
Crlz-1 Homozygous Null Knockout Mouse Embryos Are Lethally Stopped in Their Early Development



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
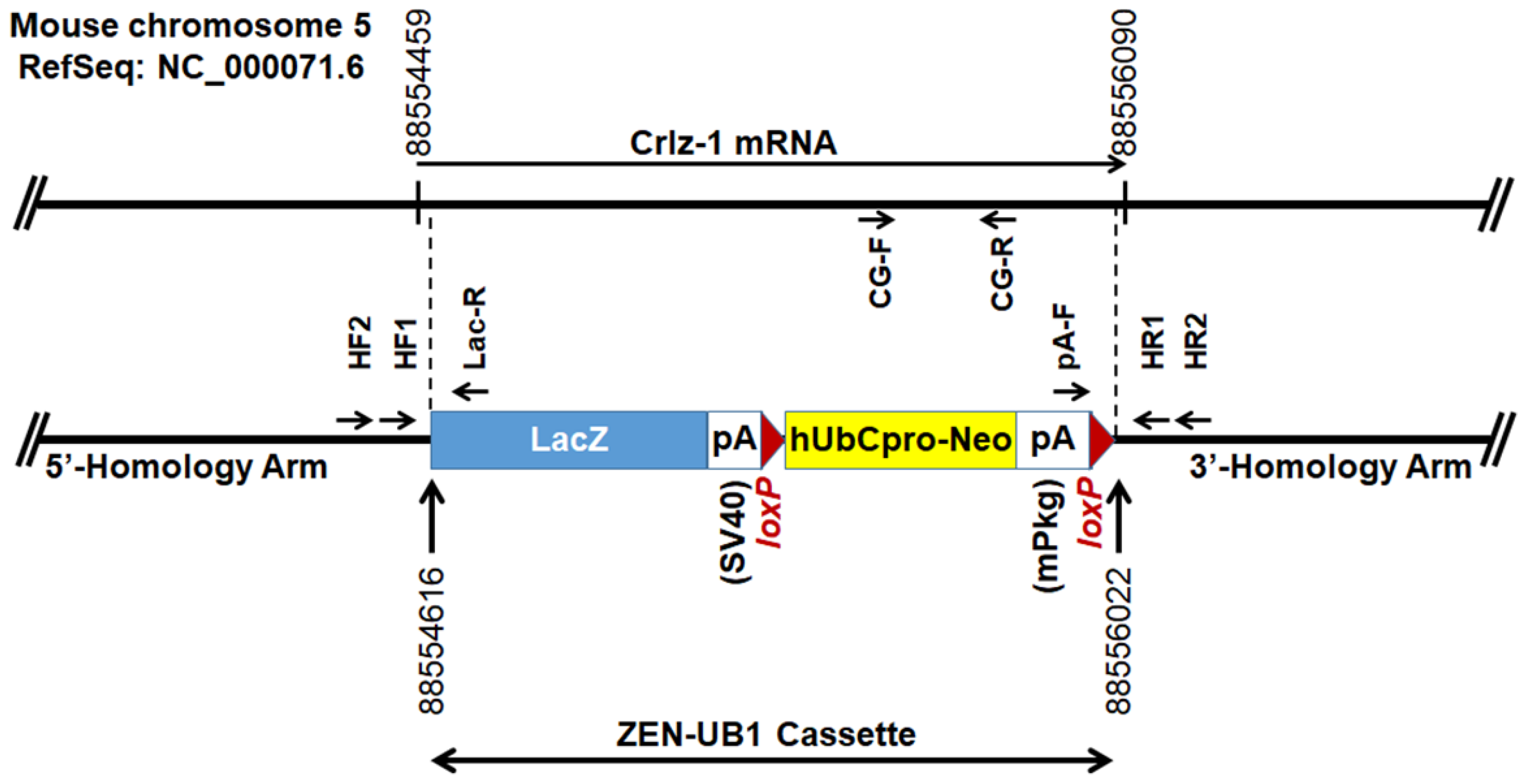
2.1. Construction of Crlz-1 (Utp3) Null KO Mice
2.2. Embryo Isolation
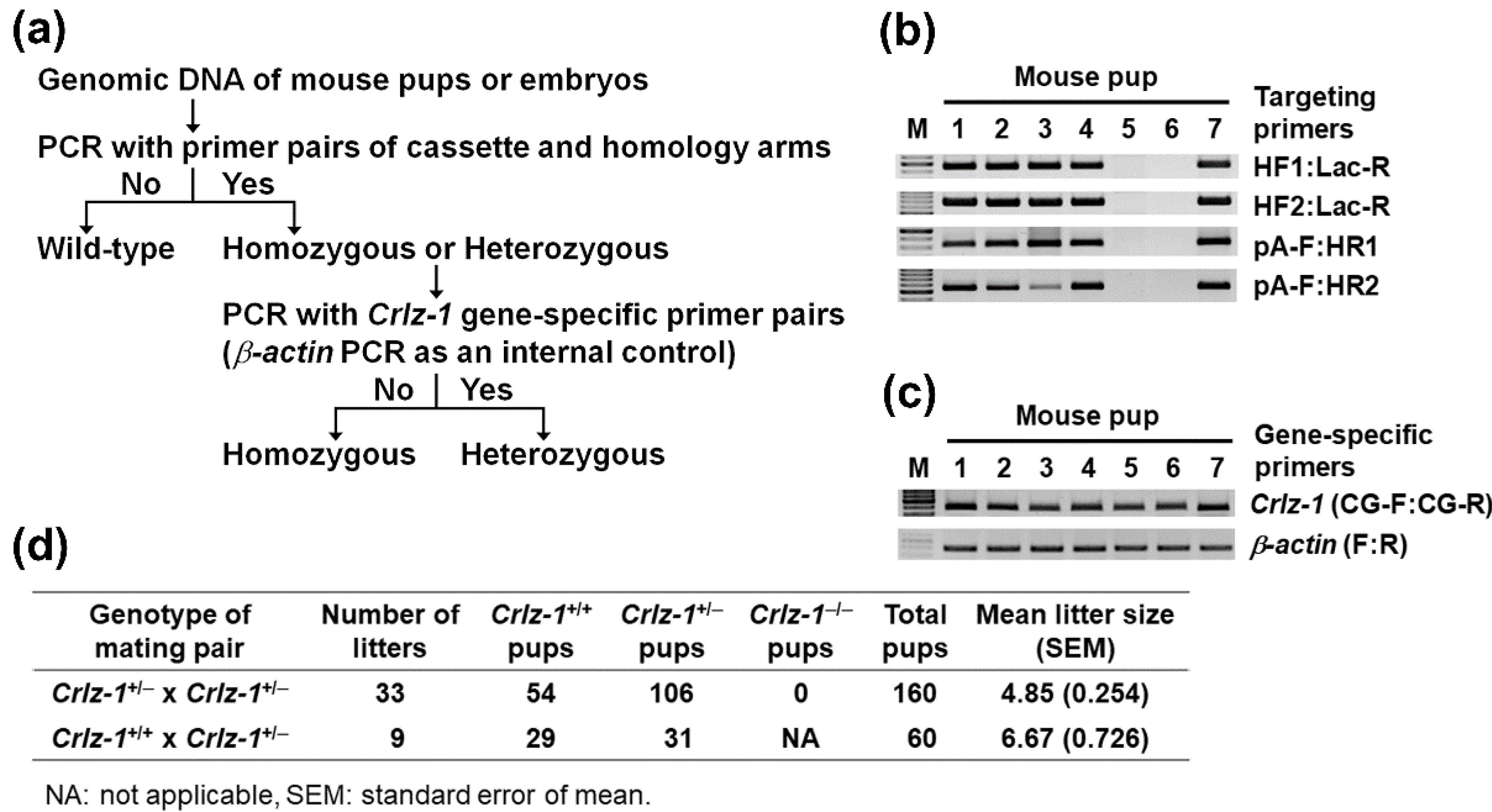
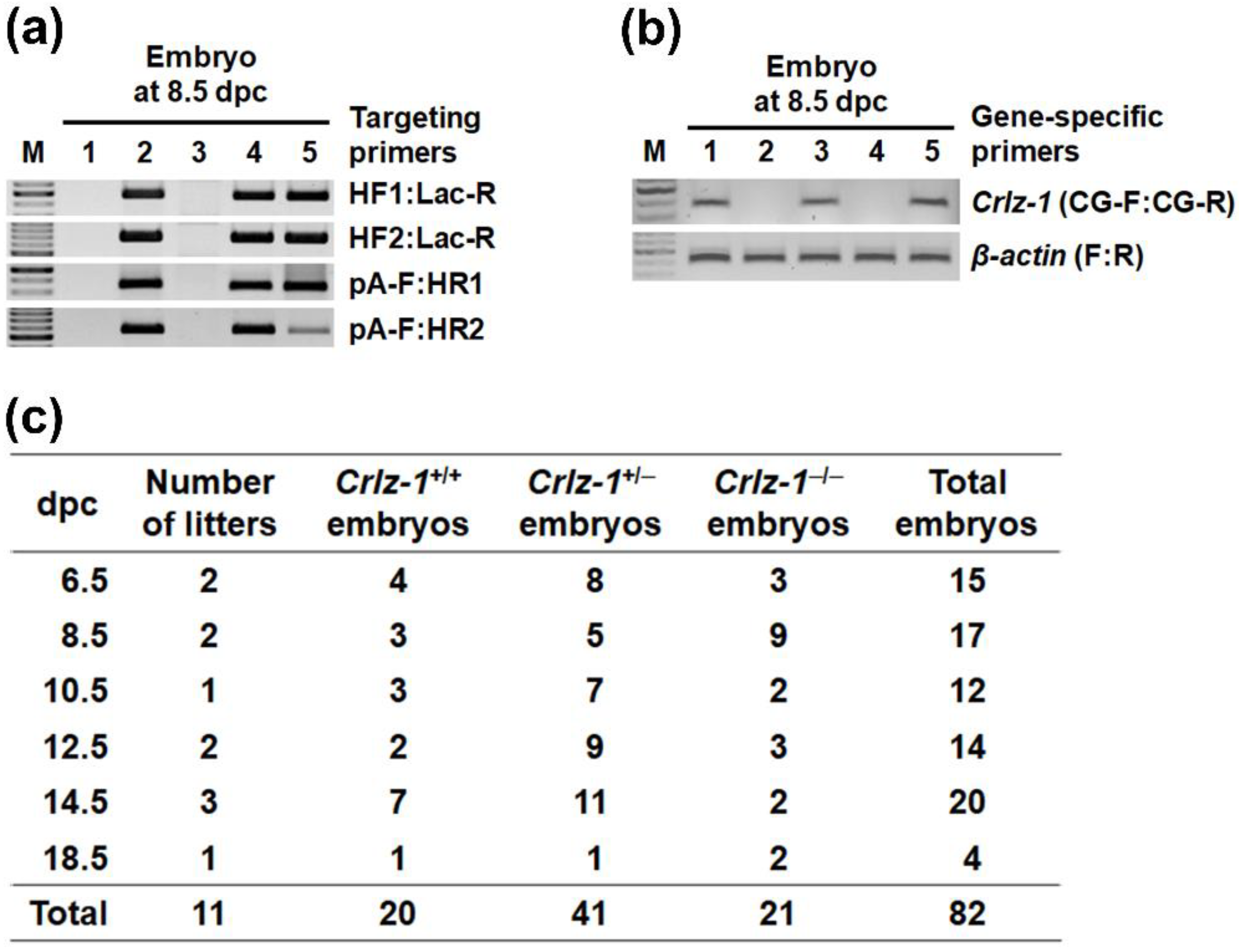
2.3. Genotyping by PCR (Polymerase Chain Reaction)
2.4. In Situ Hybridization (ISH) and Reverse Transcription (RT)-PCR
2.5. Image Analysis and Statistics
3. Results
3.1. Crlz-1 Homozygous Null KO Mouse Embryos Are Lethally Impaired in Their Development, and So Their Live Pups Are Not Born
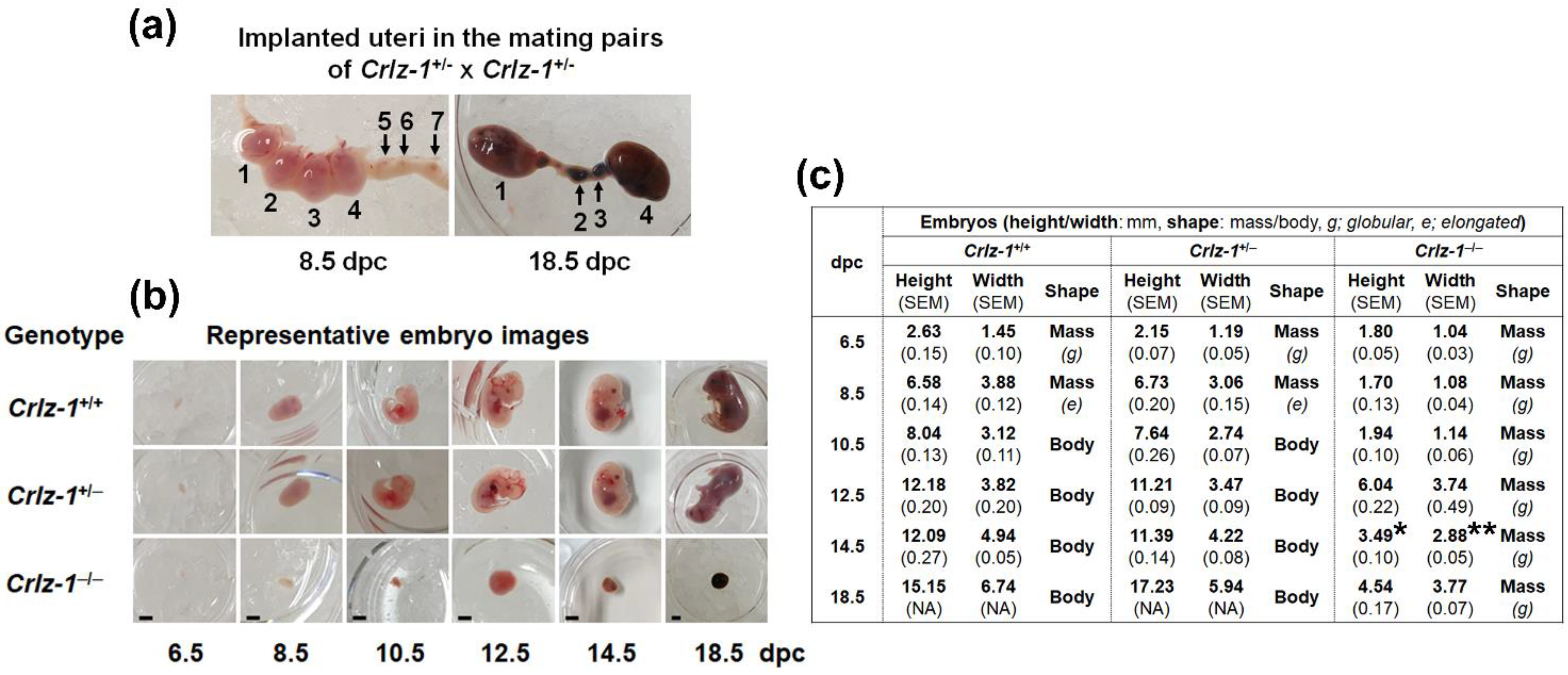
3.2. The Development of Crlz-1−/− Homozygous Null KO Mouse Embryos Was Stopped from Around 8.5 dpc, Remained in a State of Small Globular Mass, and Did Not Ever Lead to a Body Shape
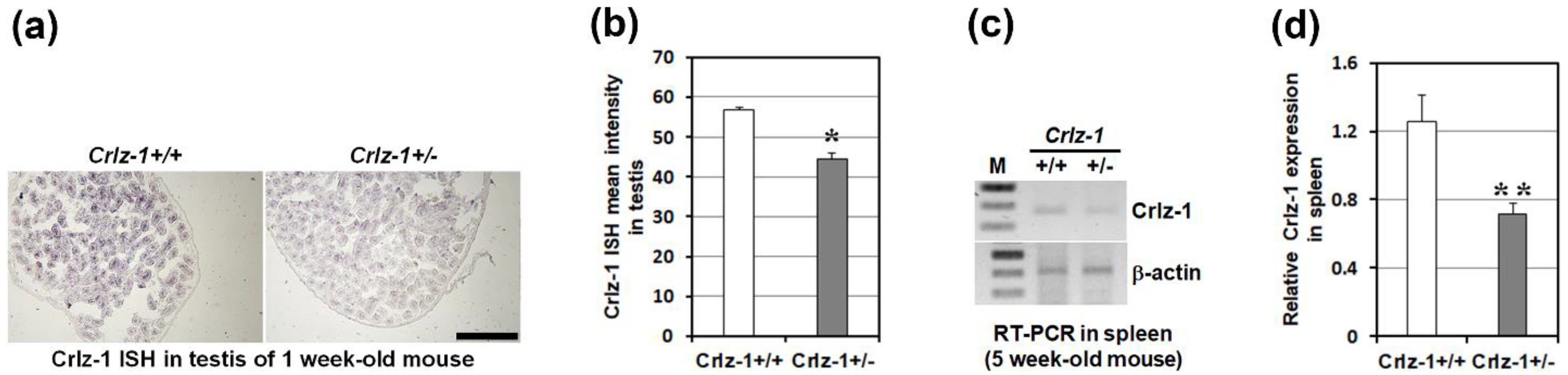
3.3. Crlz-1 Heterozygous KO Mice Looked Generally Normal despite of Its Haploinsufficient Expression
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernstein, K.A.; Baserga, S.J. The small subunit processome is required for cell cycle progression at G1. Mol. Biol. Cell 2004, 15, 5038–5046. [Google Scholar] [CrossRef] [PubMed]
- Kamakaka, R.T.; Rine, J. Sir- and silencer-independent disruption of silencing in Saccharomyces by Sas10p. Genetics 1998, 149, 903–914. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, T.; Li, Q.L.; Jin, Y.; Choi, L.W.; Kim, E.G.; Ito, K.; Ito, Y.; Nomura, S.; Bae, S.C. Cloning and expression pattern of a novel PEBP2β-binding protein (charged amino acid rich leucine zipper-1 [Crl-1]) in the mouse. Mech. Dev. 2001, 104, 151–154. [Google Scholar] [CrossRef]
- Lim, J.H.; Cho, S.J.; Park, S.K.; Kim, J.; Cho, D.; Lee, W.J.; Kang, C.J. Stage-specific expression of two neighboring Crlz1 and IgJ genes during B cell development is regulated by their chromatin accessibility and histone acetylation. J. Immunol. 2006, 177, 5420–5429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Park, S.K.; Kim, H.G.; Cho, S.J.; Kim, J.; Kang, C.J. The HSS3/4 enhancer of Crlz1-IgJ locus is another target of EBF in the pre-B cell stage of B cell development. Immunol. Lett. 2006, 107, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.J.; Sheridan, C.; Koshland, M.E. A stage-specific enhancer of immunoglobulin J chain gene is induced by interleukin-2 in a presecretor B cell stage. Immunity 1998, 8, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.J.; Oh, U.; Koshland, M.E. Dynamic chromatin remodeling in the vicinity of J chain gene for the regulation of two stage-specific genes during B cell differentiation. Mol. Cells 2000, 10, 32–37. [Google Scholar] [CrossRef]
- Park, S.K.; Son, Y.; Kang, C.J. A strong promoter activity of pre-B cell stage-specific Crlz1 gene is caused by one distal LEF-1 and multiple proximal Ets sites. Mol. Cells 2011, 32, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.Y.; Park, S.K.; Yoo, H.W.; Pi, J.H.; Kang, C.J. Charged amino acid-rich leucine zipper-1 (Crlz-1) as a target of Wnt signaling pathway controls pre-B cell proliferation by affecting Runx/CBFβ-targeted VpreB and λ5 genes. J. Biol. Chem. 2016, 291, 15008–15019. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.Y.; Pi, J.H.; Park, S.; Kang, C.J. Crlz-1 controls germinal center reaction by relaying a Wnt signal to the Bcl-6 expression in centroblasts during humoral immune responses. J. Immunol. 2019, 203, 2630–2643. [Google Scholar] [CrossRef]
- Lim, J.H.; Choi, S.Y.; Yoo, H.W.; Cho, S.J.; Son, Y.; Kang, C.J. Crlz-1 is prominently expressed in spermatogonia and Sertoli cells during early testis development and in spermatids during late spermatogenesis. J. Histochem. Cytochem. 2013, 61, 522–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.K.; Lim, J.H.; Kang, C.J. Crlz1 activates transcription by mobilizing cytoplasmic CBFβ into the nucleus. Biochim. Biophys. Acta-Gene Regul. Mech. 2009, 1789, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, D.M.; Murphy, A.J.; Frendewey, D.; Gale, N.W.; Economides, A.N.; Auerbach, W.; Poueymirou, W.T.; Adams, N.C.; Rojas, J.; Yasenchak, J.; et al. High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat. Biotechnol. 2003, 21, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Shea, K.; Geijsen, N. Dissection of 6.5 dpc mouse embryos. JoVE 2007, 2, e160. [Google Scholar] [CrossRef] [PubMed]
- Pryor, S.E.; Massa, V.; Savery, D.; Greene, N.D.E.; Copp, A.J. Convergent extension analysis in mouse whole embryo culture. In Planar Cell Polarity: Methods in Molecular Biology (Methods and Protocols); Turksen, K., Ed.; Springer: New York, NY, USA, 2012; Volume 839, pp. 133–146. [Google Scholar]
- Rivron, N.; Fu, J. SnapShot: Embryo models. Stem Cell Rep. 2021, 16, 1142. [Google Scholar] [CrossRef]
- Chen, Y.J.C.; Wang, H.J.; Jauh, G.Y. Dual role of a SAS10/C1D family protein in ribosomal RNA gene expression and processing is essential for reproduction in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006408. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, Y.; Chen, F.; Huang, D.; Shi, H.; Lo, L.J.; Chen, J.; Peng, J. Sas10 controls ribosome biogenesis by stabilizing Mpp10 and delivering the Mpp10–Imp3–Imp4 complex to nucleolus. Nucleic Acids Res. 2019, 47, 2996–3012. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Robitaille, A.M.; Berndt, J.D.; Davidson, K.C.; Fischer, K.A.; Mathieu, J.; Potter, J.C.; Ruohola-Baker, H.; Moon, R.T. Wnt/β-catenin signaling promotes self-renewal and inhibits the primed state transition in naïve human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2016, 113, E6382–E6390. [Google Scholar] [CrossRef] [Green Version]
- Lien, W.; Fuchs, E. Wnt some lose some: Transcriptional governance of stem cells by Wnt/ß-catenin signaling. Genes Dev. 2014, 28, 1517–1532. [Google Scholar] [CrossRef] [Green Version]
- Kurek, D.; Neagu, A.; Tastemel, M.; Tuysuz, N.; Lehmann, J.; van de Werken, H.J.G.; Philipsen, S.; van der Linden, R.; Maas, A.; van IJcken, W.F.J.; et al. Endogenous WNT signals mediate BMP-induced and spontaneous differentiation of epiblast stem cells and human embryonic stem cells. Stem Cell Rep. 2015, 4, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Gong, X.; Miao, L.; Fang, J.; Zhang, J. Efficient induction of syncytiotrophoblast layer II cells from trophoblast stem cells by canonical Wnt signaling activation. Stem Cell Rep. 2017, 9, 2034–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Bruijn, M.; Dzierzak, E. Runx transcription factors in the development and function of the definitive hematopoietic system. Blood 2017, 129, 2061–2069. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.; Kamikubo, Y.; Liu, P. Role of RUNX1 in hematological malignancies. Blood 2017, 129, 2070–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tober, J.; Yzaguirre, A.D.; Piwarzyk, E.; Speck, N.A. Distinct temporal requirements for Runx1 in hematopoietic progenitors and stem cells. Development 2013, 140, 3765–3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Wang, Y.; Shao, J.; Wang, J.; Chen, W.; Li, Y. Cbfβ governs osteoblast−adipocyte lineage commitment through enhancing β-catenin signaling and suppressing adipogenesis gene expression. Proc. Natl. Acad. Sci. USA 2017, 114, 10119–10124. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.-Y.; Pi, J.-H.; Jeong, S.-E.; Kang, C.-J. Crlz-1 Homozygous Null Knockout Mouse Embryos Are Lethally Stopped in Their Early Development. Genes 2022, 13, 511. https://doi.org/10.3390/genes13030511
Choi S-Y, Pi J-H, Jeong S-E, Kang C-J. Crlz-1 Homozygous Null Knockout Mouse Embryos Are Lethally Stopped in Their Early Development. Genes. 2022; 13(3):511. https://doi.org/10.3390/genes13030511
Chicago/Turabian StyleChoi, Seung-Young, Joo-Hyun Pi, So-Eun Jeong, and Chang-Joong Kang. 2022. "Crlz-1 Homozygous Null Knockout Mouse Embryos Are Lethally Stopped in Their Early Development" Genes 13, no. 3: 511. https://doi.org/10.3390/genes13030511