
Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
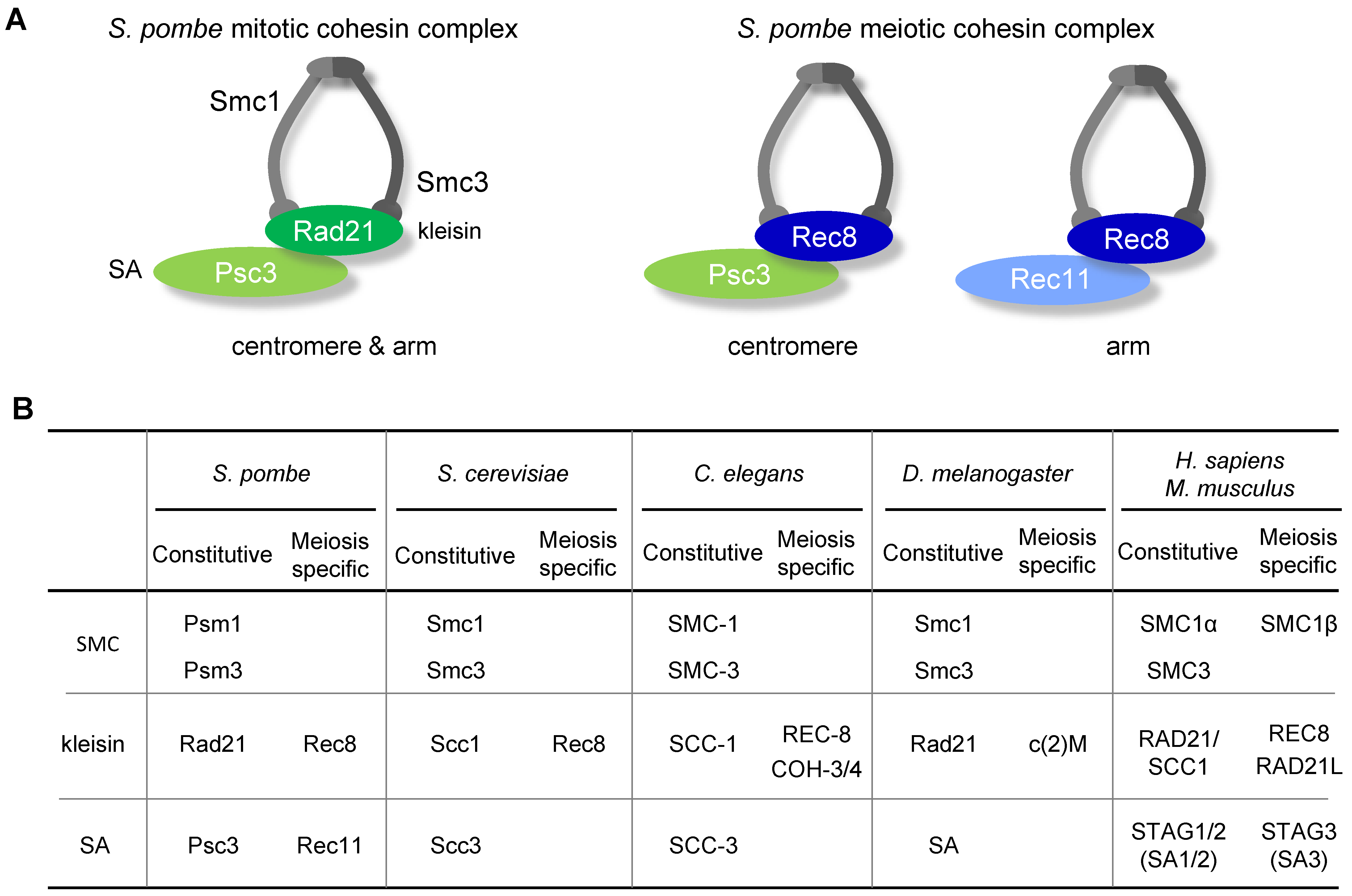
2. Components of Cohesin Complexes
3. Roles of Meiotic Cohesin in Chromosomal Events
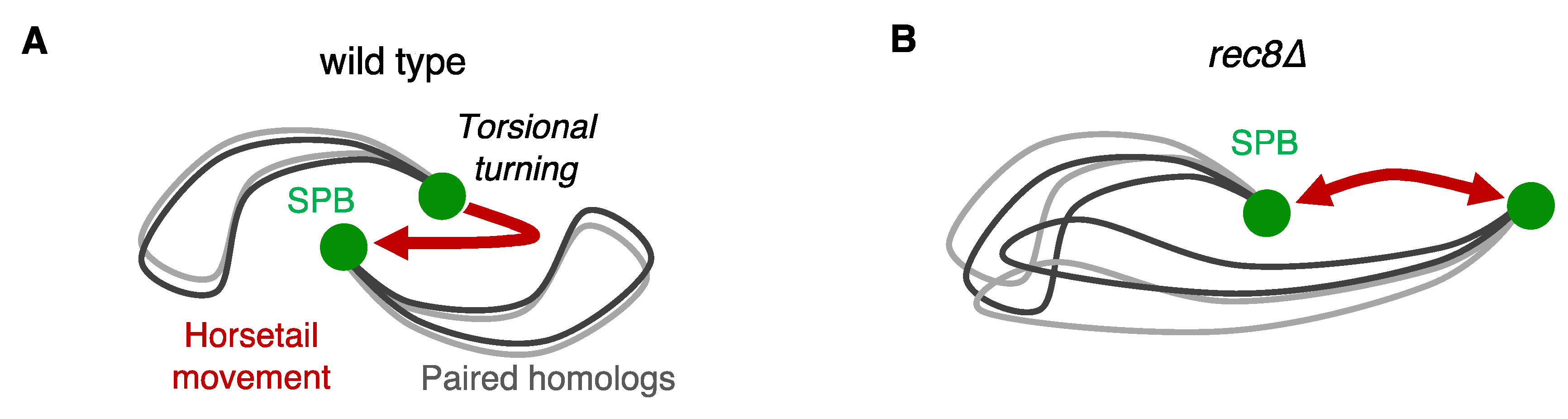
3.1. Pairing of Homologous Chromosomes
3.2. Regulation of Recombination Bias between Homologous Chromosomes and Sister Chromatids
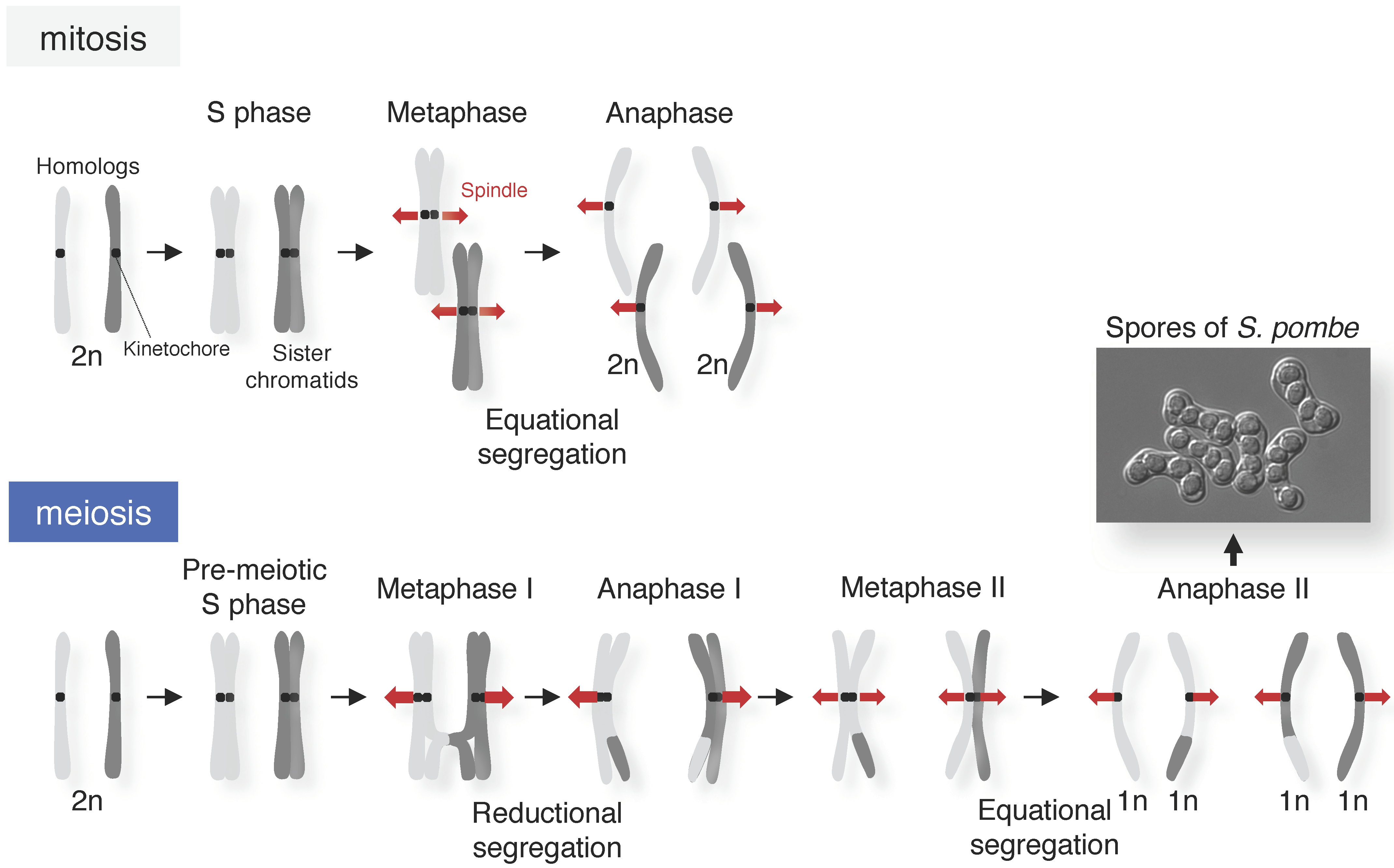
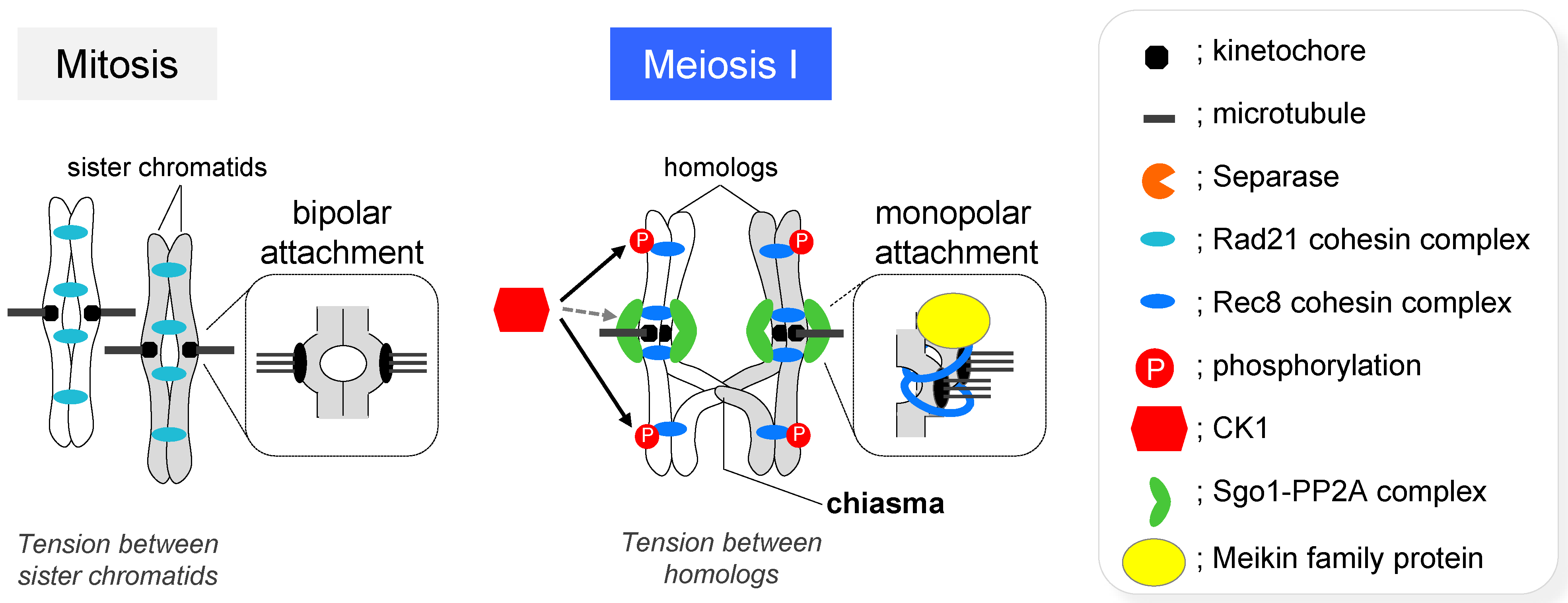
3.3. Segregation of Homologous Chromosomes and Sister Chromatids
4. Roles of Meiotic Cohesin in Chromosome Architecture
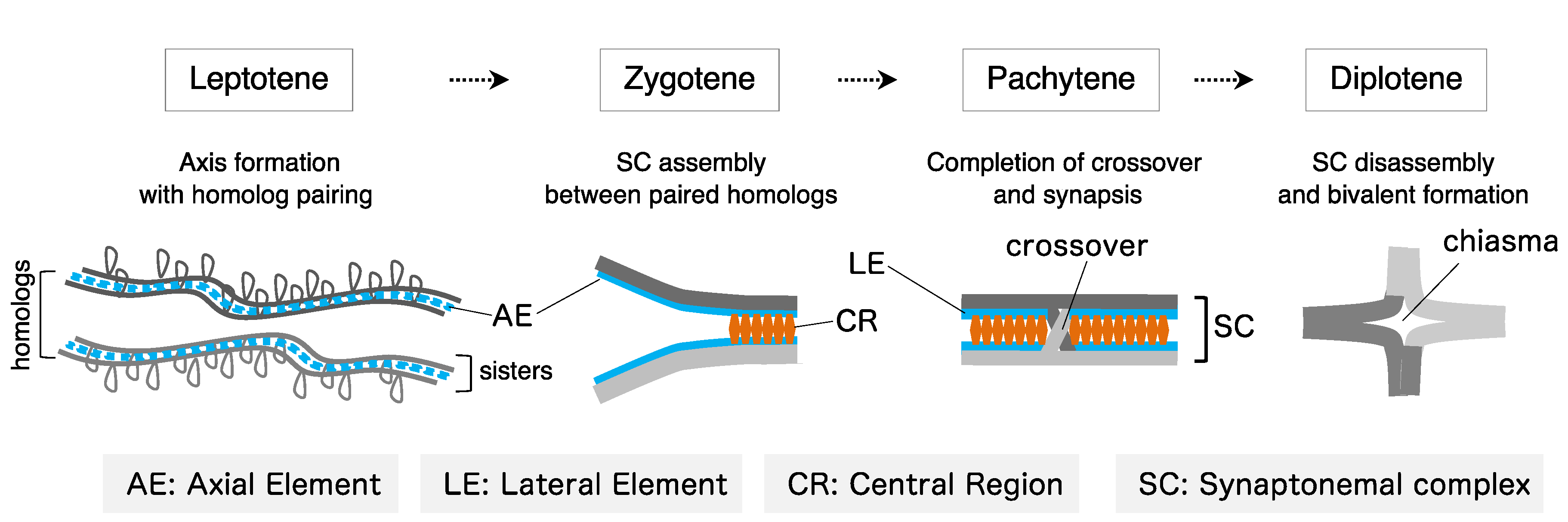
4.1. Formation of Axial Elements
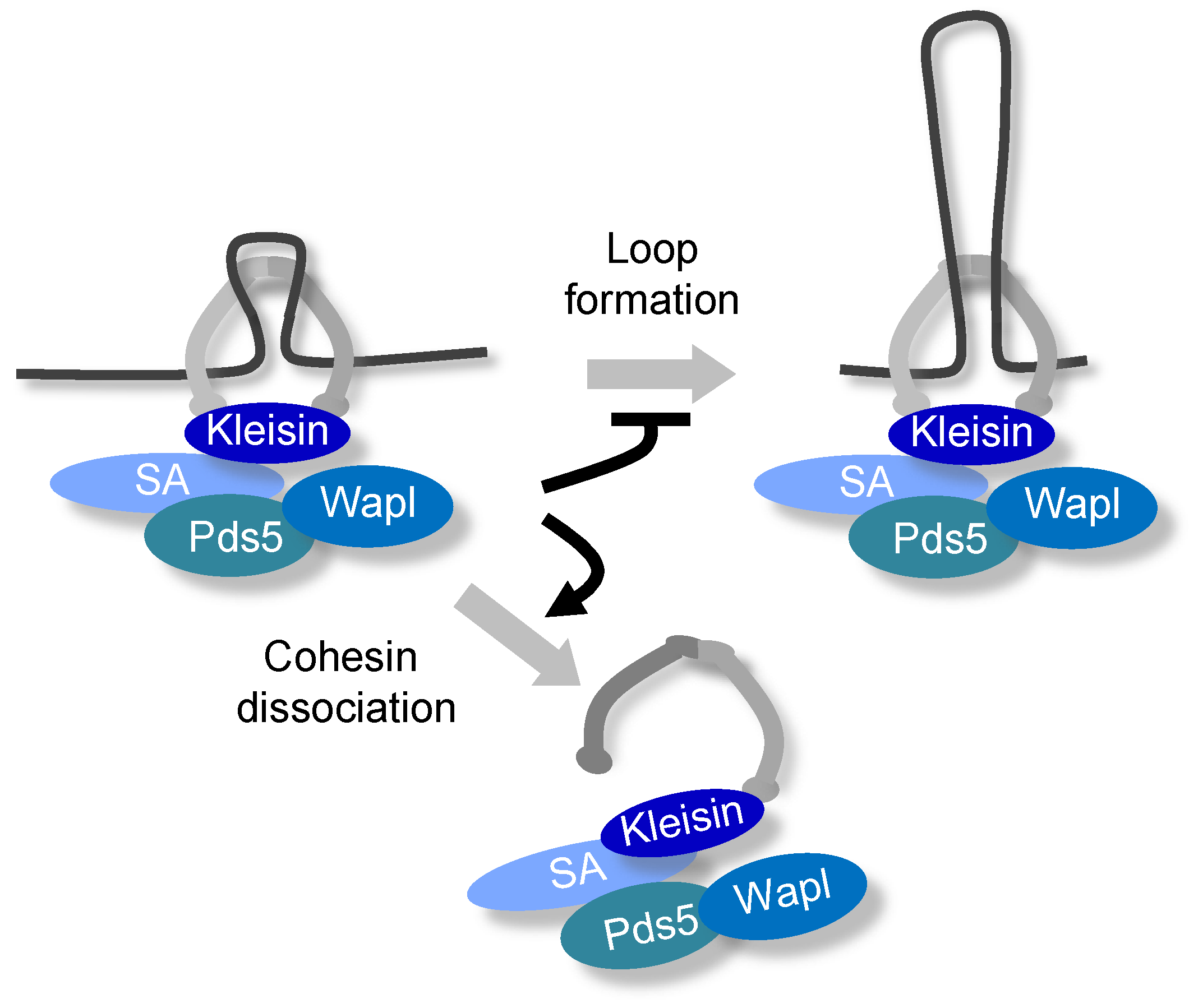
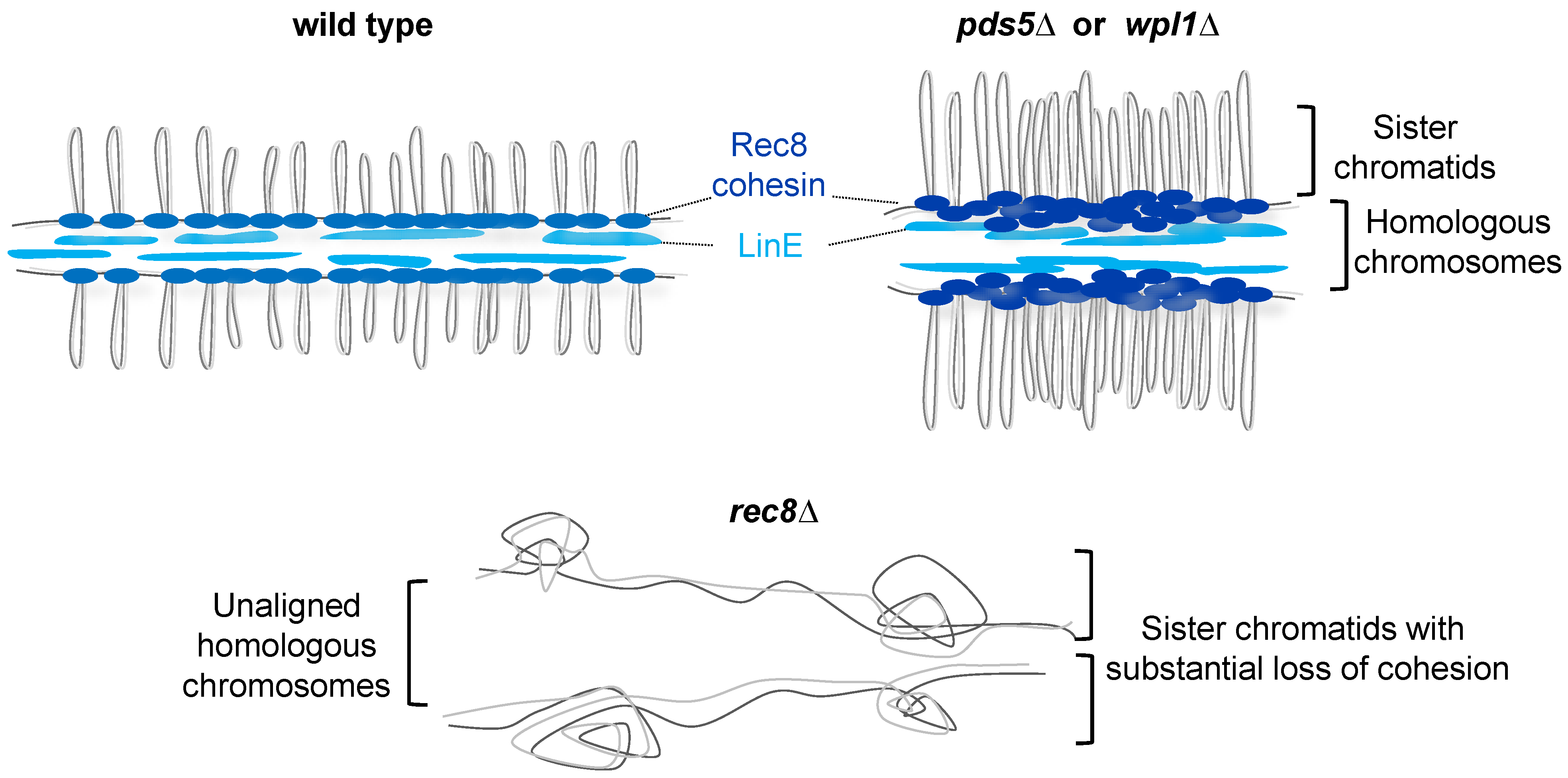
4.2. Formation of the Axis-Loop Chromatin Structure
5. Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nasmyth, K. Segregating sister genomes: The molecular biology of chromosome separation. Science 2002, 297, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlmann, F. Chromosome cohesion and segregation in mitosis and meiosis. Curr. Opin. Cell Biol. 2001, 13, 754–761. [Google Scholar] [CrossRef]
- Petronczki, M.; Siomos, M.F.; Nasmyth, K. Un menage a quatre: The molecular biology of chromosome segregation in meiosis. Cell 2003, 112, 423–440. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Orr-Weaver, T.L. The molecular basis of sister-chromatid cohesion. Annu. Rev. Cell Dev. Biol. 2001, 17, 753–777. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.I.; Hassold, T.J.; Hunt, P.A. Human aneuploidy: Mechanisms and new insights into an age-old problem. Nat. Rev. Genet. 2012, 13, 493–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, A.; Schuh, M. Mechanisms of Aneuploidy in Human Eggs. Trends Cell Biol. 2017, 27, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Kleckner, N. Chiasma formation: Chromatin/axis interplay and the role(s) of the synaptonemal complex. Chromosoma 2006, 115, 175–194. [Google Scholar] [CrossRef]
- Grey, C.; de Massy, B. Chromosome Organization in Early Meiotic Prophase. Front. Cell Dev. Biol. 2021, 9, 688878. [Google Scholar] [CrossRef] [PubMed]
- Gerton, J.L.; Hawley, R.S. Homologous chromosome interactions in meiosis: Diversity amidst conservation. Nat. Rev. Genet. 2005, 6, 477–487. [Google Scholar] [CrossRef]
- Barzel, A.; Kupiec, M. Finding a match: How do homologous sequences get together for recombination? Nat. Rev. Genet. 2008, 9, 27–37. [Google Scholar] [CrossRef]
- Zickler, D.; Kleckner, N. Recombination, Pairing, and Synapsis of Homologs during Meiosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a016626. [Google Scholar] [CrossRef] [Green Version]
- Niwa, O.; Shimanuki, M.; Miki, F. Telomere-led bouquet formation facilitates homologous chromosome pairing and restricts ectopic interaction in fission yeast meiosis. EMBO J. 2000, 19, 3831–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherthan, H.; Bähler, J.; Kohli, J. Dynamics of chromosome organization and pairing during meiotic prophase in fission yeast. J. Cell Biol. 1994, 127, 273–285. [Google Scholar] [CrossRef]
- Yamamoto, A.; West, R.R.; McIntosh, J.R.; Hiraoka, Y. A cytoplasmic dynein heavy chain is required for oscillatory nuclear movement of meiotic prophase and efficient meiotic recombination in fission yeast. J. Cell Biol. 1999, 145, 1233–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, F.; Mahr, P.; Galova, M.; Buonomo, S.B.C.; Michaelis, C.; Nairz, K.; Nasmyth, K. A central role for cohesins in sister chromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell 1999, 98, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Pasierbek, P.; Jantsch, M.; Melcher, M.; Schleiffer, A.; Schweizer, D.; Loidl, J. A Caenorhabditis elegans cohesion protein with functions in meiotic chromosome pairing and disjunction. Genes Dev. 2001, 15, 1349–1360. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Dong, F.; Edelmann, R.E.; Makaroff, C.A. The Arabidopsis SYN1 cohesin protein is required for sister chromatid arm cohesion and homologous chromosome pairing. J. Cell Sci. 2003, 116, 2999–3007. [Google Scholar] [CrossRef] [Green Version]
- Golubovskaya, I.N.; Hamant, O.; Timofejeva, L.; Wang, C.J.; Braun, D.; Meeley, R.; Cande, W.Z. Alleles of afd1 dissect REC8 functions during meiotic prophase I. J. Cell Sci. 2006, 119, 3306–3315. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Beasley, M.D.; Warren, W.D.; van der Horst, G.T.; McKay, M.J. Absence of mouse REC8 cohesin promotes synapsis of sister chromatids in meiosis. Dev. Cell 2005, 8, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, K.; Kim, J.; Shibuya, H.; Hernandez-Hernandez, A.; Suzuki, A.; Fukagawa, T.; Shioi, G.; Kiyonari, H.; Li, X.C.; Schimenti, J.; et al. Meiosis-specific cohesin mediates homolog recognition in mouse spermatocytes. Genes Dev. 2014, 28, 594–607. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, T.; Fukuda, N.; Agostinho, A.; Hernandez-Hernandez, A.; Kouznetsova, A.; Hoog, C. STAG3-mediated stabilization of REC8 cohesin complexes promotes chromosome synapsis during meiosis. EMBO J. 2014, 33, 1243–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, J.; Hwang, G.; Jacob, J.; Sapp, N.; Bedigian, R.; Oka, K.; Overbeek, P.; Murray, S.; Jordan, P.W. Meiosis-specific cohesin component, Stag3 is essential for maintaining centromere chromatid cohesion, and required for DNA repair and synapsis between homologous chromosomes. PLoS Genet. 2014, 10, e1004413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llano, E.; Gomez, H.L.; Garcia-Tunon, I.; Sanchez-Martin, M.; Caburet, S.; Barbero, J.L.; Schimenti, J.C.; Veitia, R.A.; Pendas, A.M. STAG3 is a strong candidate gene for male infertility. Hum. Mol. Genet. 2014, 23, 3421–3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winters, T.; McNicoll, F.; Jessberger, R. Meiotic cohesin STAG3 is required for chromosome axis formation and sister chromatid cohesion. EMBO J. 2014, 33, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, Y.; Horisawa-Takada, Y.; Inoue, E.; Tani, N.; Shibuya, H.; Fujimura, S.; Kariyazono, R.; Sakata, T.; Ohta, K.; Araki, K.; et al. Meiotic cohesins mediate initial loading of HORMAD1 to the chromosomes and coordinate SC formation during meiotic prophase. PLoS Genet. 2020, 16, e1009048. [Google Scholar] [CrossRef]
- Loidl, J.S. pombe linear elements: The modest cousins of synaptonemal complexes. Chromosoma 2006, 115, 260–271. [Google Scholar] [CrossRef]
- Bähler, J.; Wyler, T.; Loidl, J.; Kohli, J. Unusual nuclear structures in meiosis prophase of fission yeast: A cytological analysis. J. Cell Biol. 1993, 121, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; Rozalen, A.E.; Moreno, S.; Smith, G.R.; Martin-Castellanos, C. Rec25 and Rec27, novel linear-element components, link cohesin to meiotic DNA breakage and recombination. Curr. Biol. 2008, 18, 849–854. [Google Scholar] [CrossRef]
- Estreicher, A.; Lorenz, A.; Loidl, J. Mug20, a novel protein associated with linear elements in fission yeast meiosis. Curr. Genet. 2012, 58, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, A.; Wells, J.L.; Pryce, D.W.; Novatchkova, M.; Eisenhaber, F.; McFarlane, R.J.; Loidl, J.S. pombe meiotic linear elements contain proteins related to synaptonemal complex components. J. Cell Sci. 2004, 117, 3343–3351. [Google Scholar] [CrossRef] [Green Version]
- Olson, L.W.; Eden, U.; Egel-Mitani, M.; Egel, R. Asynaptic meiosis in fission yeast? Hereditas 1978, 89, 189–199. [Google Scholar] [CrossRef]
- Molnar, M.; Bahler, J.; Sipiczki, M.; Kohli, J. The rec8 gene of Schizosaccharomyces pombe is involved in linear element formation, chromosome pairing and sister-chromatid cohesion during meiosis. Genetics 1995, 141, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Schalbetter, S.A.; Fudenberg, G.; Baxter, J.; Pollard, K.S.; Neale, M.J. Principles of meiotic chromosome assembly revealed in S. cerevisiae. Nat. Commun. 2019, 10, 4795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, H.; Scolari, V.F.; Agier, N.; Piazza, A.; Thierry, A.; Mercy, G.; Descorps-Declere, S.; Lazar-Stefanita, L.; Espeli, O.; Llorente, B.; et al. Characterizing meiotic chromosomes’ structure and pairing using a designer sequence optimized for Hi-C. Mol. Syst. Biol. 2018, 14, e8293. [Google Scholar] [CrossRef]
- Sakuno, T.; Tashiro, S.; Tanizawa, H.; Iwasak, O.; Ding, D.; Haraguchi, T.; Noma, K.; Hiraoka, Y. Rec8 cohesin-mediated loop-axis chromatin architecture is required for meiotic recombination. BioRxiv 2021. [Google Scholar] [CrossRef]
- Chatzidaki, E.E.; Powell, S.; Dequeker, B.J.H.; Gassler, J.; Silva, M.C.C.; Tachibana, K. Ovulation suppression protects against chromosomal abnormalities in mouse eggs at advanced maternal age. Curr. Biol. 2021, 31, 4038–4051.e4037. [Google Scholar] [CrossRef] [PubMed]
- Nasmyth, K.; Haering, C.H. The structure and function of SMC and kleisin complexes. Annu. Rev. Biochem. 2005, 74, 595–648. [Google Scholar] [CrossRef] [PubMed]
- Haering, C.H.; Nasmyth, K. Building and breaking bridges between sister chromatids. Bioessays 2003, 25, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Onn, I.; Heidinger-Pauli, J.M.; Guacci, V.; Unal, E.; Koshland, D.E. Sister chromatid cohesion: A simple concept with a complex reality. Annu. Rev. Cell Dev. Biol. 2008, 24, 105–129. [Google Scholar] [CrossRef] [Green Version]
- Mehta, G.D.; Kumar, R.; Srivastava, S.; Ghosh, S.K. Cohesin: Functions beyond sister chromatid cohesion. FEBS Lett. 2013, 587, 2299–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasmyth, K. Disseminating the genome: Joining, resolving, and separating sister chromatids during mitosis and meiosis. Annu. Rev. Genet. 2001, 35, 673–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishiguro, K.I. The cohesin complex in mammalian meiosis. Genes Cells 2019, 24, 6–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.Q.; Okamasa, K.; Katou, Y.; Oya, E.; Nakayama, J.I.; Chikashige, Y.; Shirahige, K.; Haraguchi, T.; Hiraoka, Y. Chromosome-associated RNA-protein complexes promote pairing of homologous chromosomes during meiosis in Schizosaccharomyces pombe. Nat. Commun. 2019, 10, 5598. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Hiraoka, Y. How do meiotic chromosomes meet their homologous partners?: Lessons from fission yeast. Bioessays 2001, 23, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, Y. Phase separation drives pairing of homologous chromosomes. Curr. Genet. 2020, 66, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Chikashige, Y.; Ding, D.Q.; Funabiki, H.; Haraguchi, T.; Mashiko, S.; Yanagida, M.; Hiraoka, Y. Telomere-led premeiotic chromosome movement in fission yeast. Science 1994, 264, 270–273. [Google Scholar] [CrossRef]
- Ding, D.Q.; Chikashige, Y.; Haraguchi, T.; Hiraoka, Y. Oscillatory nuclear movement in fission yeast meiotic prophase is driven by astral microtubules, as revealed by continuous observation of chromosomes and microtubules in living cells. J. Cell Sci. 1998, 111 Pt 6, 701–712. [Google Scholar] [CrossRef]
- Takao, K.; Takamiya, K.; Ding, D.; Haraguchi, T.; Hiraoka, Y.; Nishimori, H.; Awazu, A. Torsional Turning Motion of Chromosomes as an Accelerating Force to Align Homologous Chromosomes during Meiosis. J. Phys. Soc. Jpn. 2019, 88, 023801. [Google Scholar] [CrossRef]
- Chacón, M.R.; Delivani, P.; Tolić, I.M. Meiotic Nuclear Oscillations Are Necessary to Avoid Excessive Chromosome Associations. Cell Rep. 2016, 17, 1632–1645. [Google Scholar] [CrossRef] [Green Version]
- Hiraoka, Y.; Dernburg, A.F. The SUN rises on meiotic chromosome dynamics. Dev. Cell 2009, 17, 598–605. [Google Scholar] [CrossRef] [Green Version]
- Koszul, R.; Kleckner, N. Dynamic chromosome movements during meiosis: A way to eliminate unwanted connections? Trends Cell Biol. 2009, 19, 716–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.Q.; Haraguchi, T.; Hiraoka, Y. Chromosomally-retained RNA mediates homologous pairing. Nucleus 2012, 3, 516–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.Q.; Yamamoto, A.; Haraguchi, T.; Hiraoka, Y. Dynamics of homologous chromosome pairing during meiotic prophase in fission yeast. Dev. Cell 2004, 6, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.Q.; Sakurai, N.; Katou, Y.; Itoh, T.; Shirahige, K.; Haraguchi, T.; Hiraoka, Y. Meiotic cohesins modulate chromosome compaction during meiotic prophase in fission yeast. J. Cell Biol. 2006, 174, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.Q.; Matsuda, A.; Okamasa, K.; Nagahama, Y.; Haraguchi, T.; Hiraoka, Y. Meiotic cohesin-based chromosome structure is essential for homologous chromosome pairing in Schizosaccharomyces pombe. Chromosoma 2016, 125, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covo, S.; Westmoreland, J.W.; Gordenin, D.A.; Resnick, M.A. Cohesin Is limiting for the suppression of DNA damage-induced recombination between homologous chromosomes. PLoS Genet. 2010, 6, e1001006. [Google Scholar] [CrossRef] [Green Version]
- Schwacha, A.; Kleckner, N. Interhomolog bias during meiotic recombination: Meiotic functions promote a highly differentiated interhomolog-only pathway. Cell 1997, 90, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.P.; Mirkin, E.V. So similar yet so different: The two ends of a double strand break. Mutat. Res. 2018, 809, 70–80. [Google Scholar] [CrossRef]
- Kim, K.P.; Weiner, B.M.; Zhang, L.; Jordan, A.; Dekker, J.; Kleckner, N. Sister cohesion and structural axis components mediate homolog bias of meiotic recombination. Cell 2010, 143, 924–937. [Google Scholar] [CrossRef] [Green Version]
- Lao, J.P.; Cloud, V.; Huang, C.C.; Grubb, J.; Thacker, D.; Lee, C.Y.; Dresser, M.E.; Hunter, N.; Bishop, D.K. Meiotic crossover control by concerted action of Rad51-Dmc1 in homolog template bias and robust homeostatic regulation. PLoS Genet. 2013, 9, e1003978. [Google Scholar] [CrossRef] [Green Version]
- Cromie, G.A.; Hyppa, R.W.; Taylor, A.F.; Zakharyevich, K.; Hunter, N.; Smith, G.R. Single Holliday junctions are intermediates of meiotic recombination. Cell 2006, 127, 1167–1178. [Google Scholar] [CrossRef] [Green Version]
- Sakuno, T.; Tanaka, K.; Hauf, S.; Watanabe, Y. Repositioning of Aurora B promoted by chiasmata ensures sister chromatid mono-orientation in meiosis I. Dev. Cell 2011, 21, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Suzuki, R.; Ohba, T.; Hinohara, Y.; Matsuhara, H.; Yoshida, M.; Itabashi, Y.; Murakami, H.; Yamamoto, A. Chiasmata promote monopolar attachment of sister chromatids and their co-segregation toward the proper pole during meiosis I. PLoS Genet. 2011, 7, e1001329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, A.; Hiraoka, Y. Monopolar spindle attachment of sister chromatids is ensured by two distinct mechanisms at the first meiotic division in fission yeast. EMBO J. 2003, 22, 2284–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokobayashi, S.; Yamamoto, M.; Watanabe, Y. Cohesins determine the attachment manner of kinetochores to spindle microtubules at meiosis I in fission yeast. Mol. Cell Biol. 2003, 23, 3965–3973. [Google Scholar] [CrossRef] [Green Version]
- Heidmann, D.; Horn, S.; Heidmann, S.; Schleiffer, A.; Nasmyth, K.; Lehner, C.F. The Drosophila meiotic kleisin C(2)M functions before the meiotic divisions. Chromosoma 2004, 113, 177–187. [Google Scholar] [CrossRef]
- Rabitsch, K.P.; Toth, A.; Galova, M.; Schleiffer, A.; Schaffner, G.; Aigner, E.; Rupp, C.; Penkner, A.M.; Moreno-Borchart, A.C.; Primig, M.; et al. A screen for genes required for meiosis and spore formation based on whole-genome expression. Curr. Biol. 2001, 11, 1001–1009. [Google Scholar] [CrossRef] [Green Version]
- Rabitsch, K.P.; Petronczki, M.; Javerzat, J.P.; Genier, S.; Chwalla, B.; Schleiffer, A.; Tanaka, T.U.; Nasmyth, K. Kinetochore recruitment of two nucleolar proteins is required for homolog segregation in meiosis I. Dev. Cell 2003, 4, 535–548. [Google Scholar] [CrossRef] [Green Version]
- Tóth, A.; Rabitsch, K.P.; Gálová, M.; Schleiffer, A.; Buonomo, S.B.; Nasmyth, K. Functional genomics identifies monopolin: A kinetochore protein required for segregation of homologs during meiosis i. Cell 2000, 103, 1155–1168. [Google Scholar] [CrossRef] [Green Version]
- Sakuno, T.; Tada, K.; Watanabe, Y. Kinetochore geometry defined by cohesion within the centromere. Nature 2009, 458, 852–858. [Google Scholar] [CrossRef]
- Ogushi, S.; Rattani, A.; Godwin, J.; Metson, J.; Schermelleh, L.; Nasmyth, K. Loss of sister kinetochore co-orientation and peri-centromeric cohesin protection after meiosis I depends on cleavage of centromeric REC8. Dev. Cell 2021, 56, 3100–3114.e3104. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ishiguro, K.; Nambu, A.; Akiyoshi, B.; Yokobayashi, S.; Kagami, A.; Ishiguro, T.; Pendas, A.M.; Takeda, N.; Sakakibara, Y.; et al. Meikin is a conserved regulator of meiosis-I-specific kinetochore function. Nature 2015, 517, 466–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, S.; Kim, J.; Yamagishi, Y.; Ishiguro, T.; Okada, Y.; Tanno, Y.; Sakuno, T.; Watanabe, Y. Meikin-associated polo-like kinase specifies Bub1 distribution in meiosis I. Genes Cells 2017, 22, 552–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galander, S.; Barton, R.E.; Borek, W.E.; Spanos, C.; Kelly, D.A.; Robertson, D.; Rappsilber, J.; Marston, A.L. Reductional Meiosis I Chromosome Segregation Is Established by Coordination of Key Meiotic Kinases. Dev. Cell 2019, 49, 526–541.e525. [Google Scholar] [CrossRef] [PubMed]
- Katis, V.L.; Matos, J.; Mori, S.; Shirahige, K.; Zachariae, W.; Nasmyth, K. Spo13 facilitates monopolin recruitment to kinetochores and regulates maintenance of centromeric cohesion during yeast meiosis. Curr. Biol. 2004, 14, 2183–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.H.; Kiburz, B.M.; Amon, A. Spo13 maintains centromeric cohesion and kinetochore coorientation during meiosis I. Curr. Biol. 2004, 14, 2168–2182. [Google Scholar] [CrossRef] [Green Version]
- Mehta, G.; Anbalagan, G.K.; Bharati, A.P.; Gadre, P.; Ghosh, S.K. An interplay between Shugoshin and Spo13 for centromeric cohesin protection and sister kinetochore mono-orientation during meiosis I in Saccharomyces cerevisiae. Curr. Genet. 2018, 64, 1141–1152. [Google Scholar] [CrossRef]
- Petronczki, M.; Matos, J.; Mori, S.; Gregan, J.; Bogdanova, A.; Schwickart, M.; Mechtler, K.; Shirahige, K.; Zachariae, W.; Nasmyth, K. Monopolar attachment of sister kinetochores at meiosis I requires casein kinase 1. Cell 2006, 126, 1049–1064. [Google Scholar] [CrossRef] [Green Version]
- Moore, D.P.; Orr-Weaver, T.L. Chromosome segregation during meiosis: Building an unambivalent bivalent. Curr. Top. Dev. Biol. 1998, 37, 263–299. [Google Scholar]
- Watanabe, Y.; Nurse, P. Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature 1999, 400, 461–464. [Google Scholar] [CrossRef]
- Chen, S.Y.; Tsubouchi, T.; Rockmill, B.; Sandler, J.S.; Richards, D.R.; Vader, G.; Hochwagen, A.; Roeder, G.S.; Fung, J.C. Global analysis of the meiotic crossover landscape. Dev. Cell 2008, 15, 401–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancera, E.; Bourgon, R.; Brozzi, A.; Huber, W.; Steinmetz, L.M. High-resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature 2008, 454, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beadle, G.W. A Possible Influence of the Spindle Fibre on Crossing-Over in Drosophila. Proc. Natl. Acad. Sci. USA 1932, 18, 160–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rockmill, B.; Voelkel-Meiman, K.; Roeder, G.S. Centromere-proximal crossovers are associated with precocious separation of sister chromatids during meiosis in Saccharomyces cerevisiae. Genetics 2006, 174, 1745–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassold, T.; Hunt, P. To err (meiotically) is human: The genesis of human aneuploidy. Nat. Rev. Genet. 2001, 2, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Kitajima, T.S.; Kawashima, S.A.; Watanabe, Y. The conserved kinetochore protein shugoshin protects centromeric cohesion during meiosis. Nature 2004, 427, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Indjeian, V.B.; Stern, B.M.; Murray, A.W. The centromeric protein Sgo1 is required to sense lack of tension on mitotic chromosomes. Science 2005, 307, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Katis, V.L.; Galova, M.; Rabitsch, K.P.; Gregan, J.; Nasmyth, K. Maintenance of cohesin at centromeres after meiosis I in budding yeast requires a kinetochore-associated protein related to MEI-S332. Curr. Biol. 2004, 14, 560–572. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, T.S.; Sakuno, T.; Ishiguro, K.; Iemura, S.; Natsume, T.; Kawashima, S.A.; Watanabe, Y. Shugoshin collaborates with protein phosphatase 2A to protect cohesin. Nature 2006, 441, 46–52. [Google Scholar] [CrossRef]
- Riedel, C.G.; Katis, V.L.; Katou, Y.; Mori, S.; Itoh, T.; Helmhart, W.; Gálová, M.; Petronczki, M.; Gregan, J.; Cetin, B.; et al. Protein phosphatase 2A protects centromeric sister chromatid cohesion during meiosis I. Nature 2006, 441, 53–61. [Google Scholar] [CrossRef]
- Ishiguro, T.; Tanaka, K.; Sakuno, T.; Watanabe, Y. Shugoshin-PP2A counteracts casein-kinase-1-dependent cleavage of Rec8 by separase. Nat. Cell Biol. 2010, 12, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kitajima, T.S.; Tanno, Y.; Yoshida, K.; Morita, T.; Miyano, T.; Miyake, M.; Watanabe, Y. Unified mode of centromeric protection by shugoshin in mammalian oocytes and somatic cells. Nat. Cell Biol. 2008, 10, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Ai, J.S.; Shi, L.H.; Wei, L.; Yuan, J.; Ouyang, Y.C.; Hou, Y.; Chen, D.Y.; Schatten, H.; Sun, Q.Y. Shugoshin1 may play important roles in separation of homologous chromosomes and sister chromatids during mouse oocyte meiosis. PLoS ONE 2008, 3, e3516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Huang, L.; Markowitz, T.E.; Blitzblau, H.G.; Chen, D.; Klein, F.; Hochwagen, A. Transcription dynamically patterns the meiotic chromosome-axis interface. Elife 2015, 4, e07424. [Google Scholar] [CrossRef] [PubMed]
- Panizza, S.; Mendoza, M.A.; Berlinger, M.; Huang, L.; Nicolas, A.; Shirahige, K.; Klein, F. Spo11-accessory proteins link double-strand break sites to the chromosome axis in early meiotic recombination. Cell 2011, 146, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuno, T.; Watanabe, Y. Phosphorylation of cohesin Rec11/SA3 by casein kinase 1 promotes homologous recombination by assembling the meiotic chromosome axis. Dev. Cell 2015, 32, 220–230. [Google Scholar] [CrossRef] [Green Version]
- Phadnis, N.; Cipak, L.; Polakova, S.; Hyppa, R.W.; Cipakova, I.; Anrather, D.; Karvaiova, L.; Mechtler, K.; Smith, G.R.; Gregan, J. Casein Kinase 1 and Phosphorylation of Cohesin Subunit Rec11 (SA3) Promote Meiotic Recombination through Linear Element Formation. PLoS Genet. 2015, 11, e1005225. [Google Scholar] [CrossRef] [Green Version]
- Ponticelli, A.S.; Smith, G.R. Meiotic recombination-deficient mutants of Schizosaccharomyces pombe. Genetics 1989, 123, 45–54. [Google Scholar] [CrossRef]
- Miyoshi, T.; Ito, M.; Kugou, K.; Yamada, S.; Furuichi, M.; Oda, A.; Yamada, T.; Hirota, K.; Masai, H.; Ohta, K. A central coupler for recombination initiation linking chromosome architecture to S phase checkpoint. Mol. Cell 2012, 47, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Biswas, U.; Hempel, K.; Llano, E.; Pendas, A.; Jessberger, R. Distinct Roles of Meiosis-Specific Cohesin Complexes in Mammalian Spermatogenesis. PLoS Genet. 2016, 12, e1006389. [Google Scholar] [CrossRef]
- Ward, A.; Hopkins, J.; Mckay, M.; Murray, S.; Jordan, P.W. Genetic Interactions Between the Meiosis-Specific Cohesin Components, STAG3, REC8, and RAD21L. G3 Genes Genomes Genet. 2016, 6, 1713–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Pratto, F.; Schimenti, J.C.; Turner, J.M.; Camerini-Otero, R.D.; Hoog, C. Phosphorylation of chromosome core components may serve as axis marks for the status of chromosomal events during mammalian meiosis. PLoS Genet. 2012, 8, e1002485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasierbek, P.; Fodermayr, M.; Jantsch, V.; Jantsch, M.; Schweizer, D.; Loidl, J. The Caenorhabditis elegans SCC-3 homologue is required for meiotic synapsis and for proper chromosome disjunction in mitosis and meiosis. Exp. Cell Res. 2003, 289, 245–255. [Google Scholar] [CrossRef]
- Fukuda, T.; Daniel, K.; Wojtasz, L.; Toth, A.; Höög, C. A novel mammalian HORMA domain-containing protein, HORMAD1, preferentially associates with unsynapsed meiotic chromosomes. Exp. Cell Res. 2010, 316, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Pelttari, J.; Hoja, M.R.; Yuan, L.; Liu, J.G.; Brundell, E.; Moens, P.; Santucci-Darmanin, S.; Jessberger, R.; Barbero, J.L.; Heyting, C.; et al. A meiotic chromosomal core consisting of cohesin complex proteins recruits DNA recombination proteins and promotes synapsis in the absence of an axial element in mammalian meiotic cells. Mol. Cell Biol. 2001, 21, 5667–5677. [Google Scholar] [CrossRef] [Green Version]
- Zickler, D.; Kleckner, N. Meiotic chromosomes: Integrating structure and function. Annu. Rev. Genet. 1999, 33, 603–754. [Google Scholar] [CrossRef]
- Patel, L.; Kang, R.; Rosenberg, S.C.; Qiu, Y.; Raviram, R.; Chee, S.; Hu, R.; Ren, B.; Cole, F.; Corbett, K.D. Dynamic reorganization of the genome shapes the recombination landscape in meiotic prophase. Nat. Struct. Mol. Biol. 2019, 26, 164–174. [Google Scholar] [CrossRef]
- Silva, M.C.C.; Powell, S.; Ladstätter, S.; Gassler, J.; Stocsits, R.; Tedeschi, A.; Peters, J.M.; Tachibana, K. Wapl releases Scc1-cohesin and regulates chromosome structure and segregation in mouse oocytes. J. Cell Biol. 2020, 219, e201906100. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, H.; Zhang, Y.; Du, Z.; Si, W.; Fan, S.; Qin, D.; Wang, M.; Duan, Y.; Li, L.; et al. Reprogramming of Meiotic Chromatin Architecture during Spermatogenesis. Mol. Cell 2019, 73, 547–561.e546. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Wang, X.; Jiang, H.; Wang, R.; Chen, J.; Chen, Y.; Xu, Q.; Cao, J.; Gong, X.; Wu, J.; et al. Reorganized 3D Genome Structures Support Transcriptional Regulation in Mouse Spermatogenesis. iScience 2020, 23, 101034. [Google Scholar] [CrossRef]
- Alavattam, K.G.; Maezawa, S.; Sakashita, A.; Khoury, H.; Barski, A.; Kaplan, N.; Namekawa, S.H. Attenuated chromatin compartmentalization in meiosis and its maturation in sperm development. Nat. Struct. Mol. Biol. 2019, 26, 175–184. [Google Scholar] [CrossRef]
- Zuo, W.; Chen, G.; Gao, Z.; Li, S.; Chen, Y.; Huang, C.; Chen, J.; Chen, Z.; Lei, M.; Bian, Q. Stage-resolved Hi-C analyses reveal meiotic chromosome organizational features influencing homolog alignment. Nat. Commun. 2021, 12, 5827. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zheng, H.; Kawamura, Y.K.; Zhang, K.; Gassler, J.; Powell, S.; Xu, Q.; Lin, Z.; Xu, K.; Zhou, Q.; et al. Polycomb Group Proteins Regulate Chromatin Architecture in Mouse Oocytes and Early Embryos. Mol. Cell 2020, 77, 825–839. [Google Scholar] [CrossRef] [PubMed]
- Vara, C.; Paytuví-Gallart, A.; Cuartero, Y.; Le Dily, F.; Garcia, F.; Salvà-Castro, J.; Gómez-H, L.; Julià, E.; Moutinho, C.; Aiese Cigliano, R.; et al. Three-Dimensional Genomic Structure and Cohesin Occupancy Correlate with Transcriptional Activity during Spermatogenesis. Cell Rep. 2019, 28, 352–367.e359. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Hu, T.; Jiang, H.; Wang, R.; Xu, Q.; Zhang, S.; Cao, J.; Song, X. Rearrangement of macronucleus chromosomes correspond to TAD-like structures of micronucleus chromosomes in. Genome Res. 2020, 30, 406–414. [Google Scholar] [CrossRef]
- Costantino, L.; Hsieh, T.S.; Lamothe, R.; Darzacq, X.; Koshland, D. Cohesin residency determines chromatin loop patterns. Elife 2020, 9, e59889. [Google Scholar] [CrossRef]
- Del Mazo, J.; Gil-Alberdi, L. Multistranded organization of the lateral elements of the synaptonemal complex in the rat and mouse. Cytogenet. Genome Res. 1986, 41, 219–224. [Google Scholar] [CrossRef]
- Kierszenbaum, A.L.; Tres, L.L. Transcription sites in spread meiotic prophase chromosomes from mouse spermatocytes. J. Cell Biol. 1974, 63, 923–935. [Google Scholar] [CrossRef]
- Kim, Y.; Shi, Z.; Zhang, H.; Finkelstein, I.J.; Yu, H. Human cohesin compacts DNA by loop extrusion. Science 2019, 366, 1345–1349. [Google Scholar] [CrossRef]
- Davidson, I.F.; Peters, J.M. Genome folding through loop extrusion by SMC complexes. Nat. Rev. Mol. Cell Biol. 2021, 22, 445–464. [Google Scholar] [CrossRef]
- Mizuguchi, T.; Fudenberg, G.; Mehta, S.; Belton, J.M.; Taneja, N.; Folco, H.D.; FitzGerald, P.; Dekker, J.; Mirny, L.; Barrowman, J.; et al. Cohesin-dependent globules and heterochromatin shape 3D genome architecture in S. pombe. Nature 2014, 516, 432–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fudenberg, G.; Imakaev, M.; Lu, C.; Goloborodko, A.; Abdennur, N.; Mirny, L.A. Formation of Chromosomal Domains by Loop Extrusion. Cell Rep. 2016, 15, 2038–2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.S.P.; Huang, S.C.; Glenn St Hilaire, B.; Engreitz, J.M.; Perez, E.M.; Kieffer-Kwon, K.R.; Sanborn, A.L.; Johnstone, S.E.; Bascom, G.D.; Bochkov, I.D.; et al. Cohesin Loss Eliminates All Loop Domains. Cell 2017, 171, 305–320.e324. [Google Scholar] [CrossRef] [Green Version]
- Hartman, T.; Stead, K.; Koshland, D.; Guacci, V. Pds5p is an essential chromosomal protein required for both sister chromatid cohesion and condensation in Saccharomyces cerevisiae. J. Cell Biol. 2000, 151, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Panizza, S.; Tanaka, T.; Hochwagen, A.; Eisenhaber, F.; Nasmyth, K. Pds5 cooperates with cohesin in maintaining sister chromatid cohesion. Curr. Biol. 2000, 10, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Stead, K.; Aguilar, C.; Hartman, T.; Drexel, M.; Meluh, P.; Guacci, V. Pds5p regulates the maintenance of sister chromatid cohesion and is sumoylated to promote the dissolution of cohesion. J. Cell Biol. 2003, 163, 729–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Hao, Z.; Kai, M.; Okayama, H. Establishment and maintenance of sister chromatid cohesion in fission yeast by a unique mechanism. EMBO J. 2001, 20, 5779–5790. [Google Scholar] [CrossRef] [Green Version]
- Gandhi, R.; Gillespie, P.J.; Hirano, T. Human Wapl is a cohesin-binding protein that promotes sister-chromatid resolution in mitotic prophase. Curr. Biol. 2006, 16, 2406–2417. [Google Scholar] [CrossRef] [Green Version]
- Kueng, S.; Hegemann, B.; Peters, B.H.; Lipp, J.J.; Schleiffer, A.; Mechtler, K.; Peters, J.M. Wapl controls the dynamic association of cohesin with chromatin. Cell 2006, 127, 955–967. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, T.; Ladurner, R.; Schmitz, J.; Kreidl, E.; Schleiffer, A.; Bhaskara, V.; Bando, M.; Shirahige, K.; Hyman, A.A.; Mechtler, K.; et al. Sororin mediates sister chromatid cohesion by antagonizing Wapl. Cell 2010, 143, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Challa, K.; Lee, M.S.; Shinohara, M.; Kim, K.P.; Shinohara, A. Rad61/Wpl1 (Wapl), a cohesin regulator, controls chromosome compaction during meiosis. Nucleic Acids Res. 2016, 44, 3190–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challa, K.; Shinohara, M.; Shinohara, A. Meiotic prophase-like pathway for cleavage-independent removal of cohesin for chromosome morphogenesis. Curr. Genet. 2019, 65, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Haarhuis, J.H.I.; van der Weide, R.H.; Blomen, V.A.; Yáñez-Cuna, J.O.; Amendola, M.; van Ruiten, M.S.; Krijger, P.H.L.; Teunissen, H.; Medema, R.H.; van Steensel, B.; et al. The Cohesin Release Factor WAPL Restricts Chromatin Loop Extension. Cell 2017, 169, 693–707.e614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Joo, J.H.; Yun, H.; Kleckner, N.; Kim, K.P. Recruitment of Rec8, Pds5 and Rad61/Wapl to meiotic homolog pairing, recombination, axis formation and S-phase. Nucleic Acids Res. 2019, 47, 11691–11708. [Google Scholar] [CrossRef] [Green Version]
- Tedeschi, A.; Wutz, G.; Huet, S.; Jaritz, M.; Wuensche, A.; Schirghuber, E.; Davidson, I.F.; Tang, W.; Cisneros, D.A.; Bhaskara, V.; et al. Wapl is an essential regulator of chromatin structure and chromosome segregation. Nature 2013, 501, 564–568. [Google Scholar] [CrossRef]
- Viera, A.; Berenguer, I.; Ruiz-Torres, M.; Gómez, R.; Guajardo, A.; Barbero, J.L.; Losada, A.; Suja, J.A. PDS5 proteins regulate the length of axial elements and telomere integrity during male mouse meiosis. EMBO Rep. 2020, 21, e49273. [Google Scholar] [CrossRef]
- Jin, H.; Guacci, V.; Yu, H.G. Pds5 is required for homologue pairing and inhibits synapsis of sister chromatids during yeast meiosis. J. Cell Biol. 2009, 186, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Crawley, O.; Barroso, C.; Testori, S.; Ferrandiz, N.; Silva, N.; Castellano-Pozo, M.; Jaso-Tamame, A.L.; Martinez-Perez, E. Cohesin-interacting protein WAPL-1 regulates meiotic chromosome structure and cohesion by antagonizing specific cohesin complexes. Elife 2016, 5, e10851. [Google Scholar] [CrossRef]
- Wutz, G.; Várnai, C.; Nagasaka, K.; Cisneros, D.A.; Stocsits, R.R.; Tang, W.; Schoenfelder, S.; Jessberger, G.; Muhar, M.; Hossain, M.J.; et al. Topologically associating domains and chromatin loops depend on cohesin and are regulated by CTCF, WAPL, and PDS5 proteins. EMBO J. 2017, 36, 3573–3599. [Google Scholar] [CrossRef]
- Li, Y.; Haarhuis, J.H.I.; Sedeño Cacciatore, Á.; Oldenkamp, R.; van Ruiten, M.S.; Willems, L.; Teunissen, H.; Muir, K.W.; de Wit, E.; Rowland, B.D.; et al. The structural basis for cohesin-CTCF-anchored loops. Nature 2020, 578, 472–476. [Google Scholar] [CrossRef]
- Nora, E.P.; Goloborodko, A.; Valton, A.L.; Gibcus, J.H.; Uebersohn, A.; Abdennur, N.; Dekker, J.; Mirny, L.A.; Bruneau, B.G. Targeted Degradation of CTCF Decouples Local Insulation of Chromosome Domains from Genomic Compartmentalization. Cell 2017, 169, 930–944.e922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, I.F.; Bauer, B.; Goetz, D.; Tang, W.; Wutz, G.; Peters, J.M. DNA loop extrusion by human cohesin. Science 2019, 366, 1338–1345. [Google Scholar] [CrossRef] [PubMed]
- Ganji, M.; Shaltiel, I.A.; Bisht, S.; Kim, E.; Kalichava, A.; Haering, C.H.; Dekker, C. Real-time imaging of DNA loop extrusion by condensin. Science 2018, 360, 102–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Kerssemakers, J.; Shaltiel, I.A.; Haering, C.H.; Dekker, C. DNA-loop extruding condensin complexes can traverse one another. Nature 2020, 579, 438–442. [Google Scholar] [CrossRef]
- Kschonsak, M.; Merkel, F.; Bisht, S.; Metz, J.; Rybin, V.; Hassler, M.; Haering, C.H. Structural Basis for a Safety-Belt Mechanism That Anchors Condensin to Chromosomes. Cell 2017, 171, 588–600.e524. [Google Scholar] [CrossRef] [Green Version]
- Terakawa, T.; Bisht, S.; Eeftens, J.M.; Dekker, C.; Haering, C.H.; Greene, E.C. The condensin complex is a mechanochemical motor that translocates along DNA. Science 2017, 358, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Hore, T.A.; Deakin, J.E.; Marshall Graves, J.A. The evolution of epigenetic regulators CTCF and BORIS/CTCFL in amniotes. PLoS Genet. 2008, 4, e1000169. [Google Scholar] [CrossRef] [Green Version]
- Barbero, J.L. Genetic basis of cohesinopathies. Appl. Clin. Genet. 2013, 6, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, Z.; Bando, M.; Itoh, T.; Deardorff, M.A.; Clark, D.; Kaur, M.; Tandy, S.; Kondoh, T.; Rappaport, E.; et al. Transcriptional dysregulation in NIPBL and cohesin mutant human cells. PLoS Biol. 2009, 7, e1000119. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakuno, T.; Hiraoka, Y. Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes. Genes 2022, 13, 200. https://doi.org/10.3390/genes13020200
Sakuno T, Hiraoka Y. Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes. Genes. 2022; 13(2):200. https://doi.org/10.3390/genes13020200
Chicago/Turabian StyleSakuno, Takeshi, and Yasushi Hiraoka. 2022. "Rec8 Cohesin: A Structural Platform for Shaping the Meiotic Chromosomes" Genes 13, no. 2: 200. https://doi.org/10.3390/genes13020200