
Oligo-FISH Can Identify Chromosomes and Distinguish Hippophaë rhamnoides L. Taxa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oligo-Probe Preparation
2.2. FISH and Karyotype Analysis
3. Results
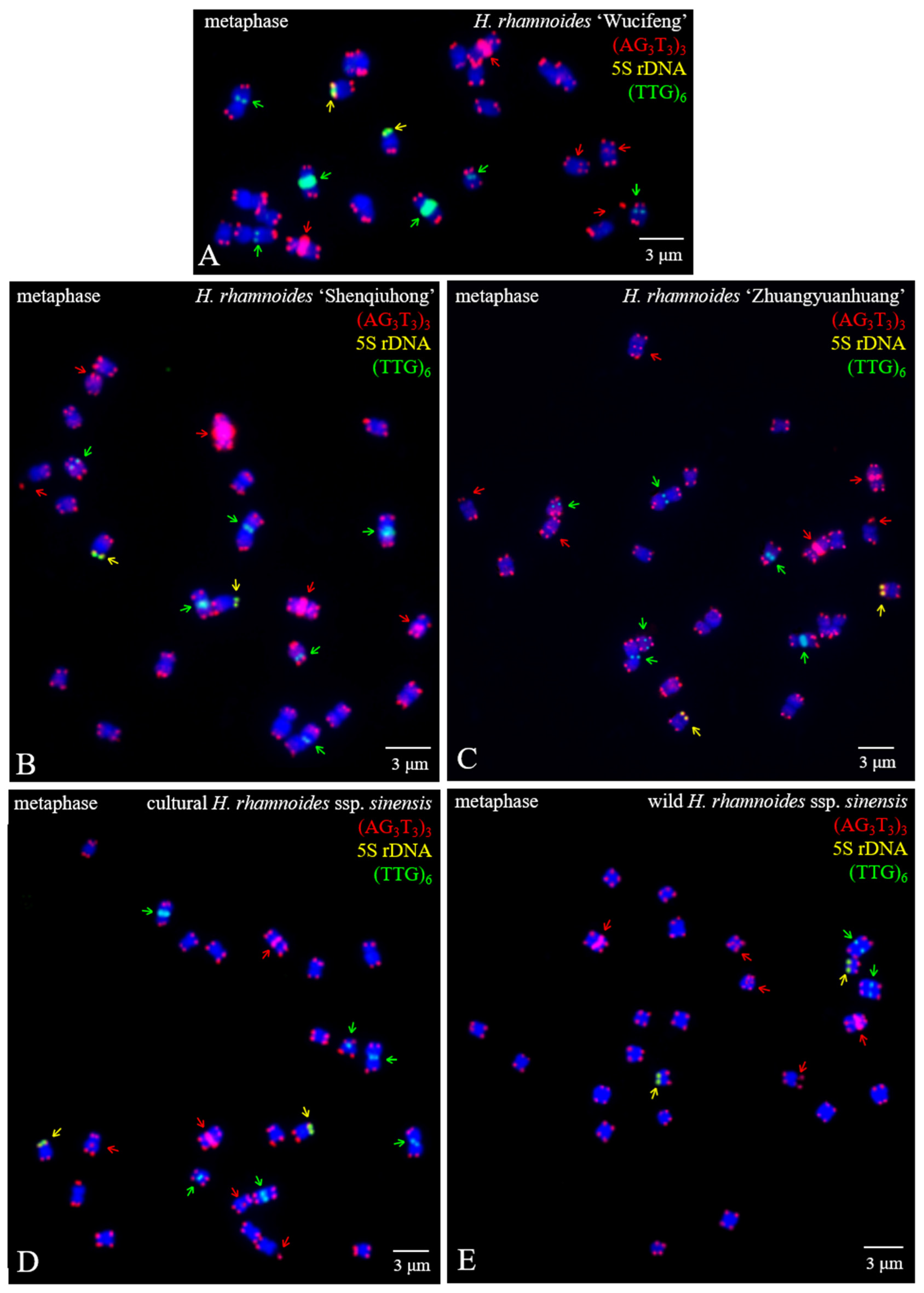
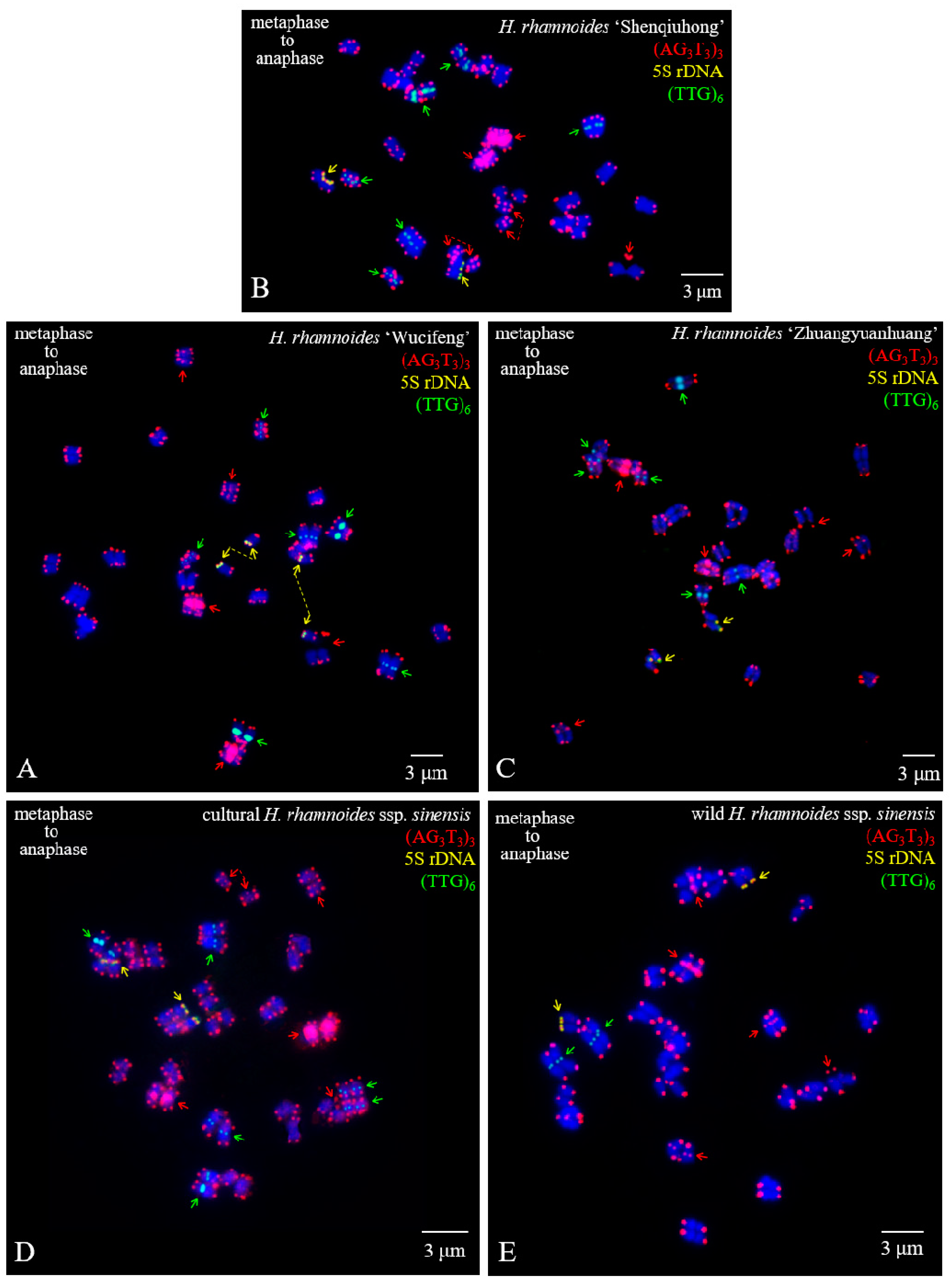
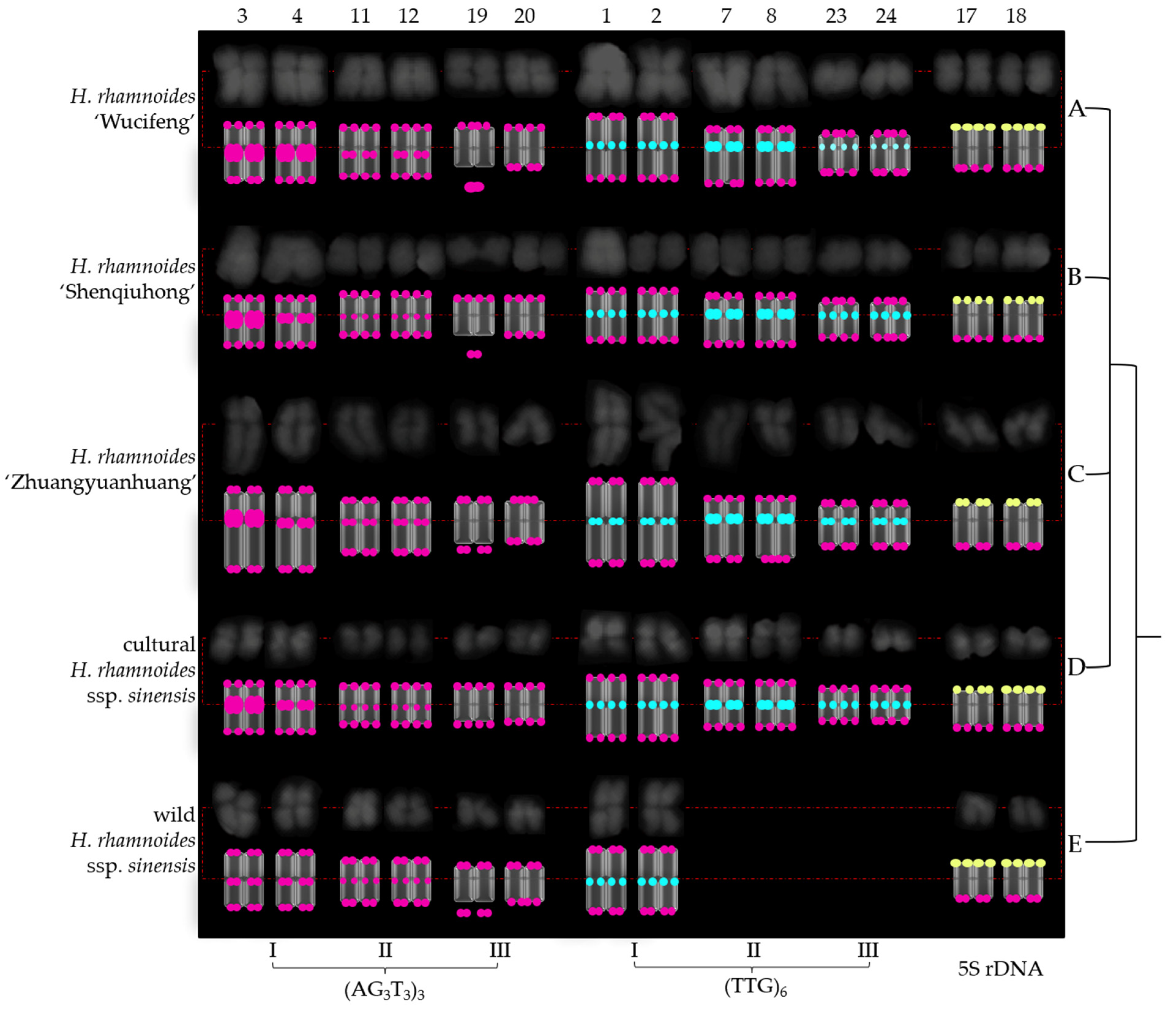
3.1. FISH-Enabled Visualization of H. rhamnoides Chromosomes
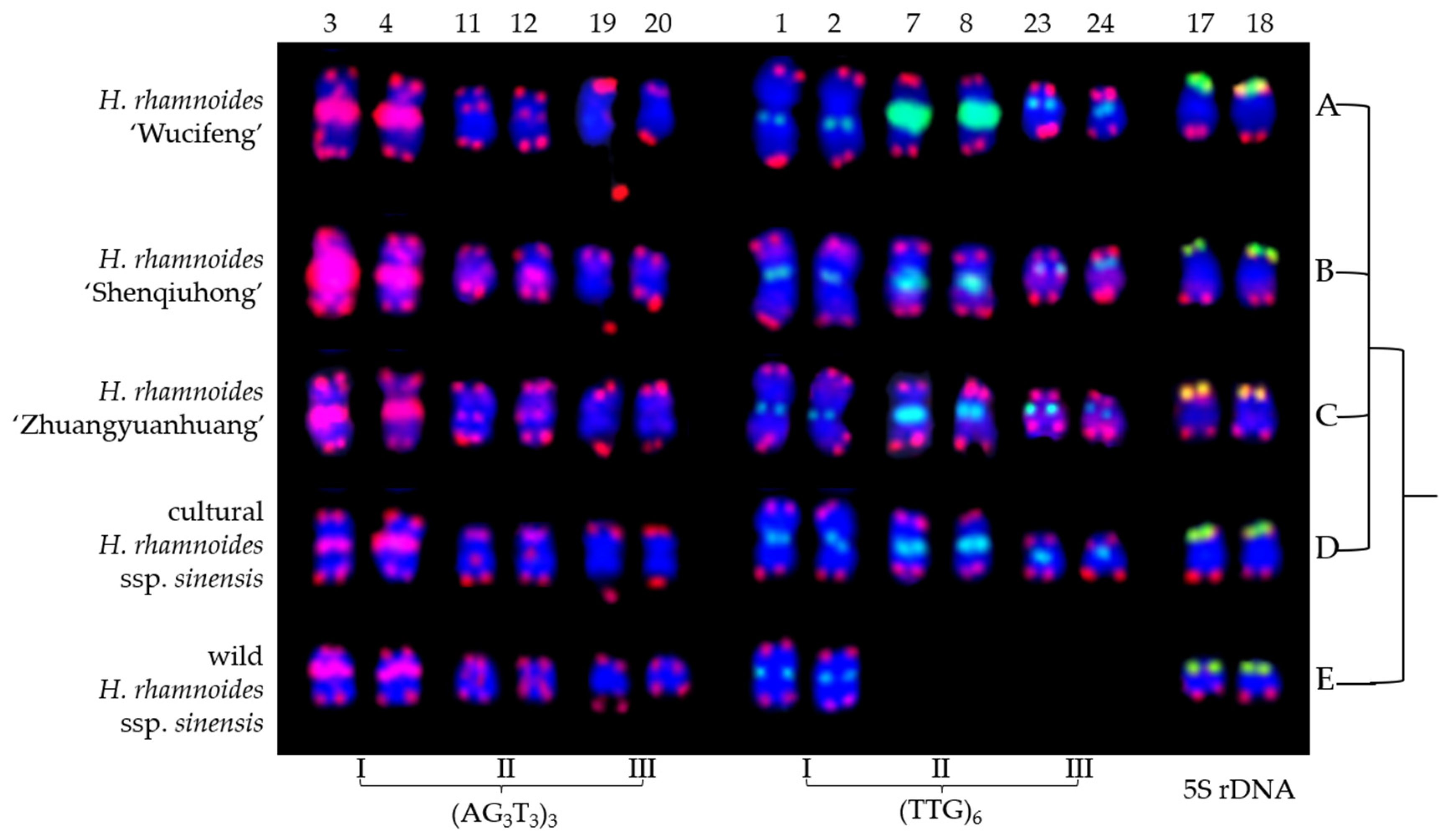
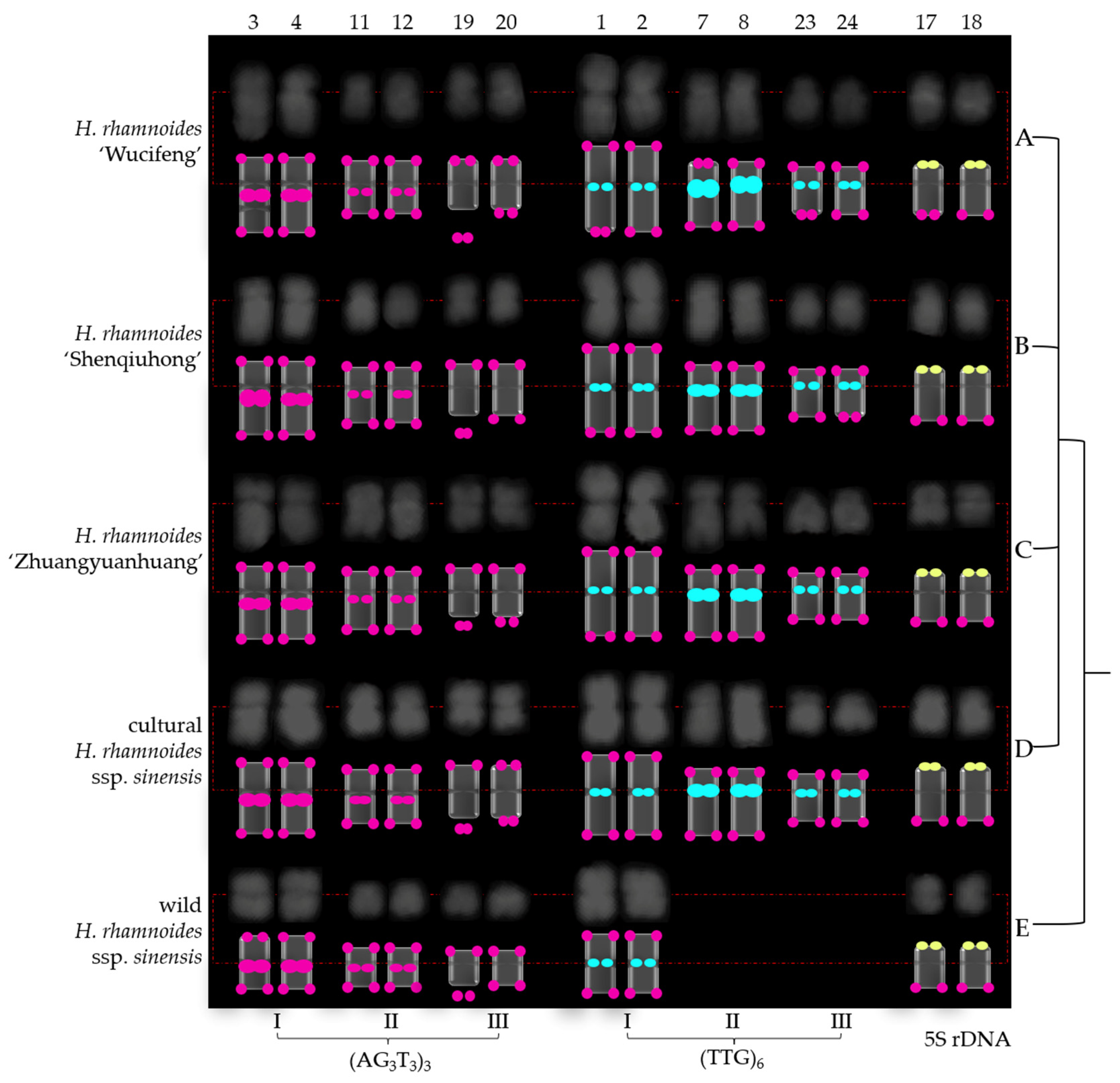
3.2. Physical Map Distinguished Chromosomes
4. Discussion
4.1. Karyotype Analysis
4.2. Role of (AG3T3)3, (TTG)6, and 5S rDNA
4.3. Detection of the X/Y-Chromosome in H. rhamnoides
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, B.; Arora, S.; Sahoo, D.; Deswal, R. Comparative fatty acid profiling of Indian sea buckthorn showed altitudinal gradient dependent species-specific variations. Physiol. Mol. Biol. Plants 2020, 26, 41–49. [Google Scholar] [CrossRef]
- Jia, D.R.; Abbott, R.J.; Liu, T.L.; Mao, K.S.; Bartish, I.V.; Liu, J.Q. Out of the Qinghai-Tibet Plateau: Evidence for the origin and dispersal of Eurasian temperate plants from a phylogeographic study of Hippophaë rhamnoides (Elaeagnaceae). New Phytol. 2012, 194, 1123–1133. [Google Scholar] [CrossRef]
- Bonciu, E.; Olaru, A.L.; Rosculete, E.; Rosculete, C.A. Cytogenetic study on the biostimulation potential of the aqueous fruit extract of Hippophaë rhamnoides for a sustainable agricultural ecosystem. Plants 2020, 9, 843. [Google Scholar] [CrossRef]
- Lee, Y.H.; Jang, H.J.; Park, K.H.; Kim, S.H.; Kim, J.K.; Kim, J.C.; Jang, T.S.; Kim, K.H. Phytochemical analysis of the fruits of sea buckthorn (Hippophaë rhamnoides): Identification of organic acid derivatives. Plants 2021, 10, 860. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Xu, Z.; Liao, X. Bioactive compounds, health benefits and functional food products of sea buckthorn: A review. Crit. Rev. Food Sci. Nutr. 2021, 3, 2–12. [Google Scholar] [CrossRef]
- Gâtlan, A.M.; Gutt, G. Sea Buckthorn in plant based diets. An analytical approach of sea buckthorn fruits composition: Nutritional value, applications, and health Benefits. Int. J. Environ. Res. Public. Health 2021, 18, 8986. [Google Scholar] [CrossRef]
- Avdeyev, V.I. Novaya taxonomiya roda oblepikha: Hippophaë L. Izv. Akad. Nauk. Tadzhikskoy SSR Otd. Biol. Nauk. 1983, 93, 11–17. [Google Scholar]
- Servettaz, C. Monographie der Elaeagnaceae. Beih. Zum Bot. Cent. 1908, 25, 18. [Google Scholar]
- Lian, T.S.; Chen, X.L. Taxonomy of Genus Hippophaë L. Hippophaë 1996, 9, 15–24. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=2121204 (accessed on 21 December 2021).
- Rousi, A. The genus Hippophaë L. A taxonomic study. Ann. Bot. Fenn. 1971, 8, 177–227. [Google Scholar]
- Hyvönen, J. On phylogeny of Hippophaë (Eleagnaceae). Nordic. J. Bot. 1996, 16, 51–62. [Google Scholar] [CrossRef]
- Bartish, I.V.; Jeppsson, N.; Nybom, H.; Swenson, U. Phylogeny of Hippophaë (Elaeagnaceae) inferred from parsimony analysis of chloroplast DNA and morphology. Syst. Bot. 2002, 27, 41–54. [Google Scholar] [CrossRef]
- WFO. World Flora Online. Published on the Internet. 2021. Available online: http://www.worldfloraonline.org (accessed on 24 October 2021).
- Li, H.; Ruan, C.; Ding, J.; Li, J.; Wang, L.; Tian, X. Diversity in sea buckthorn (Hippophaë rhamnoides L.) accessions with different origins based on morphological characteristics, oil traits, and microsatellite markers. PLoS ONE 2020, 15, e0230356. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, W.; Liu, C.; Zhang, Y.; Chen, Y.; Song, M.; Fan, G.; Liu, X.; Xiang, L.; Zhang, Y. Identification of Hippophaë species (Shaji) through DNA barcodes. Chin. Med. 2015, 10, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Ganie, S.H.; Mangla, Y.; Dar, T.U.; Chaudhary, M.; Thakur, R.K.; Tandon, R.; Raina, S.N.; Goel, S. ISSR markers for gender identification and genetic diagnosis of Hippophaë rhamnoides ssp. turkestanica growing at high altitudes in Ladakh region (Jammu and Kashmir). Protoplasma 2017, 254, 1063–1077. [Google Scholar] [CrossRef]
- Puterova, J.; Razumova, O.; Martinek, T.; Alexandrov, O.; Divashuk, M.; Kubat, Z.; Hobza, R.; Karlov, G.; Kejnovsky, E. Satellite DNA and transposable elements in seabuckthorn (Hippophaë rhamnoides), a dioecious plant with small Y and large X chromosomes. Genome Biol. Evol. 2017, 9, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Jiang, C.; Yuan, W.; Zhang, M.; Fang, Z.; Li, Y.; Li, G.; Jia, J.; Yang, Z. A universal karyotypic system for hexaploid and diploid Avena species brings oat cytogenetics into the genomics era. BMC Plant Biol. 2021, 21, 213–227. [Google Scholar] [CrossRef]
- Fu, L.; Wang, Q.; Li, L.; Lang, T.; Guo, J.; Wang, S.; Sun, Z.; Han, S.; Huang, B.; Dong, W.; et al. Physical mapping of repetitive oligonucleotides facilitates the establishment of a genome map-based karyotype to identify chromosomal variations in peanut. BMC Plant Biol. 2021, 21, 107–118. [Google Scholar] [CrossRef]
- Meng, Z.; Han, J.; Lin, Y.; Zhao, Y.; Lin, Q.; Ma, X.; Wang, J.; Zhang, M.; Zhang, L.; Yang, Q.; et al. Characterization of a Saccharum spontaneum with a basic chromosome number of x = 10 provides new insights on genome evolution in genus Saccharum. Theor. Appl. Genet. 2020, 133, 187–199. [Google Scholar] [CrossRef]
- He, L.; Zhao, H.; He, J.; Yang, Z.; Guan, B.; Chen, K.; Hong, Q.; Wang, J.; Liu, J.; Jiang, J. Extraordinarily conserved chromosomal synteny of Citrus species revealed by chromosome-specific painting. Plant J. 2020, 103, 2225–2235. [Google Scholar] [CrossRef]
- Deng, H.; Tang, G.; Xu, N.; Gao, Z.; Lin, L.; Liang, D.; Xia, H.; Deng, Q.; Wang, J.; Cai, Z.; et al. Integrate and d karyotypes of diploid and tetraploid carrizo citrange (Citrus sinensis L. Osbeck x Poncirus trifoliata L. Raf.) as determined by sequential multicolor fluorescence in situ hybridization with tandemly repeated DNA sequences. Front. Plant Sci. 2020, 11, 569–578. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, T.; Wu, Y.; Zhang, W.; Zhang, P.; Xi, M.; Jiang, J. An extraordinarily stable karyotype of the woody Populus species revealed by chromosome painting. Plant J. 2020, 101, 253–264. [Google Scholar] [CrossRef]
- Cai, Q.; Zhang, D.; Liu, Z.L.; Wang, X.R. Chromosomal localization of 5S and 18S rDNA in five species of subgenus Strobus and their implications for genome evolution of Pinus. Ann. Bot. 2006, 97, 715–722. [Google Scholar] [CrossRef]
- Hizume, M.; Shibata, F.; Matsusaki, Y.; Garajova, Z. Chromosome identification and comparative karyotypic analyses of four Pinus species. Theor. Appl. Genet. 2002, 105, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.X.; Zeng, H.; Li, X.L.; Chen, C.B.; Song, W.Q.; Chen, R.Y. The molecular characterization of maize B chromosome specific AFLPs. Cell Res. 2002, 12, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Luo, X. First report of bicolour FISH of Berberis diaphana and B. soulieana reveals interspecific differences and co-localization of (AGGGTTT)3 and rDNA 5S in B. diaphana. Hereditas 2019, 156, 13–20. [Google Scholar] [CrossRef]
- Luo, X.; Liu, J. Fluorescence in situ hybridization (FISH) analysis of the locations of the oligonucleotides 5S rDNA, (AGGGTTT)3, and (TTG)6 in three genera of Oleaceae and their phylogenetic framework. Genes 2019, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Chen, J. Physical Map of FISH 5S rDNA and (AG3T3)3 Signals displays Chimonanthus campanulatus R.H. Chang & C.S. Ding chromosomes, reproduces its metaphase dynamics and distinguishes its chromosomes. Genes 2019, 10, 904. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Chen, J. Distinguishing Sichuan walnut cultivars and examining their relationships with Juglans regia and J. sigillata by FISH, early-fruiting gene analysis, and SSR analysis. Front. Plant Sci. 2020, 11, 27–41. [Google Scholar] [CrossRef]
- Luo, X.; He, Z. Distribution of FISH oligo-5S rDNA and oligo-(AGGGTTT)3 in Hibiscus mutabilis L. Genome 2021, 64, 655–664. [Google Scholar] [CrossRef]
- Luo, X.; Liu, J.; Zhao, A.J.; Chen, X.H.; Wan, W.L.; Chen, L. Karyotype analysis of Piptanthus concolor based on FISH with a oligonucleotides for rDNA 5S. Sci. Hortic. 2017, 226, 361–365. [Google Scholar] [CrossRef]
- Luo, X.; Liu, J.; Wang, J.; Gong, W.; Chen, L.; Wan, W. FISH analysis of Zanthoxylum armatum based on oligonucleotides for 5S rDNA and (GAA)6. Genome 2018, 61, 699–702. [Google Scholar] [CrossRef]
- Luo, X.; Tinker, N.A.; Zhou, Y.; Liu, J.; Wan, W.; Chen, L. Chromosomal distributions of oligo-Am1 and (TTG)6 trinucleotide and their utilization in genome association analysis of sixteen Avena species. Genet. Resour. Crop Evol. 2018, 65, 1625–1635. [Google Scholar] [CrossRef]
- Liu, H.Z.; Sheng, L.X. Studies on karyotype of three sea buckthorn. J. Jilin Agr. Univ. 2011, 33, 628–631, 636. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, B.S. Karyotype Analysis of Hippophaë rhamnoides ssp. sinensis. J. Wuhan Bot. Res. 1988, 6, 195–197. Available online: https://kns.cnki.net/KCMS/detail/detail.aspx?dbname=cjfd1988&filename=wzxy198802016 (accessed on 21 December 2021).
- Xing, M.S.; Xue, C.J.; Li, R.P. Karyotype analysis of sea buckthorn. J. Shanxi Univ. (Nat. Sci. Ed.). 1989, 12, 323–330. [Google Scholar] [CrossRef]
- Guerra, M. Cytogenetics of Rutaceae. V. high chromosomal variability in Citrus species revealed by CMA/DAPI staining. Heredity 1993, 71, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Liang, W.F.; Li, C.B.; Lian, Y.S. Karyotype Analysis of Four Sea Buckthorn in Qinghai. Hippophaë 1997, 10, 5–11. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=2814051 (accessed on 21 December 2021).
- Li, X.L.; Song, W.Q.; Chen, R.Y. Studies on Karyotype of Some Berry Plants in North China. J. Wuhan Bot. Res. 1983, 11, 289–293. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=1196583 (accessed on 21 December 2021).
- Rousi, A.; Arohonka, T. C-Band and Ploidy Level of Hippophaë rhamnoides. Hereditas 1980, 92, 327–330. Available online: https://www.cabdirect.org/cabdirect/abstract/19821606781 (accessed on 21 December 2021). [CrossRef]
- Shchapov, N.S. On the karyology of Hippophaë¨ rhamnoides L. Tsitol. Genet. 1979, 13, 45–47. [Google Scholar]
- Borodina, N.A. Polutshenie isskustvennykh poliploidov oblepikhi. Bjull. Glavn. Bot. Sada. 1976, 102, 62–67. [Google Scholar]
- Darmer, G. Rassenbildung bei Hippophaë rhamnoides (Sanddorn). Biol. Zentralbl. 1947, 66, 166–170. [Google Scholar]
- Arohonka, A.T.; Rousi, A. Karyotypes and C-bands of Shepherdia and Elaeagnus. Ann. Bot. Fenn. 1980, 17, 258–263. [Google Scholar]
- Ris, H.; Kubai, D.F. Chromosome structure. Annu. Rev. Genet. 1970, 4, 263–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebel, B.R. Chromosome Structure. Bot. Rev. 1939, 5, 563–626. [Google Scholar] [CrossRef]
- Peska, V.; Garcia, S. Origin, diversity, and evolution of telomere sequences in plants. Front. Plant Sci. 2020, 11, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Xiang, S.; Guo, Q.; Jin, W.; Cai, Z.; Liang, G. Molecular cytogenetic analysis of genome-specific repetitive elements in Citrus clementina Hort. Ex Tan. and its taxonomic implications. BMC Plant Biol. 2019, 19, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.C.; Pellerin, R.J.; Waminal, N.E.; Yang, T.J.; Kim, H.H. Pre-labelled oligo probe-FISH karyotype analyses of four Araliaceae species using rDNA and telomeric repeat. Genes Genom. 2019, 41, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Waminal, N.E.; Pellerin, R.J.; Kim, N.S.; Jayakodi, M.; Park, J.Y.; Yang, T.J.; Kim, H.H. Rapid and efficient FISH using pre-labeled oligomer probes. Sci. Rep. 2018, 8, 8224–8233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, I.V.; Van Laere, K.; Van Roy, N.; Khrustaleva, L.I. Towards a FISH-based karyotype of Rosa L. (Rosaceae). Comp. Cytogenet. 2016, 10, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Peska, V.; Fajkus, P.; Fojtova, M.; Dvorackova, M.; Hapala, J.; Dvoracek, V.; Polanska, P.; Leitch, A.R.; Sykorova, E.; Fajkus, J. Characterisation of an unusual telomere motif (TTTTTTAGGG)(n) in the plant Cestrum elegans (Solanaceae), a species with a large genome. Plant J. 2015, 82, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.G.; Friesen, N.; Heslop-Harrison, J.S. Molecular cytogenetic analysis of Podocarpus and comparison with other gymnosperm species. Ann. Bot. 2002, 89, 483–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykorova, E.; Lim, K.Y.; Fajkus, J.; Leitch, A.R. The signature of the Cestrum genome suggests an evolutionary response to the loss of (TTTAGGG)n telomeres. Chromosoma 2003, 112, 164–172. [Google Scholar] [CrossRef]
- Sykorova, E.; Lim, K.Y.; Chase, M.W.; Knapp, S.; Leitch, I.J.; Leitch, A.R.; Fajkus, J. The absence of Arabidopsis-type telomeres in Cestrum and closely related genera Vestia and Sessea (Solanaceae): First evidence from eudicots. Plant J. 2003, 34, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Nahm, S.H.; Kim, Y.M.; Kim, B.D. Characterization and molecular genetic mapping of microsatellite loci in pepper. Theor. Appl. Genet. 2004, 108, 619–627. [Google Scholar] [CrossRef]
- Ma, Z.Q.; Röder, M.; Sorrells, M.E. Frequencies and sequence characteristics of di-, tri-, and tetra-nucleotide microsatellites in wheat. Genome 1996, 39, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Matsumoto, S.; Kojima, S.; Ohshima, K.; Okada, N.; Machida, Y. Molecular characterization of a short interspersed repetitive element from tobacco that exhibits sequence homology to specific tRNAs. Proc. Natl. Acad. Sci. USA 1993, 90, 6562–6566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falistocco, E.; Ferradini, N. Advances in the cytogenetics of Annonaceae, the case of Annona cherimola L. Genome 2020, 63, 357–364. [Google Scholar] [CrossRef] [PubMed]
- de Melo, N.F.; Guerra, M. Variability of the 5S and 45S rDNA sites in Passiflora L. species with distinct base chromosome numbers. Ann. Bot. 2003, 92, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, M.; Zhang, L.; Li, K.; Sun, J.; Li, Z.; Han, Y. Karyotypic stability of Fragaria (strawberry) species revealed by cross-species chromosome painting. Chromosome Res. 2021, 29, 285–300. [Google Scholar] [CrossRef]
- Schmidt, T.; Heitkam, T.; Liedtke, S.; Schubert, V.; Menzel, G. Adding color to a century-old enigma: Multi-color chromosome identification unravels the autotriploid nature of saffron (Crocus sativus) as a hybrid of wild Crocus cartwrightianus cytotypes. New Phytol. 2019, 222, 1965–1980. [Google Scholar] [CrossRef] [PubMed]
- Frello, S.; Heslop-Harrison, J.S. Repetitive DNA sequences in Crocus vernus Hill (Iridaceae): The genomic organization and distribution of dispersed elements in the genus Crocus and its allies. Genome 2000, 43, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Twab, M.H.; Kondo, K. FISH physical mapping of 5S, 45S and Arabidopsis-type telomere sequence repeats in Chrysanthemum zawadskii showing intra-chromosomal variation and complexity in nature. Chromosome Bot. 2006, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Grabowska-Joachimiak, A.; Mosiolek, M.; Lech, A.; Góralski, G. C-Banding/DAPI and in situ hybridization reflect karyotype structure and sex chromosome differentiation in Humulus japonicus Siebold & Zucc. Cytogenet. Genome Res. 2011, 132, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, S.; Yu, L.; Li, N.; Li, S.; Zhang, Y.; Qin, R.; Gao, W.; Deng, C. Cloning and physical localization of male-biased repetitive DNA sequences in Spinacia oleracea (Amaranthaceae). Comp. Cytogenet. 2021, 15, 101–118. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Liu, J.; He, Z. Oligo-FISH Can Identify Chromosomes and Distinguish Hippophaë rhamnoides L. Taxa. Genes 2022, 13, 195. https://doi.org/10.3390/genes13020195
Luo X, Liu J, He Z. Oligo-FISH Can Identify Chromosomes and Distinguish Hippophaë rhamnoides L. Taxa. Genes. 2022; 13(2):195. https://doi.org/10.3390/genes13020195
Chicago/Turabian StyleLuo, Xiaomei, Juncheng Liu, and Zhoujian He. 2022. "Oligo-FISH Can Identify Chromosomes and Distinguish Hippophaë rhamnoides L. Taxa" Genes 13, no. 2: 195. https://doi.org/10.3390/genes13020195