
Genome-Wide Identification of CCD Gene Family in Six Cucurbitaceae Species and Its Expression Profiles in Melon




Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Physicochemical Properties of CCD Gene Family Members in Cucurbitaceae
2.2. Evolutionary Analysis
2.3. Chromosomal Localization
2.4. Gene Structure and Conserved Motifs Analysis
2.5. Cis-Acting Regulating Element Prediction
2.6. Expression Analysis
3. Results
3.1. Genome-Wide Identification of CCD Gene Family in Cucurbitaceae
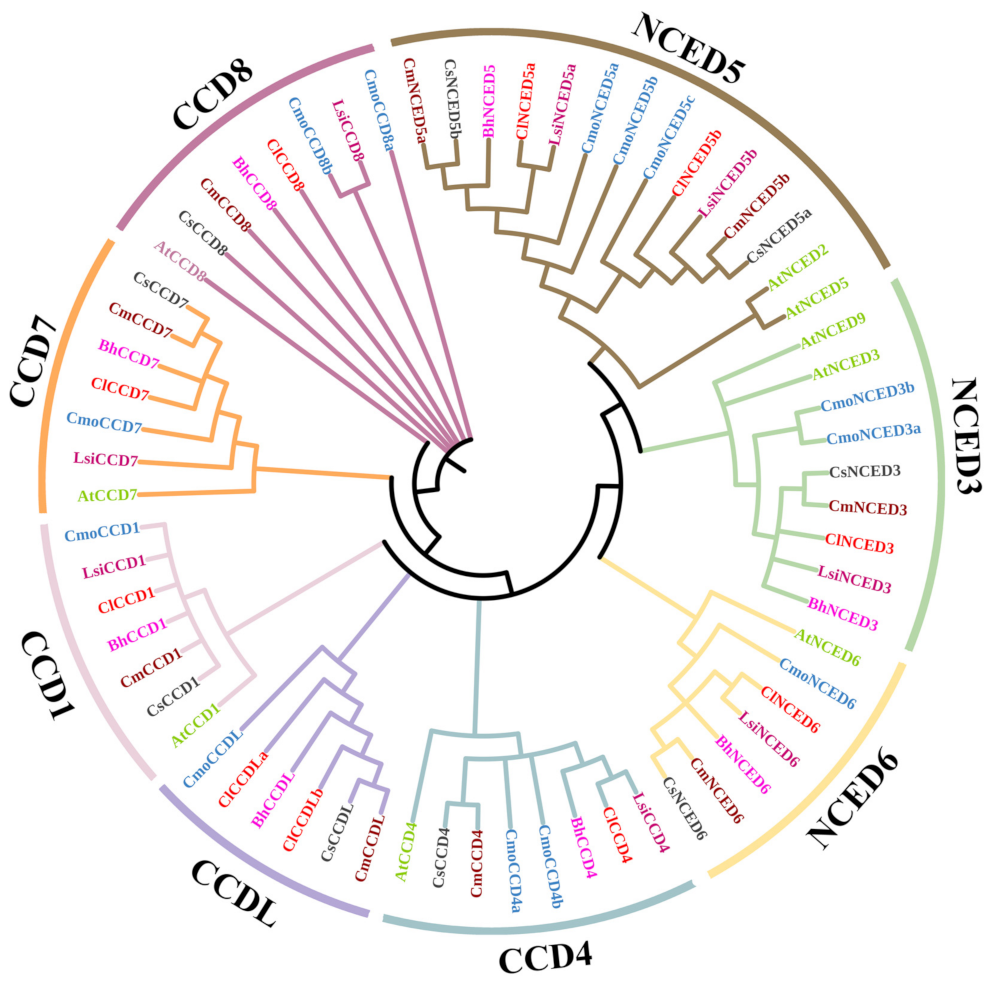
3.2. Evolutionary Analysis of CCD Gene Family in Cucurbitaceae
3.3. Members of CCDs Showed Variations in Chromosomal Localization in Cucurbitaceae
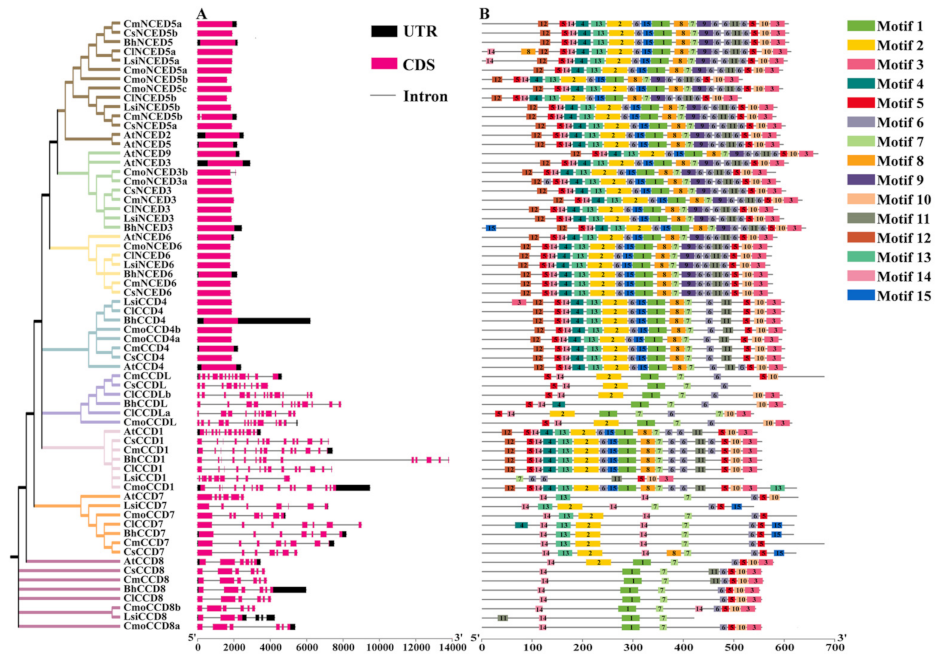
3.4. CCD Gene Family Presents Characteristic Gene Structure and Conserved Motifs in Cucurbitaceae
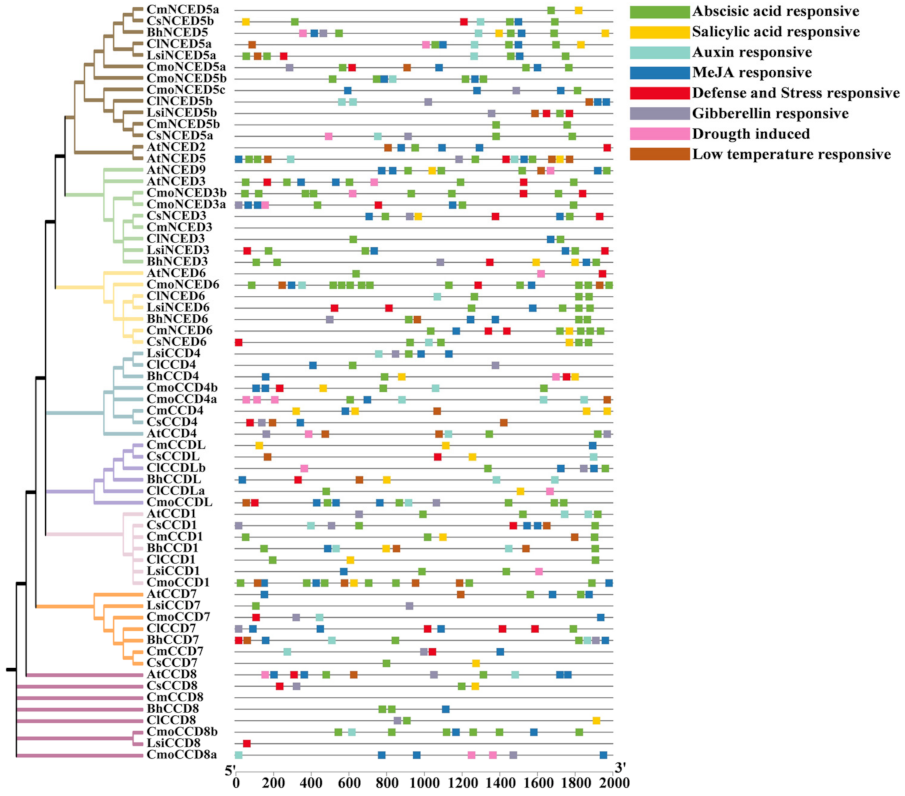
3.5. Cis-Acting Elements of CCDs Were Identified in Cucurbitaceae
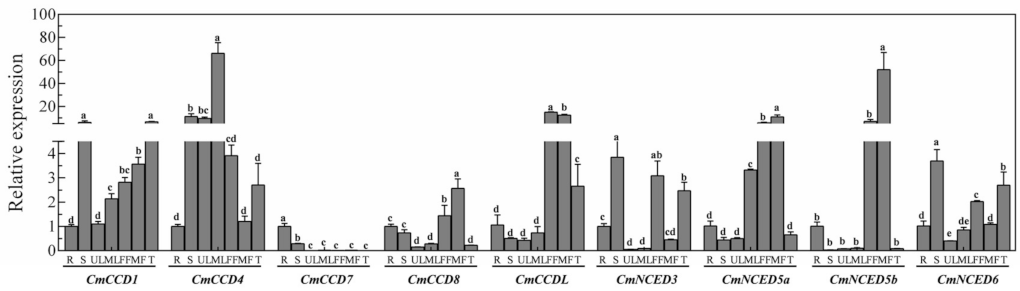
3.6. Expression Profiles of CCD Candidate Genes in Different Tissues of Melon
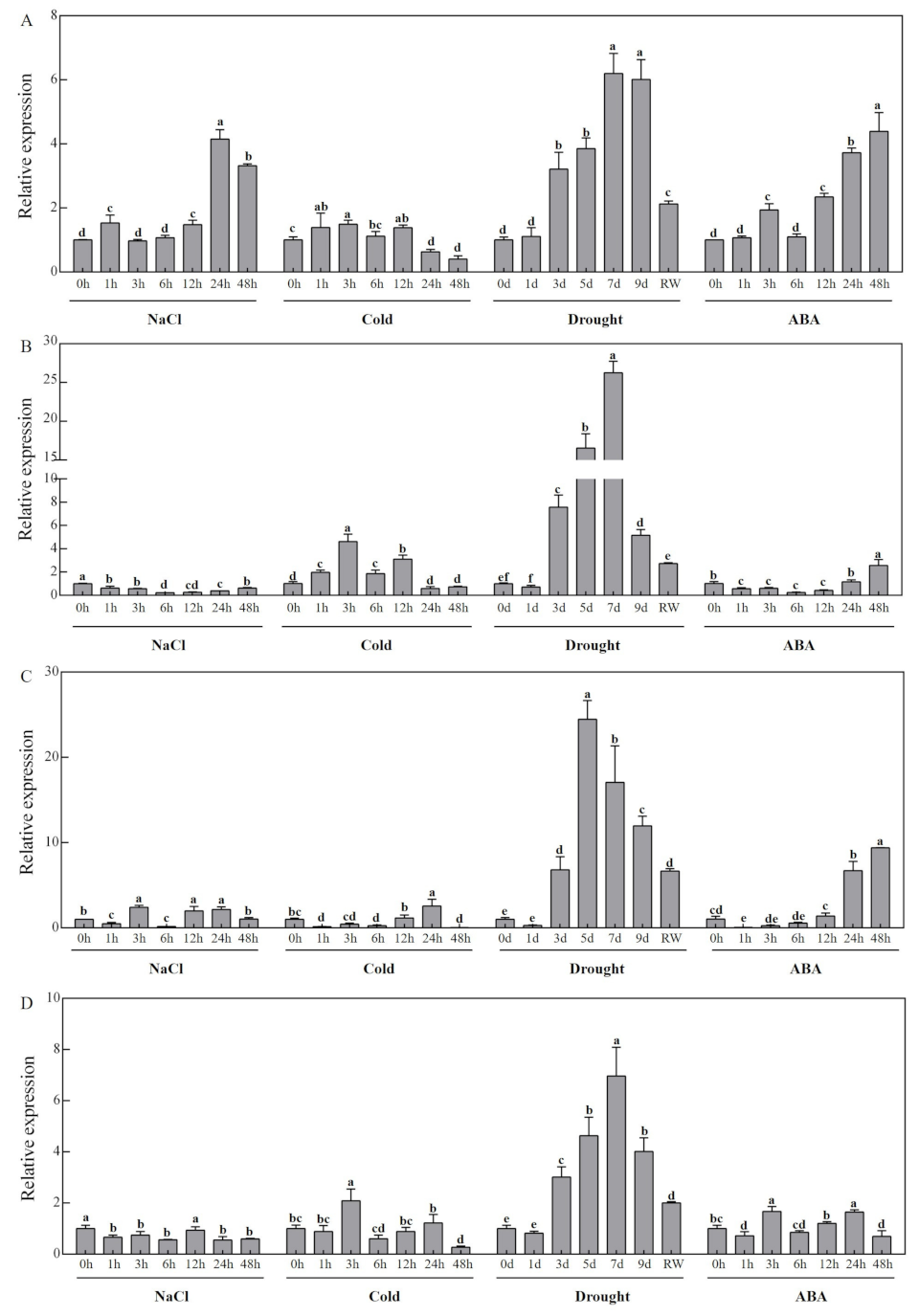
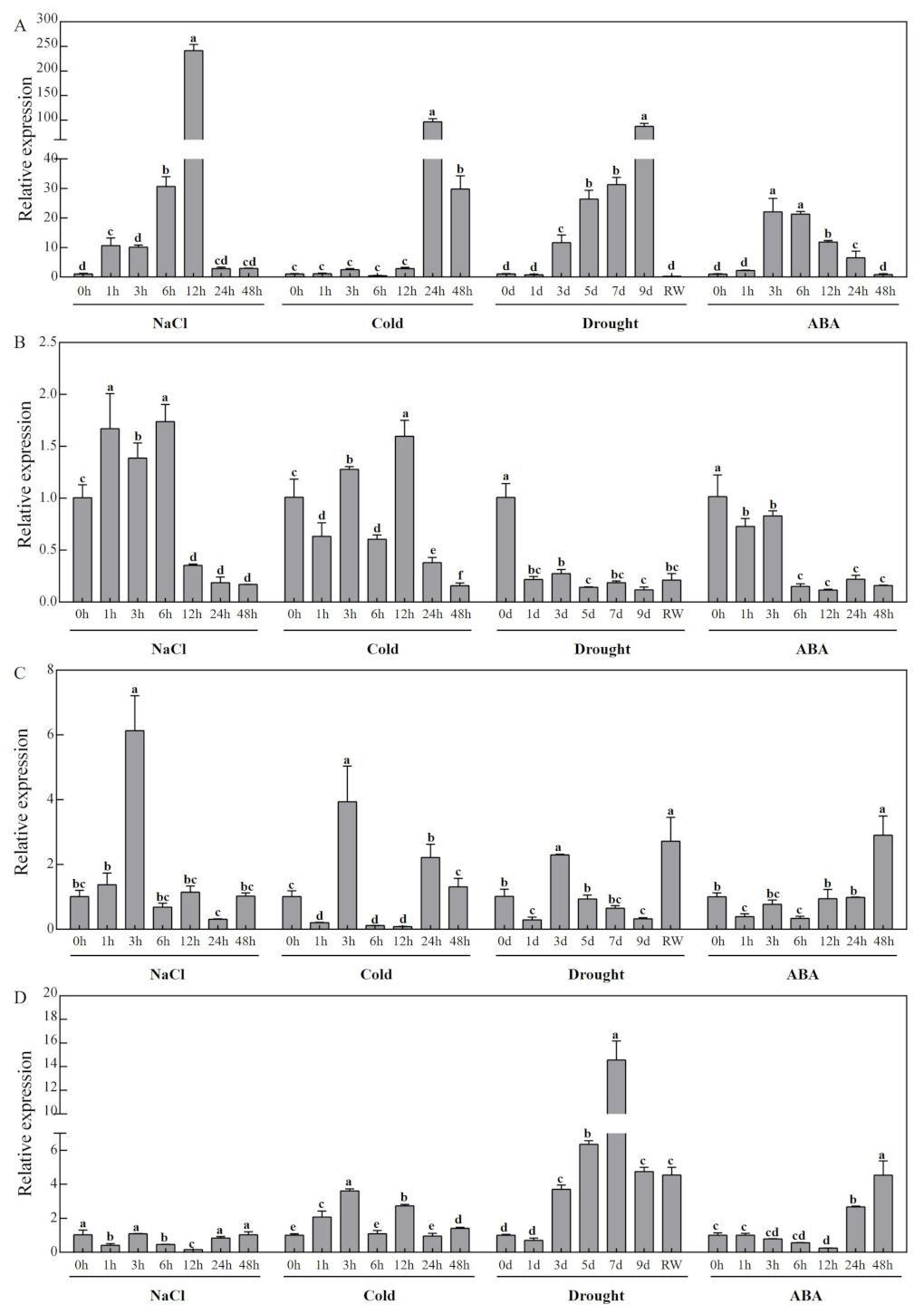
3.7. Expression Profiles of CCD Family Candidate Genes in Response to Abiotic Stress in Melon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou:, Q.; Li, Q.; Li, P.; Zhang, S.; Liu, C.; Jin, J.; Cao, P.; Yang, Y. Carotenoid Cleavage Dioxygenases: Identification, Expression, and Evolutionary Analysis of This Gene Family in Tobacco. Int. J. Mol. Sci. 2019, 20, 5796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahrazem, O.; Gomez-Gomez, L.; Rodrigo, M.J.; Avalos, J.; Limon, M.C. Carotenoid Cleavage Oxygenases From Microbes and Photosynthetic Organisms: Features and Functions. Int. J. Mol. Sci. 2016, 17, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, P.D.; Bramley, P.M. The biosynthesis and nutritional uses of carotenoids. Prog. Lipid. Res. 2004, 43, 228–265. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Bernaldo de Quirós, A.; Costa, H.S. Analysis of carotenoids in vegetable and plasma samples: A review. J. Food Compos. Anal. 2006, 19, 97–111. [Google Scholar] [CrossRef]
- Botella-Pavía, P.; Rodríguez-Concepción, M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol. Plant. 2010, 126, 369–381. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Shan, L.; Khin, N.C.; Pogson, B. Carotenoid Metabolism in Plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.C.; Schwartz, S.H.; Zeevaart, J.A.D.; McCarty, D.R. Genetic control of abscisic acid biosynthesis in maize. Proc. Natl. Acad. Sci. USA 1997, 94, 12235. [Google Scholar] [CrossRef] [Green Version]
- Ohmiya, A. Carotenoid cleavage dioxygenases and their apocarotenoid products in plants. Plant Biotechnol. 2009, 26, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Zhang, L.; Matsuta, A.; Matsutani, K.; Yamawaki, K.; Yahata, M.; Wahyudi, A.; Motohashi, R.; Kato, M. Enzymatic Formation of β-Citraurin from β-Cryptoxanthin and Zeaxanthin by Carotenoid Cleavage Dioxygenase4 in the Flavedo of Citrus Fruit. Plant Physiol. 2013, 163, 682–695. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.H.; Strack, D. Carotenoids and their cleavage products: Biosynthesis and functions. Nat. Prod. Rep. 2011, 28, 663–692. [Google Scholar] [CrossRef]
- Wei, Y.; Wan, H.; Wu, Z.; Wang, R.; Ruan, M.; Ye, Q.; Li, Z.; Zhou, G.; Yao, Z.; Yang, Y. A Comprehensive Analysis of Carotenoid Cleavage Dioxygenases Genes in Solanum Lycopersicum. Plant Mol. Biol. Rep. 2016, 34, 512–523. [Google Scholar] [CrossRef]
- Kim, Y.; Hwang, I.; Jung, H.-J.; Park, J.-I.; Kang, J.-G.; Nou, I.-S. Genome-Wide Classification and Abiotic Stress-Responsive Expression Profiling of Carotenoid Oxygenase Genes in Brassica rapa and Brassica oleracea. J. Plant Growth Regul. 2015, 35, 202–214. [Google Scholar] [CrossRef]
- Walter, M.H.; Floss, D.S.; Strack, D. Apocarotenoids: Hormones, mycorrhizal metabolites and aroma volatiles. Planta 2010, 232, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kloer, D.P.; Schulz, G.E. Structural and biological aspects of carotenoid cleavage. Cell Mol. Life Sci. 2006, 63, 2291–2303. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.C.; Molnar, P.; Schwab, W. Cloning and functional characterization of carotenoid cleavage dioxygenase 4 genes. J. Exp. Bot. 2009, 60, 3011–3022. [Google Scholar] [CrossRef] [Green Version]
- Bouvier, F.; Keller, Y.; d’Harlingue, A.; Camara, B. Xanthophyll biosynthesis: Molecular and functional characterization of carotenoid hydroxylases from pepper fruits (Capsicum annuum L.). Biochim. Biophys. Acta Lipids Lipid Metab. 1998, 1391, 320–328. [Google Scholar] [CrossRef]
- Enzell, C.R. Biodegradation of carotenoids—An important route to aroma compounds. Pure Appl. Chem. 1985, 57, 693–700. [Google Scholar] [CrossRef]
- Hou, X.; Rivers, J.; Leon, P.; McQuinn, R.P.; Pogson, B.J. Synthesis and Function of Apocarotenoid Signals in Plants. Trends Plant Sci. 2016, 21, 792–803. [Google Scholar] [CrossRef]
- McQuinn, R.P.; Giovannoni, J.J.; Pogson, B.J. More than meets the eye: From carotenoid biosynthesis, to new insights into apocarotenoid signaling. Curr. Opin. Plant Biol. 2015, 27, 172–179. [Google Scholar] [CrossRef]
- Tian, X.; Ji, J.; Wang, G.; Jin, C.; Guan, C.; Wu, D.; Li, Z. Cloning and Expression Analysis of 9-cis-Epoxycarotenoid Dioxygenase Gene 1 Involved in Fruit Maturation and Abiotic Stress Response in Lycium chinense. J. Plant Growth Regul. 2015, 34, 465–474. [Google Scholar] [CrossRef]
- Tian, X.; Ji, J.; Wang, G.; Jin, C.; Guan, C.; Wu, G. Molecular cloning and characterization of a novel carotenoid cleavage dioxygenase 1 from Lycium chinense. Biotechnol. Appl. Biochem. 2016, 62, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Underwood, B.A.; Auldridge, M.; Loucas, H.M.; Shibuya, K.; Schmelz, E.; Clark, D.G.; Klee, H.J. Circadian Regulation of the PhCCD1 Carotenoid Cleavage Dioxygenase Controls Emission of β-ionone, a Fragrance Volatile of Petunia Flowers. Plant Physiol. 2004, 136, 3504–3514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, B.C.; Joseph, L.M.; Deng, W.T.; Liu, L.; Li, Q.B.; Cline, K.; McCarty, D.R. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 2003, 35, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Verdejo, C.I.; Obrero, A.; Roman, B.; Gomez, P. Expression Profile of Carotenoid Cleavage Dioxygenase Genes in Summer Squash (Cucurbita pepo L.). Plant Foods Hum. Nutr. 2015, 70, 200–206. [Google Scholar] [CrossRef]
- Ibdah, M.; Azulay, Y.; Portnoy, V.; Wasserman, B.; Bar, E.; Meir, A.; Burger, Y.; Hirschberg, J.; Schaffer, A.A.; Katzir, N.; et al. Functional characterization of CmCCD1, a carotenoid cleavage dioxygenase from melon. Phytochemistry 2006, 67, 1579–1589. [Google Scholar] [CrossRef]
- Lv, P.; Na, L.; Liu, H.; Gu, H.; Zhao, W.E. Changes in carotenoid profiles and in the expression pattern of the genes in carotenoid metabolisms during fruit development and ripening in four watermelon cultivars. Food Chem. 2015, 174, 52–59. [Google Scholar] [CrossRef]
- Kong, Q.; Yuan, J.; Gao, L.; Liu, P.; Cao, L.; Huang, Y.; Zhao, L.; Lv, H.; Bie, Z. Transcriptional regulation of lycopene metabolism mediated by rootstock during the ripening of grafted watermelons. Food Chem. 2017, 214, 406–411. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wei, C.; Zhang, R.; Yang, X.; Zhu, C.; Li, H.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Comparative Analysis of Calcium-Dependent Protein Kinase in Cucurbitaceae and Expression Studies in Watermelon. Int. J. Mol. Sci. 2019, 20, 2527. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Pan, Y.; Zhi, C.; Zheng, Y.; Wang, X.; Li, X.; Cheng, Z. Genome-Wide Identification and Characterization of KNOTTED-Like Homeobox (KNOX) Homologs in Garlic (Allium sativum L.) and Their Expression Profilings Responding to Exogenous Cytokinin and Gibberellin. Int. J. Mol. Sci. 2021, 22, 9237. [Google Scholar] [CrossRef]
- Wang, Z.; Yadav, V.; Yan, X.; Cheng, D.; Wei, C.; Zhang, X. Systematic genome-wide analysis of the ethylene-responsive ACS gene family: Contributions to sex form differentiation and development in melon and watermelon. Gene 2021, 805, 145910. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Zhang, C.; Feng, J.; Feng, A.; You, C.; Ren, Y.; Wang, D.; Sun, T.; Su, Y.; Xu, L.; et al. Genome-wide identification, characterization and expression analysis of the carotenoid cleavage oxygenase (CCO) gene family in Saccharum. Plant Physiol. Biochem. 2021, 162, 196–210. [Google Scholar] [CrossRef]
- Zhang, X.H.; Liu, H.Q.; Guo, Q.W.; Zheng, C.F.; Li, C.S.; Xiang, X.M.; Zhao, D.F.; Liu, J.; Luo, J.; Zhao, D.K.; et al. Genome-wide identification, phylogenetic relationships, and expression analysis of the carotenoid cleavage oxygenase gene family in pepper. Genet. Mol. Res. 2016, 15, 15048695. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.T.; Jia, L.D.; Duan, M.Z.; Chen, X.; Qiao, C.L.; Ma, J.Q.; Zhang, C.; Jing, F.Y.; Zhang, S.S.; Yang, B.; et al. Genome-wide identification and expression profiling of the carotenoid cleavage dioxygenase (CCD) gene family in Brassica napus L. PLoS ONE 2020, 15, e0238179. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef]
- Cartharius, K.; Frech, K.; Grote, K.; Klocke, B.; Haltmeier, M.; Klingenhoff, A.; Frisch, M.; Bayerlein, M.; Werner, T. MatInspector and beyond: Promoter analysis based on transcription factor binding sites. Bioinformatics 2005, 21, 2933–2942. [Google Scholar] [CrossRef] [Green Version]
- Vallabhaneni, R.; Bradbury, L.M.; Wurtzel, E.T. The carotenoid dioxygenase gene family in maize, sorghum, and rice. Arch. Biochem. Biophys. 2010, 504, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Floss, D.S.; Walter, M.H. Role of carotenoid cleavage dioxygenase 1 (CCD1) in apocarotenoid biogenesis revisited. Plant Signal. Behav. 2009, 4, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Simkin, A.J.; Schwartz, S.H.; Auldridge, M.; Taylor, M.G.; Klee, H.J. The tomato carotenoid cleavage dioxygenase 1 genes contribute to the formation of the flavor volatiles β-ionone, pseudoionone, and geranylacetone. Plant J. 2004, 40, 882–892. [Google Scholar] [CrossRef]
- García-Limones, C.; Schnäbele, K.; Blanco-Portales, R.; Luz Bellido, M.; Caballero, J.L.; Schwab, W.; Muñoz-Blanco, J. Functional Characterization of FaCCD1: A Carotenoid Cleavage Dioxygenase from Strawberry Involved in Lutein Degradation during Fruit Ripening. J. Agric. Food Chem 2008, 56, 9277–9285. [Google Scholar] [CrossRef]
- Watanabe, K.; Oda-Yamamizo, C.; Sage-Ono, K.; Ohmiya, A.; Ono, M. Alteration of flower colour in Ipomoea nil through CRISPR/Cas9-mediated mutagenesis of carotenoid cleavage dioxygenase 4. Transgenic Res. 2018, 27, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-H.; Lim, S.-H.; Kim, J.K.; Jung, E.S.; John, K.M.M.; You, M.-K.; Ahn, S.-N.; Lee, C.H.; Ha, S.-H. In planta cleavage of carotenoids by Arabidopsis carotenoid cleavage dioxygenase 4 in transgenic rice plants. Plant Biotechnol. Rep. 2016, 10, 291–300. [Google Scholar] [CrossRef]
- Adami, M.; De Franceschi, P.; Brandi, F.; Liverani, A.; Giovannini, D.; Rosati, C.; Dondini, L.; Tartarini, S. Identifying a Carotenoid Cleavage Dioxygenase (ccd4) Gene Controlling Yellow/White Fruit Flesh Color of Peach. Plant Mol. Biol. Rep. 2013, 31, 1166–1175. [Google Scholar] [CrossRef]
- Hai, N.T.L.; Masuda, J.-I.; Miyajima, I.; Thien, N.Q.; Mojtahedi, N.; Hiramatsu, M.; Kim, J.-H.; Okubo, H. Involvement of Carotenoid Cleavage Dioxygenase 4 Gene in Tepal Color Change in Lilium brownii var. colchesteri. J. Jpn. Soc. Hortic. Sci. 2012, 81, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.D.; Kim, Y.-S.; Ryu, J.; Choi, H.-I.; Kim, S.W.; Kang, H.S.; Ahn, J.-W.; Kim, J.-B.; Kang, S.-Y.; Kim, S.H. Deletion of carotenoid cleavage dioxygenase 4a (CmCCD4a) and global up-regulation of plastid protein-coding genes in a mutant chrysanthemum cultivar producing yellow petals. Sci. Hortic. 2016, 212, 49–59. [Google Scholar] [CrossRef]
- Zhao, J.; Li, J.; Zhang, J.; Chen, D.; Zhang, H.; Liu, C.; Qin, G. Genome-Wide Identification and Expression Analysis of the Carotenoid Cleavage Oxygenase Gene Family in Five Rosaceae Species. Plant Mol. Biol. Rep. 2021, 1194, 1–13. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, Y.; Zhang, Y.; Guo, J.; Li, K.; Fu, W.; Jia, Z.; Li, W.; Tran, L.-S.P.; Jia, K.-P.; et al. Genome-wide identification, characterization and expression profiles of the CCD gene family in Gossypium species. 3 Biotech 2021, 11, 249. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef]
- Ledger, S.E.; Janssen, B.J.; Karunairetnam, S.; Wang, T.; Snowden, K.C. Modified Carotenoid Cleavage Dioxygenase8 expression correlates with altered branching in kiwifruit (Actinidia chinensis). New Phytol. 2010, 188, 803–813. [Google Scholar] [CrossRef]
- Vogel, J.T.; Walter, M.H.; Giavalisco, P.; Lytovchenko, A.; Kohlen, W.; Charnikhova, T.; Simkin, A.J.; Goulet, C.; Strack, D.; Bouwmeester, H.J.; et al. SlCCD7 controls strigolactone biosynthesis, shoot branching and mycorrhiza-induced apocarotenoid formation in tomato. Plant J. 2010, 61, 300–311. [Google Scholar] [CrossRef]
- Kulkarni, K.P.; Vishwakarma, C.; Sahoo, S.P.; Lima, J.M.; Nath, M.; Dokku, P.; Gacche, R.N.; Mohapatra, T.; Robin, S.; Sarla, N.; et al. A substitution mutation in OsCCD7 cosegregates with dwarf and increased tillering phenotype in rice. J. Genet. 2014, 93, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, T.; Xu, B.; Jia, L.; Xiao, B.; Liu, H.; Liu, L.; Yan, H.; Xia, Q. CRISPR/Cas9-Mediated Mutagenesis of Carotenoid Cleavage Dioxygenase 8 (CCD8) in Tobacco Affects Shoot and Root Architecture. Int. J. Mol. Sci. 2018, 19, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pages, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; et al. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic Acid: Emergence of a Core Signaling Network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.; Gao, Z.; Chen, L.; Wei, K.; Liu, J.; Fan, Y.; Davies, W.J.; Jia, W.; Zhang, J. Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit. J. Exp. Bot. 2007, 58, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A. Elucidation of the Indirect Pathway of Abscisic Acid Biosynthesis by Mutants, Genes, and Enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, P.; Sun, L.; Wang, Y.; Ji, K.; Sun, Y.; Dai, S.; Chen, P.; Duan, C.; Leng, P. Expression analysis of β-glucosidase genes that regulate abscisic acid homeostasis during watermelon (Citrullus lanatus) development and under stress conditions. J. Plant Physiol. 2012, 169, 78–85. [Google Scholar] [CrossRef]
- Ma, S.; Zhou, X.; Jahan, M.S.; Guo, S.; Tian, M.; Zhou, R.; Liu, H.; Feng, B.; Shu, S. Putrescine regulates stomatal opening of cucumber leaves under salt stress via the H2O2-mediated signaling pathway. Plant Physiol. Biochem. 2022, 170, 87–97. [Google Scholar] [CrossRef]
- Tong, S.M.; Xi, H.X.; Ai, K.J.; Hou, H.S. Overexpression of wheat TaNCED gene in Arabidopsis enhances tolerance to drought stress and delays seed germination. Biol. Plant. 2017, 61, 64–72. [Google Scholar] [CrossRef]
- Zhang, M.; Leng, P.; Zhang, G.; Li, X. Cloning and functional analysis of 9-cis-epoxycarotenoid dioxygenase (NCED) genes encoding a key enzyme during abscisic acid biosynthesis from peach and grape fruits. J. Plant Physiol. 2009, 166, 1241–1252. [Google Scholar] [CrossRef]
- Qin, X.; Zeevaart, J.A. Overexpression of a 9-cis-Epoxycarotenoid Dioxygenase Gene in Nicotiana plumbaginifolia Increases Abscisic Acid and Phaseic Acid Levels and Enhances Drought Tolerance. Plant Physiol. 2002, 128, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Ahrazem, O.; Rubio-Moraga, A.; Trapero, A.; Gomez-Gomez, L. Developmental and stress regulation of gene expression for a 9-cis-epoxycarotenoid dioxygenase, CstNCED, isolated from Crocus sativus stigmas. J. Exp. Bot. 2012, 63, 681–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuchi, S.; Kobayashi, M.; Taji, T.; Naramoto, M.; Seki, M.; Kato, T.; Tabata, S.; Kakubari, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J. 2001, 27, 325–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Gene Name | Gene ID | PI | Molecular Weight (kDa) | Chromosome Location | Gene Length (bp) | CDS Length (bp) | Number of Amino Acids (aa) | Grand Average of Hydropathicity | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| Citrullus lanatus | ClCCD1 | Cla97C09G166380 | 6.21 | 61.47 | Chr09: 3477850 .. 3484797 (+) | 6948 | 1644 | 547 | −0.291 | Cytoplasm |
| ClCCD4 | Cla97C07G137780 | 6.00 | 65.07 | Chr07: 25368259 .. 25370034 (+) | 1776 | 1776 | 591 | −0.234 | Chloroplast | |
| ClCCD7 | Cla97C06G111770 | 7.04 | 68.63 | Chr06: 2532539 .. 2541011 (−) | 8473 | 1833 | 610 | −0.215 | Chloroplast | |
| ClCCD8 | Cla97C08G155420 | 7.91 | 61.00 | Chr08: 23384307 .. 23388101 (−) | 3795 | 1644 | 547 | −0.235 | Chloroplast | |
| ClCCDLa | Cla97C11G221170 | 6.30 | 60.37 | Chr11: 27257473 .. 27262531 (+) | 5059 | 1596 | 531 | −0.301 | Cytoplasm | |
| ClCCDLb | Cla97C11G221180 | 6.84 | 67.14 | Chr11: 27264241 .. 27270176 (+) | 5936 | 1764 | 587 | −0.404 | Chloroplast | |
| ClNCED3 | Cla97C07G137260 | 5.81 | 64.04 | Chr07: 24786951 .. 24788687 (+) | 1737 | 1737 | 578 | −0.243 | Chloroplast | |
| ClNCED5a | Cla97C01G024630 | 6.28 | 67.74 | Chr01: 35670071 .. 35671885 (+) | 1815 | 1815 | 604 | −0.404 | Chloroplast | |
| ClNCED5b | Cla97C03G058030 | 5.61 | 56.93 | Chr03: 7238075 .. 7239598 (−) | 1524 | 1524 | 507 | −0.28 | Cytoplasm | |
| ClNCED6 | Cla97C06G117340 | 7.3 | 62.69 | Chr06: 9148622 .. 9150322 (+) | 1701 | 1701 | 566 | −0.279 | Cytoplasm | |
| Cucumis melo | CmCCD1 | MELO3C023555.2 | 6.01 | 61.28 | Chr01: 33650150 .. 33657132 (+) | 6983 | 1644 | 547 | −0.278 | Cytoplasm |
| CmCCD4 | MELO3C016224.2 | 6.1 | 65.08 | Chr07: 22480061 .. 22482158 (−) | 2098 | 1779 | 592 | −0.235 | Chloroplast | |
| CmCCD7 | MELO3C022291.2 | 7.82 | 75.47 | Chr11: 32532564 .. 32539629 (+) | 7066 | 2010 | 669 | −0.422 | Chloroplast | |
| CmCCD8 | MELO3C011142.2 | 6.13 | 61.14 | Chr03: 28383115 .. 28386694 (+) | 3580 | 1653 | 550 | −0.232 | Chloroplast | |
| CmCCDL | MELO3C003462.2 | 7.12 | 76.52 | Chr04: 1417853 .. 1422210 (−) | 4358 | 2010 | 669 | −0.404 | Chloroplast | |
| CmNCED3 | MELO3C027057.2 | 6.06 | 69.64 | Chr00: 1582539 .. 1584421 (+) | 1883 | 1881 | 626 | −0.214 | Chloroplast | |
| CmNCED5a | MELO3C007127.2 | 8.34 | 67.09 | Chr08: 991369 .. 993401 (−) | 2033 | 1800 | 599 | −0.389 | Chloroplast | |
| CmNCED5b | MELO3C002744.2 | 6.32 | 65.23 | Chr12: 21375227 .. 21377259 (−) | 2033 | 1728 | 575 | −0.378 | Mitochondrion | |
| CmNCED6 | MELO3C023086.2 | 7.98 | 63.02 | Chr05: 7293775 .. 7295481 (−) | 1707 | 1707 | 568 | −0.278 | Mitochondrion | |
| Cucumis sativus | CsCCD1 | CsaV3_7G031010 | 5.93 | 61.34 | Chr07: 19638561 .. 19646579 (+) | 7679 | 1644 | 547 | −0.276 | Cytoplasm |
| CsCCD4 | CsaV3_4G007180 | 5.83 | 65.04 | Chr04: 4915379 .. 4917916 (−) | 2538 | 1776 | 591 | −0.236 | Chloroplast | |
| CsCCD7 | CsaV3_6G008730 | 6.73 | 69.42 | Chr06: 7029451 .. 7035039 (+) | 5589 | 1845 | 614 | −0.345 | Chloroplast | |
| CsCCD8 | CsaV3_2G030870 | 6.17 | 60.76 | Chr02: 20280011 .. 20284196 (−) | 4186 | 1644 | 547 | −0.235 | Chloroplast | |
| CsCCDL | CsaV3_3G048570 | 8.92 | 59.86 | Chr03: 39663250 .. 39666931 (+) | 3682 | 1578 | 525 | −0.364 | Chloroplast | |
| CsNCED3 | CsaV3_4G007760 | 6.04 | 65.82 | Chr04: 5381592 .. 5384221 (−) | 2630 | 1785 | 594 | −0.273 | Chloroplast | |
| CsNCED5a | CsaV3_1G032340 | 6.82 | 66.92 | Chr01: 19294926 .. 19297672 (−) | 2747 | 1782 | 593 | −0.335 | Chloroplast | |
| CsNCED5b | CsaV3_6G051790 | 8.34 | 67.15 | Chr06: 30131435 .. 30133535 (+) | 2101 | 1803 | 600 | −0.377 | Chloroplast | |
| CsNCED6 | CsaV3_2G012080 | 8.78 | 63.02 | Chr02: 9534360 .. 9536321 (+) | 1962 | 1707 | 568 | −0.298 | Chloroplast | |
| Cucurbita moschata | CmoCCD1 | CmoCh17G009320 | 6.82 | 69.02 | Chr17: 8291575 .. 8300490 (−) | 8916 | 1848 | 615 | −0.186 | Cytoplasm |
| CmoCCD4a | CmoCh04G006500 | 5.7 | 64.61 | Chr04: 3223403 .. 3225163 (−) | 1761 | 1761 | 586 | −0.222 | Chloroplast | |
| CmoCCD4b | CmoCh16G005460 | 6.05 | 65.30 | Chr16: 2645574 .. 2647358 (+) | 1785 | 1785 | 594 | −0.231 | Chloroplast | |
| CmoCCD7 | CmoCh17G005060 | 6.45 | 68.79 | Chr17: 4737408 .. 4741956 (−) | 4549 | 1848 | 615 | −0.258 | Chloroplast | |
| CmoCCD8a | CmoCh05G006180 | 8.73 | 60.84 | Chr05: 3082399 .. 3087453 (−) | 5055 | 1644 | 547 | −0.221 | Chloroplast | |
| CmoCCD8b | CmoCh12G002040 | 6.58 | 59.53 | Chr12: 1340192 .. 1343166 (−) | 2975 | 1608 | 535 | −0.23 | Chloroplast | |
| CmoCCDL | CmoCh01G014860 | 6.59 | 68.36 | Chr01: 11513752 .. 11518925 (−) | 5174 | 1821 | 606 | −0.293 | Chloroplast | |
| CmoNCED3a | CmoCh04G006910 | 6.49 | 64.70 | Chr04: 3434234 .. 3435985 (−) | 1752 | 1752 | 583 | −0.254 | Mitochondrion | |
| CmoNCED3b | CmoCh16G004950 | 5.61 | 63.95 | Chr16: 2388840 .. 2390824 (+) | 1985 | 1725 | 574 | −0.291 | Chloroplast | |
| CmoNCED5a | CmoCh03G013970 | 6.58 | 65.79 | Chr03: 10201389 .. 10203155 (+) | 1767 | 1767 | 588 | −0.389 | Chloroplast | |
| CmoNCED5b | CmoCh07G001020 | 5.86 | 57.28 | Chr07: 595470 .. 596999 (−) | 1530 | 1530 | 509 | −0.314 | Cytoplasm | |
| CmoNCED5C | CmoCh13G005570 | 6.06 | 65.71 | Chr13: 6389844 .. 6391610 (−) | 1767 | 1767 | 588 | −0.287 | Chloroplast | |
| CmoNCED6 | CmoCh17G001180 | 7.26 | 62.53 | Chr17: 638459 .. 640162 (−) | 1704 | 1704 | 567 | −0.226 | Chloroplast | |
| Lagenaria siceraria | LsiCCD1 | Lsi02G025710 | 6.33 | 58.89 | Chr02: 32164396 .. 32169161 (+) | 4766 | 1536 | 511 | 0.037 | Plasma membrane |
| LsiCCD4 | Lsi07G007810 | 5.91 | 65.09 | Chr07: 8595598 .. 8597373 (−) | 1776 | 1776 | 591 | −0.207 | Chloroplast | |
| LsiCCD7 | Lsi09G016990 | 6.3 | 59.89 | Chr09: 25505283 .. 25512032 (+) | 6750 | 1596 | 531 | −0.212 | Cytoplasm | |
| LsiCCD8 | Lsi08G010850 | 8.34 | 46.52 | Chr08: 19415522 .. 19419519 (−) | 3998 | 1245 | 414 | −0.161 | Chloroplast | |
| LsiNCED3 | Lsi07G008450 | 5.94 | 65.41 | Chr07: 9468950 .. 9470722 (−) | 1773 | 1773 | 590 | −0.265 | Chloroplast | |
| LsiNCED5a | Lsi01G001400 | 6.65 | 66.92 | Chr01: 1451654 .. 1453447 (−) | 1794 | 1794 | 597 | −0.385 | Chloroplast | |
| LsiNCED5b | Lsi02G008960 | 6.22 | 65.05 | Chr02: 8564357 .. 8566090 (+) | 1734 | 1734 | 577 | −0.325 | Chloroplast | |
| LsiNCED6 | Lsi09G011600 | 8.16 | 62.28 | Chr09: 18638525 .. 18640216 (−) | 1692 | 1692 | 563 | −0.243 | Cytoplasm | |
| Benincasa hispida | BhCCD1 | Bhi09G000536 | 6.16 | 61.37 | Chr09: 13766074 .. 13779057 (−) | 12984 | 1644 | 547 | −0.266 | Cytoplasm |
| BhCCD4 | Bhi01G002364 | 6.05 | 64.50 | Chr01: 75445296 .. 75451135 (−) | 5840 | 1764 | 587 | −0.228 | Chloroplast | |
| BhCCD7 | Bhi12G001994 | 8.54 | 68.73 | Chr12: 71046834 .. 71054528 (+) | 7695 | 1830 | 609 | −0.192 | Chloroplast | |
| BhCCD8 | Bhi04G000698 | 8.17 | 60.39 | Chr04: 21509866 .. 21515484 (+) | 5619 | 1632 | 543 | −0.266 | Chloroplast | |
| BhCCDL | Bhi09G002182 | 6.26 | 67.97 | Chr09: 70181013 .. 70188430 (+) | 7418 | 1785 | 594 | −0.35 | Chloroplast | |
| BhNCED3 | Bhi01G002426 | 6.38 | 70.01 | Chr01: 77692588 .. 77694885 (−) | 2298 | 1902 | 633 | −0.214 | Chloroplast | |
| BhNCED5 | Bhi03G000083 | 8.53 | 67.40 | Chr03: 2257210 .. 2259286 (+) | 2077 | 1803 | 600 | −0.387 | Chloroplast | |
| BhNCED6 | Bhi12G001328 | 7.24 | 62.85 | Chr12: 49163168 .. 49165223 (+) | 2056 | 1707 | 568 | −0.271 | Chloroplast |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, D.; Wang, Z.; Li, S.; Zhao, J.; Wei, C.; Zhang, Y. Genome-Wide Identification of CCD Gene Family in Six Cucurbitaceae Species and Its Expression Profiles in Melon. Genes 2022, 13, 262. https://doi.org/10.3390/genes13020262
Cheng D, Wang Z, Li S, Zhao J, Wei C, Zhang Y. Genome-Wide Identification of CCD Gene Family in Six Cucurbitaceae Species and Its Expression Profiles in Melon. Genes. 2022; 13(2):262. https://doi.org/10.3390/genes13020262
Chicago/Turabian StyleCheng, Denghu, Zhongyuan Wang, Shiyu Li, Juan Zhao, Chunhua Wei, and Yong Zhang. 2022. "Genome-Wide Identification of CCD Gene Family in Six Cucurbitaceae Species and Its Expression Profiles in Melon" Genes 13, no. 2: 262. https://doi.org/10.3390/genes13020262