Selection of References for microRNA Quantification in Japanese Flounder (Paralichthys olivaceus) Normal Tissues and Edwardsiella tarda-Infected Livers

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Candidate miRNA References Selection
2.2.1. miRNA-Seq for Normal Tissues and Bacterial Infected Livers of Japanese Flounder
2.2.2. Published miRNA References for Teleost Species
2.2.3. Commonly Used miRNA References for qRT-PCR: U6, 5S rRNA, and 18S rRNA
2.3. RNA Extraction and miRNA cDNA Synthesis
2.4. qRT-PCR Analysis
2.5. Statistical Analysis
2.6. Reference miRNA Validation
3. Results
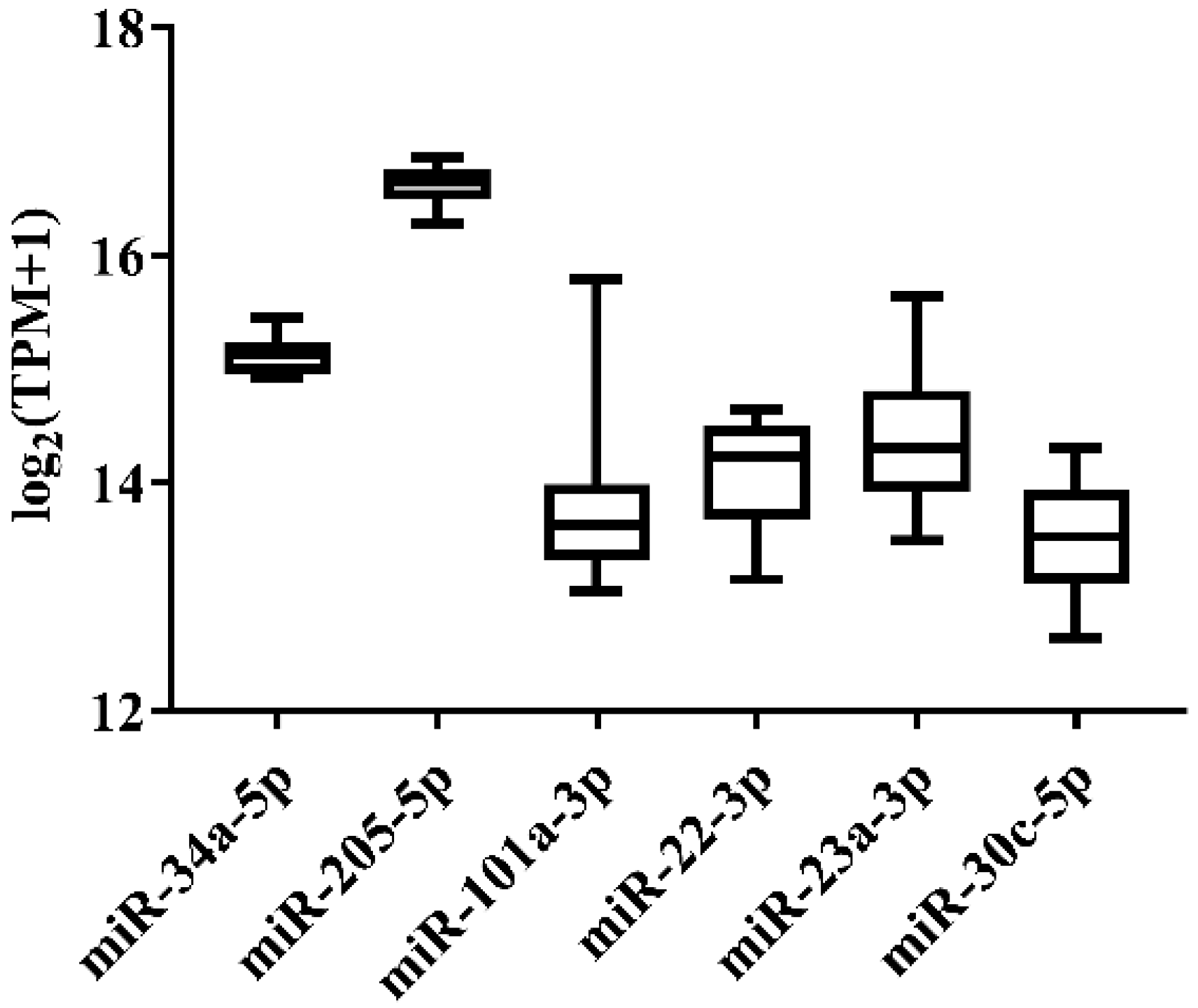
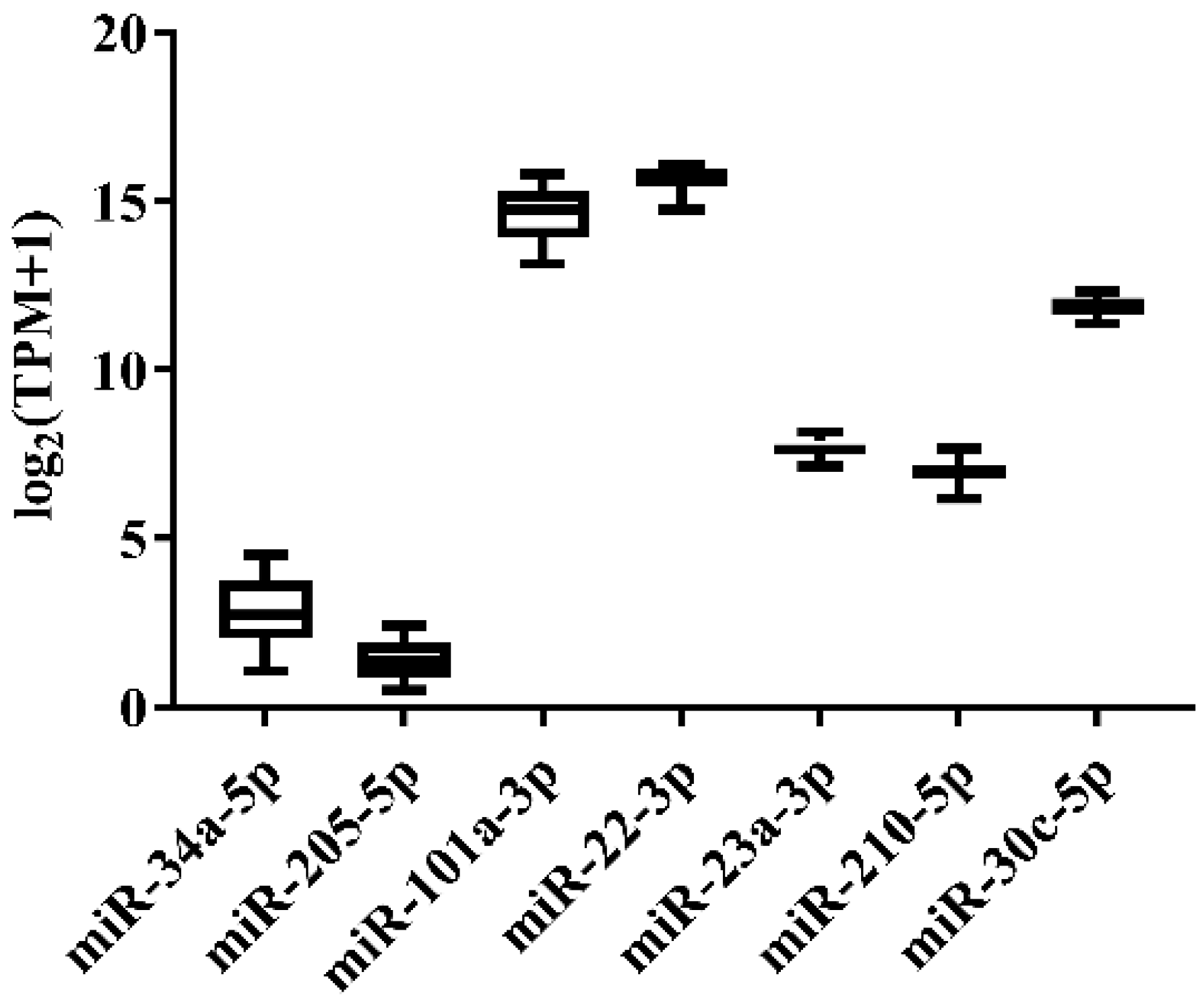
3.1. Candidate Reference Selection for Japanese Flounder miRNA Quantification
3.2. Amplification Efficiency of the Candidate References by qRT-PCR
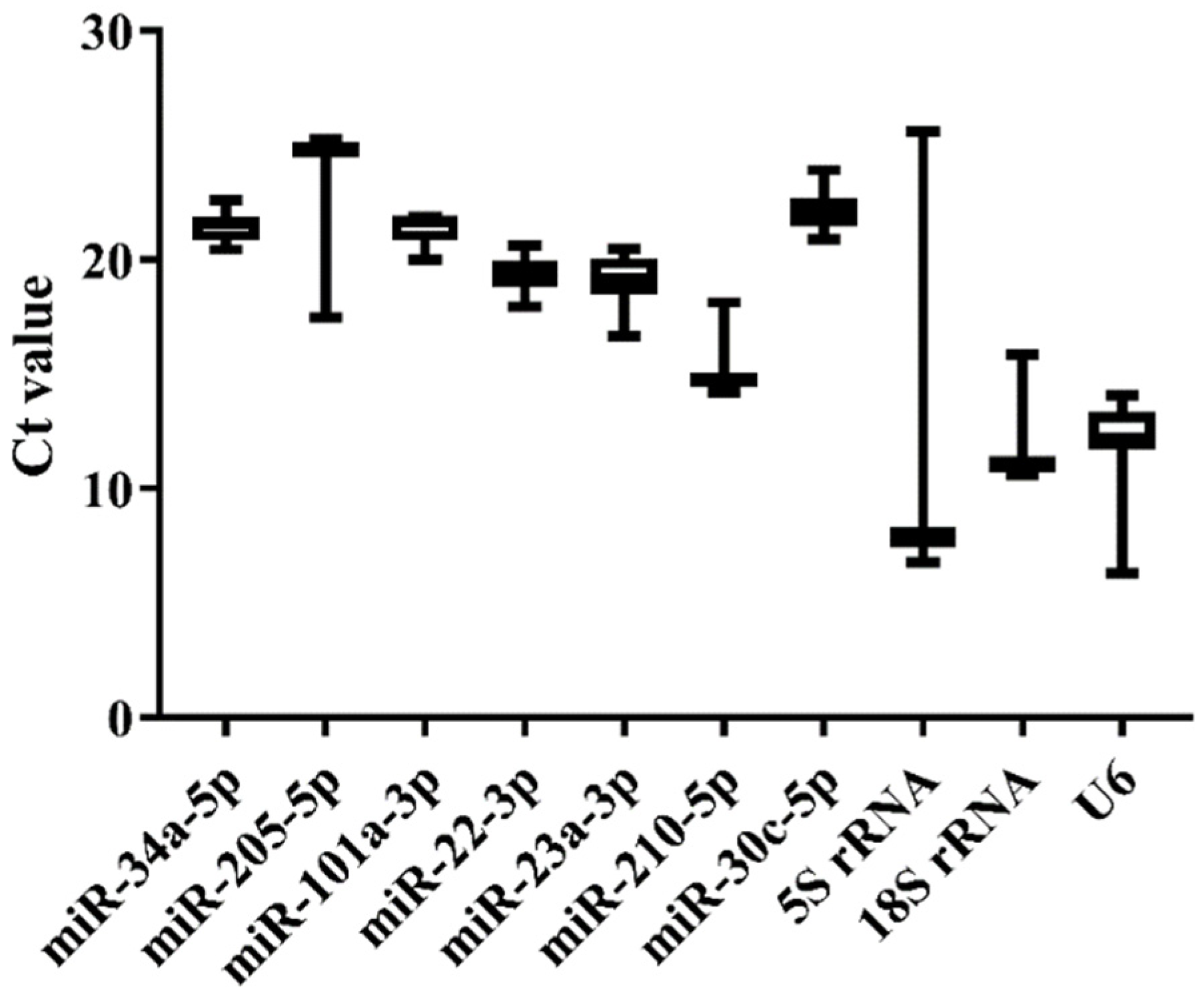
3.3. Expression Stability of the Candidate References among Normal Tissues of Japanese Flounder by qRT-PCR
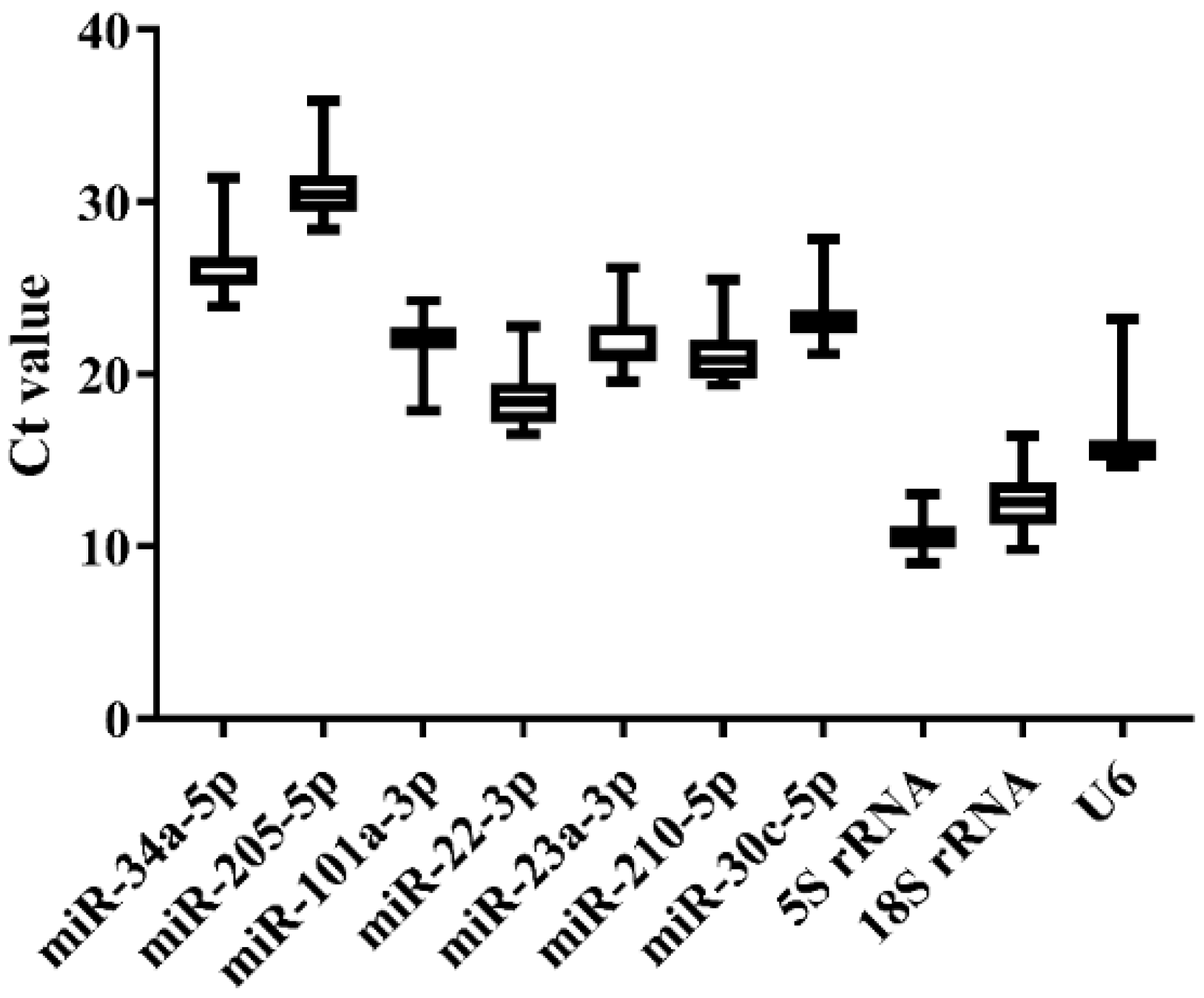
3.4. Expression Stability of Candidate References in Japanese Flounder Livers Injected with E. tarda or Ringer’s Solution by qRT-PCR
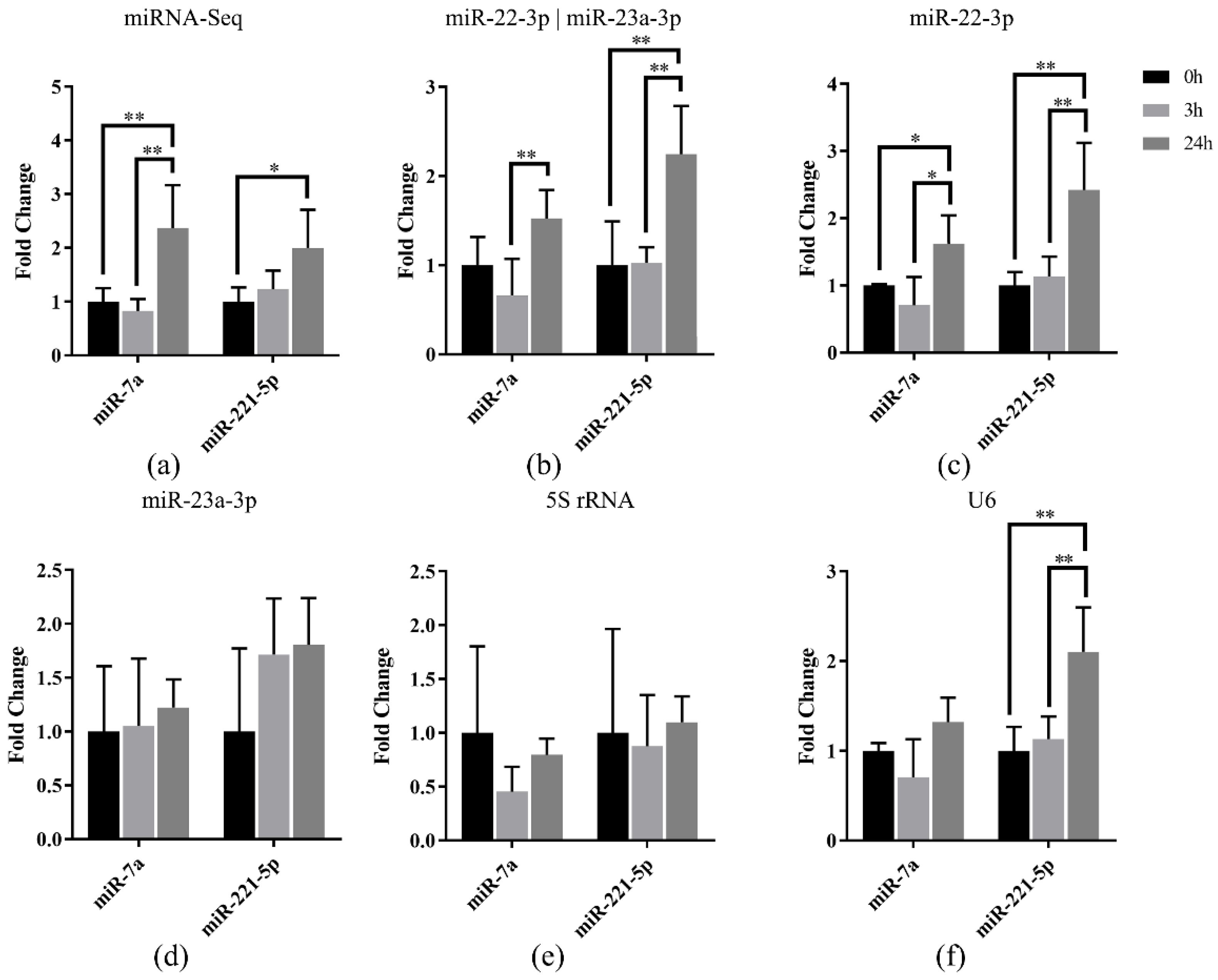
3.5. Reference miRNA Validation in Japanese Flounder Livers Infected with E. tarda by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flynt, A.S.; Lai, E.C. Biological principles of microRNA-mediated regulation: Shared themes amid diversity. Nat. Rev. Genet. 2008, 9, 831–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilahun, B.T.; Igor, B. MicroRNA in Teleost Fish. Genome Biol. Evol. 2014, 6, 1911–1937. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, J.; Hipfner, D.R.; Stark, A.; Russell, R.B.; Cohen, S.M. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 2003, 113, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leaman, D.; Chen, P.Y.; Fak, J.; Yalcin, A.; Pearce, M.; Unnerstall, U.; Marks, D.S.; Sander, C.; Tuschl, T.; Gaul, U. Antisense-mediated depletion reveals essential and specific functions of microRNAs in Drosophila development. Cell 2005, 121, 1097–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; de Bruijn, E.; Horvitz, H.R.; Kauppinen, S.; Plasterk, R.H.A. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar] [CrossRef] [Green Version]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced Expression of the let-7 MicroRNAs in Human Lung Cancers in Association with Shortened Postoperative Survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reza, A.M.M.T.; Choi, Y.J.; Han, S.G.; Song, H.; Park, C.; Hong, K.; Kim, J.H. Roles of microRNAs in mammalian reproduction: From the commitment of germ cells to per-implantation embryos. Biol. Rev. 2019, 94, 415–438. [Google Scholar] [CrossRef]
- Olejniczak, M.; Kotowska-Zimmer, A.; Krzyzosiak, W. Stress-induced changes in miRNA biogenesis and functioning. Cell Mol. Life Sci. 2018, 75, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression profiling of mammalian microRNAs uncovers a subset of brain-expressed microRNAs with possible roles in murine and human neuronal differentiation. Genome Biol. 2004, 5, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Koo, S.; White, N.; Peralta, E.; Perera, R.J. Development of a micro-array to detect human and mouse microRNAs and characterization of expression in human organs. Nucleic Acids Res. 2004, 32, e188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Davoren, P.A.; Mcneill, R.E.; Lowery, A.J.; Kerin, M.J.; Miller, N. Identification of suitable endogenous control genes for microRNA gene expression analysis in human breast cancer. BMC Mol. Biol. 2008, 9, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Mackay, I.M.; Arden, K.E.; Andreas, N. Real-time PCR in virology. Nucleic Acids Res. 2002, 30, 1292–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oriol, T.; Ingrid, B.; Sarai, C.; Anna, C.; Armand, S.; Atsushi, A. Determination of Reference microRNAs for Relative Quantification in Porcine Tissues. PLoS ONE 2012, 7, e44413. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Yang, Q.W.; Zhao, W.J.; Du, Q.Y.; Chang, Z.J. Selection of suitable candidate genes for miRNA expression normalization in Yellow River Carp (Cyprinus carpio. var). Sci. Rep. 2019, 9, 8691. [Google Scholar] [CrossRef] [PubMed]
- Schwarzenbach, H.; Da Silva, A.M.; Calin, G.; Pantel, K. Data Normalization Strategies for MicroRNA Quantification. Clin. Chem. 2015, 61, 1333–1342. [Google Scholar] [CrossRef]
- Latham, P.G.J. Normalization of microRNA expression levels in quantitative RT-PCR assays: Identification of suitable reference RNA targets in normal and cancerous human solid tissues. RNA 2008, 14, 844–852. [Google Scholar] [CrossRef] [Green Version]
- Lardizábal, M.; Nocito, A.L.; Daniele, S.M.; Ornella, L.A.; Veggi, L.M. Reference Genes for Real-Time PCR Quantification of MicroRNAs and Messenger RNAs in Rat Models of Hepatotoxicity. PLoS ONE 2012, 7, e36323. [Google Scholar] [CrossRef] [PubMed]
- Johansen, I.; Andreassen, R. Validation of miRNA genes suitable as reference genes in qPCR analyses of miRNA gene expression in Atlantic salmon (Salmo salar). BMC Res. Notes 2014, 7, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.Y.; Shen, Y.B.; Fu, J.J.; Lu, L.Q.; Li, J.L. Determination of reference microRNAs for relative quantification in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2014, 36, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, Y.L.; Chen, D.X.; Wu, P.; Yi, T. Selection of Reference Genes for MicroRNA Quantitative Expression Analysis in Chinese Perch, Siniperca chuatsi. Int. J. Mol. Sci. 2015, 16, 8310–8323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreassen, R.; Rangnes, F.; Sivertsen, M.; Chiang, M.; Tran, M.; Molton, W.M.; Prunet, P. Discovery of miRNAs and Their Corresponding miRNA Genes in Atlantic Cod (Gadus morhua): Use of Stable miRNAs as Reference Genes Reveals Subgroups of miRNAs That Are Highly Expressed in Particular Organs. PLoS ONE 2016, 11, e0153324. [Google Scholar] [CrossRef]
- Liu, J.; Jia, E.; Shi, H.; Li, X.; Jiang, G.; Chi, C.; Liu, W.; Zhang, D. Selection of reference genes for miRNA quantitative PCR and its application in miR-34a/Sirtuin-1 mediated energy metabolism in Megalobrama amblycephala. Fish Physiol. Biochem. 2019, 45, 1663–1681. [Google Scholar] [CrossRef]
- Ni, F.F.; Yu, H.Y.; Liu, Y.Z.; Meng, L.H.; Yan, W.J.; Zhang, Q.Q.; Yu, H.Y.; Wang, X.B. Roles of piwil1 gene in gonad development and gametogenesis in Japanese flounder, Paralichthys olivaceus. Gene 2019, 701, 104–112. [Google Scholar] [CrossRef]
- Song, H.F.; Xing, C.J.; Lu, W.; Liu, Z.Y.; Wang, X.B.; Cheng, J.; Zhang, Q.Q. Rapid evolution of piRNA pathway and its transposon targets in Japanese flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100609. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Z.; Liu, Y.X.; Han, M.; Du, X.X.; Liu, X.M.; Zhang, Q.Q.; Liu, J.X. Edwardsiella tarda-induced miR-7a functions as a suppressor in PI3K/AKT/GSK3β signaling pathway by targeting insulin receptor substrate-2 (IRS2a and IRS2b) in Paralichthys olivaceus. Fish Shellfish Immunol. 2019, 89, 477–485. [Google Scholar] [CrossRef]
- Yang, H.L.; Liu, J.; Huang, S.M.; Guo, T.T.; Deng, L.B.; Hua, W. Selection and evaluation of novel reference genes for quantitative reverse transcription PCR (qRT-PCR) based on genome and transcriptome data in Brassica napus L. Gene 2014, 538, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Czechowski, T.; Stitt, M.; Altmann, T.; Udvardi, M.K.; Scheible, W.-R.D. Genome-Wide Identification and Testing of Superior Reference Genes for Transcript Normalization in Arabidopsis. Plant Physiol. 2005, 139, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavala, E.; Reyes, D.; Deerenberg, R.; Vidal, R. Selection of reference genes for microRNA analysis associated to early stress response to handling and confinement in Salmo salar. Sci. Rep. 2017, 7, 1756. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.Y.; Zhang, Q.Q.; Qi, J.; Wang, Z.G.; Chen, Y.J.; Li, C.M.; Zhong, Q.W. Cloning and Expression Analysis of DMRT 1 Gene in Cynoglossus semilaevis. Nat. Sci. Ed. 2008, 54, 221–226. [Google Scholar] [CrossRef]
- Fu, Y.S.; Shi, Z.Y.; Wang, G.Y.; Li, W.J.; Zhang, J.L.; Jia, L. Expression and regulation of miR-1, -133a, -206a, and MRFs by thyroid hormone during larval development in Paralichthys olivaceus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2012, 161, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper-Excel-Based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034.1–0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.L.; Xiao, P.; Chen, D.L.; Xu, L.; Zhang, B.H. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Sun, Y.H.; Shi, R.; Clark, C.; Chiang, V.L. Novel and Mechanical Stress-Responsive MicroRNAs in Populus trichocarpa That Are Absent from Arabidopsis. Plant Cell 2005, 17, 2186–2203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanca, A.S.; Vicentini, R.; Ortiz-Morea, F.A.; Bem, L.E.D.; Silva, M.J.D.; Vincentz, M.; Nogueira, F.T. Identification and expression analysis of microRNAs and targets in the biofuel crop sugarcane. BMC Plant Biol. 2010, 10, 260. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A. Absolute quantification of mRNA using real-time reverse transcription polymerase chain reaction assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabo, A.; Perou, C.M.; Karaca, M.; Perreard, L. Statistical modeling for selecting housekeeper genes. Genome Biol. 2008, 5, 156–167. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′–3′) | Amplification Efficiency |
|---|---|---|
| miR-34a-5p-F | TGGCAGTGTCTTAGCTGGTTGT | 173.84% |
| miR-205-5p-F | TCCTTCATTCCACCGGAGTCTG | 90.74% |
| miR-101a-3p-F | TACAGTACTGTGATAACTGAAG | 96.22% |
| miR-22-3p-F | AAGCTGCCAGCTGAAGAACTGT | 99.98% |
| miR-23a-3p-F | ATCACATTGCCAGGGATTTCCA | 110.00% |
| miR-210-5p-F | AGCCACTGACTAACGCACATTG | 131.01% |
| miR-30c-5p-F | TGTAAACATCCTTGACTGGAAGCT | 220% |
| 5S rRNA-F | CCATACCACCCTGAACAC | 83.38% |
| 5S rRNA-R | CGGTCTCCCATCCAAGTA | |
| 18S rRNA-F | CCTGAGAAACGGCTACCACAT | 85.45% |
| 18S rRNA-R | CCAATTACAGGGCCTCGAAAG | |
| U6-F | TTGGAACGATACAGAGAAGATTAGC | 86.42% |
| Method | Ranking Order (Better–Good–Average) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Delta CT | miR-22-3p | miR-23a-3p | miR-101a-3p | miR-30c-5p | miR-34a-5p | miR-210-5p | 18S rRNA | miR-205-5p | U6 | 5S rRNA |
| BestKeeper | miR-34a-5p | miR-101a-3p | miR-22-3p | miR-30c-5p | miR-210-5p | miR-23a-3p | 18S rRNA | U6 | miR-205-5p | 5S rRNA |
| NormFinder | miR-22-3p | miR-210-5p | miR-23a-3p | miR-30c-5p | miR-101a-3p | 18S rRNA | miR-34a-5p | miR-205-5p | U6 | 5S rRNA |
| geNorm | miR-22-3p|miR-23a-3p | miR-101a-3p | miR-34a-5p | miR-30c-5p | miR-210-5p | 18S rRNA | miR-205-5p | U6 | 5S rRNA | |
| Recommended comprehensive ranking | miR-22-3p | miR-23a-3p | miR-101a-3p | miR-34a-5p | miR-30c-5p | miR-210-5p | 18S rRNA | miR-205-5p | U6 | 5S rRNA |
| geNorm | miR-22-3p|miR-23a-3p | miR-101a-3p | miR-34a-5p | miR-30c-5p | miR-210-5p | 18S rRNA | miR-205-5p | U6 | 5S rRNA |
|---|---|---|---|---|---|---|---|---|---|
| geNorm Stability value (M) | 0.652 | 0.770 | 0.899 | 0.997 | 1.123 | 1.263 | 1.493 | 1.736 | 2.514 |
| Method | Ranking Order (Better--Good--Average) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Delta CT | miR-210-5p | miR-22-3p | miR-23a-3p | miR-205-5p | miR-30c-5p | 18S rRNA | miR-34a-5p | U6 | miR-101a-3p | 5S rRNA |
| BestKeeper | 5S rRNA | miR-101a-3p | miR-34a-5p | miR-30c-5p | miR-210-5p | 18S rRNA | miR-205-5p | miR-22-3p | miR-23a-3p | U6 |
| NormFinder | miR-210-5p | miR-22-3p | miR-23a-3p | miR-205-5p | miR-30c-5p | 18S rRNA | miR-34a-5p | U6 | miR-101a-3p | 5S rRNA |
| geNorm | miR-22-3p|miR-23a-3p | miR-210-5p | 18S rRNA | miR-205-5p | miR-30c-5p | U6 | miR-34a-5p | miR-101a-3p | 5S rRNA | |
| Recommended comprehensive ranking | miR-210-5p | miR-22-3p | miR-23a-3p | miR-205-5p | miR-30c-5p | 18S rRNA | miR-34a-5p | 5S rRNA | miR-101a-3p | U6 |
| geNorm | miR-22-3p|miR-23a-3p | miR-210-5p | 18S rRNA | miR-205-5p | miR-30c-5p | U6 | miR-34a-5p | miR-101a-3p | 5S rRNA |
|---|---|---|---|---|---|---|---|---|---|
| geNorm Stability value (M) | 0.292 | 0.356 | 0.622 | 0.811 | 0.942 | 1.102 | 1.284 | 1.396 | 1.510 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Song, H.; Liu, Z.; Lu, W.; Zhang, Q.; Cheng, J. Selection of References for microRNA Quantification in Japanese Flounder (Paralichthys olivaceus) Normal Tissues and Edwardsiella tarda-Infected Livers. Genes 2022, 13, 175. https://doi.org/10.3390/genes13020175
Liu S, Song H, Liu Z, Lu W, Zhang Q, Cheng J. Selection of References for microRNA Quantification in Japanese Flounder (Paralichthys olivaceus) Normal Tissues and Edwardsiella tarda-Infected Livers. Genes. 2022; 13(2):175. https://doi.org/10.3390/genes13020175
Chicago/Turabian StyleLiu, Saisai, Haofei Song, Zeyu Liu, Wei Lu, Quanqi Zhang, and Jie Cheng. 2022. "Selection of References for microRNA Quantification in Japanese Flounder (Paralichthys olivaceus) Normal Tissues and Edwardsiella tarda-Infected Livers" Genes 13, no. 2: 175. https://doi.org/10.3390/genes13020175