266 | 7442–21,090 | 17,948 | 17,676–18,211 |
| N1a3a+195C! | 3188 | 1266–5135 | 1819 | −838–4477 | 3610 | 3577–3643 |
| N1b2+9325C | 6816 | 553–13,327 | 14,191 | −3 562–31,944 | 6908 | 6762–7048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diallo, M.Y.; Čížková, M.; Kulichová, I.; Podgorná, E.; Priehodová, E.; Nováčková, J.; Fernandes, V.; Pereira, L.; Černý, V. Circum-Saharan Prehistory through the Lens of mtDNA Diversity. Genes 2022, 13, 533. https://doi.org/10.3390/genes13030533
Diallo MY, Čížková M, Kulichová I, Podgorná E, Priehodová E, Nováčková J, Fernandes V, Pereira L, Černý V. Circum-Saharan Prehistory through the Lens of mtDNA Diversity. Genes. 2022; 13(3):533. https://doi.org/10.3390/genes13030533
Chicago/Turabian StyleDiallo, Mame Yoro, Martina Čížková, Iva Kulichová, Eliška Podgorná, Edita Priehodová, Jana Nováčková, Veronica Fernandes, Luísa Pereira, and Viktor Černý. 2022. "Circum-Saharan Prehistory through the Lens of mtDNA Diversity" Genes 13, no. 3: 533. https://doi.org/10.3390/genes13030533