
Analysis of Genetic Diversity of Fescue Populations from the Highlands of Bolivia Using EST-SSR Markers



Abstract
:1. Introduction
2. Materials and Methods
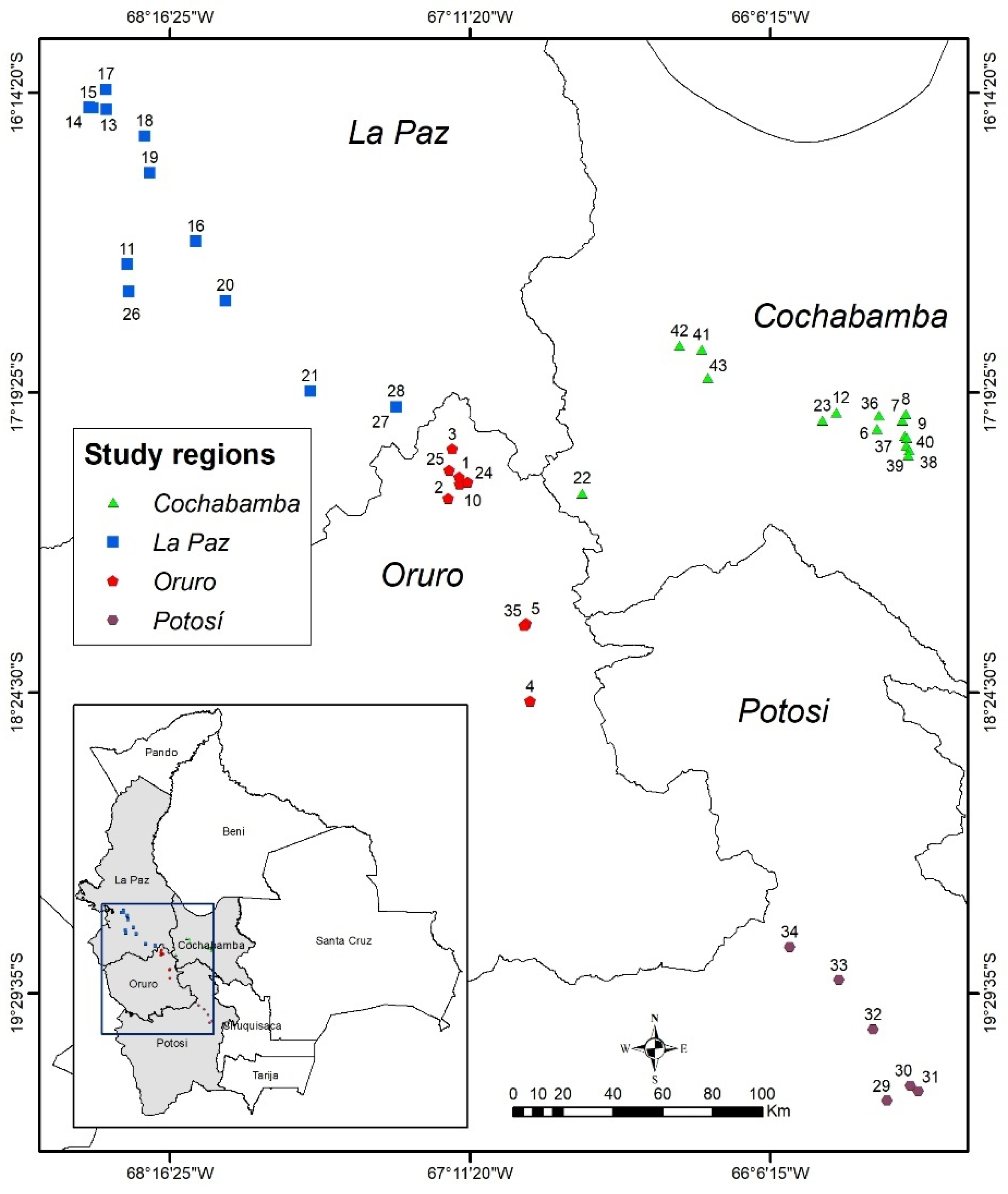
2.1. Plant Material
2.2. DNA Isolation
2.3. Designing and Screening Primer-Pairs
2.4. PCR Amplification and Capillary Electrophoresis
- (1)
- Initial denaturation step of 3 min at 95 °C, 38 cycles of 30 s denaturation at 94 °C, 30 s primer annealing at 60 °C, 45 s primer extension at 72 °C, and a final primer extension at 72 °C for 20 min (for NFA036).
- (2)
- Initial denaturation step of 3 min at 95 °C followed by nine touchdown cycles of 30 s denaturation at 94 °C, 30 s primer annealing at 58 °C reduced by 1 °C every cycle and 45 s extension at 72 °C. This was followed by 29 cycles of 30 s denaturation at 94 °C, 30 s annealing at 48 °C and 45 s primer extension at 72 °C with a 20 min final extension step at 72 °C (for FES09, FES14, NFA094, NFA142, NFA147 and NFA150).
- (3)
- Initial denaturation step of 3 min at 95 °C followed by nine touchdown cycles of 30 s denaturation at 94 °C, 30 s annealing at 59 °C reduced by 1 °C every cycle and 45 s primer extension at 72 °C. This was followed by 29 cycles of 30 s denaturation at 94 °C, 30 s annealing at 49 °C and 45 s primer extension at 72 °C with a 20 min final extension step at 72 °C (for FES04 and NFA126).
- (4)
- Initial denaturation step of 3 min at 95 °C followed by nine touchdown cycles of 30 s denaturation at 94 °C, 30 s annealing at 60 °C reduced by 1 °C every cycle and 45 s primer extension at 72 °C. This was followed by 29 cycles of 30 s denaturation at 94 °C, 30 s annealing at 50 °C and 45 s primer extension at 72 °C with a 20 min final extension step at 72 °C (for FES13, FES24 and NFA136).
2.5. Allele Scoring and Evaluation of Polymorphism
3. Results
3.1. EST SSR Polymorphism and Genetic Diversity
3.2. Within-Population Genetic Diversity
3.3. Genetic Variation among Populations and Groups of Populations
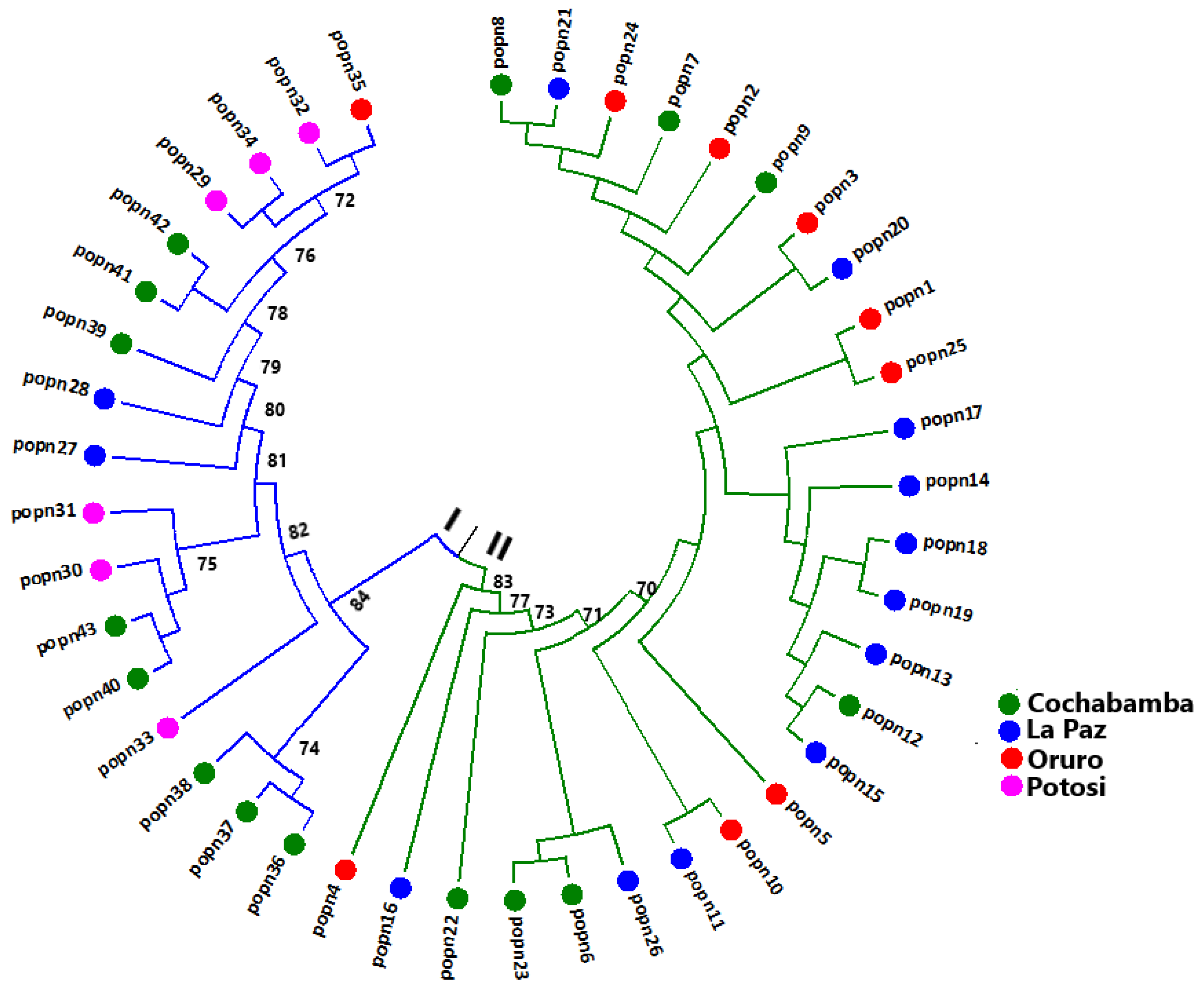
3.4. Genetic Distance and Cluster Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smarda, P.; Stancik, D. Ploidy Level variability in South American fescues (Festuca L., Poaceae): Use of flow cytometry in up to 5 1/2 –year-old caryopses and herbarium specimens. Plant Biol. 2006, 8, 73–80. [Google Scholar] [CrossRef]
- Clayton, W.D.; Renvoize, S.A. Genera Graminum Grasses of the World, 1st ed.; Kew Bulletin Additional Series XIII; Royal Botanical Garden: Kew, UK, 1986; 389p. [Google Scholar]
- Watson, L.; Dallwitz, M.J. The Grass Genera of the World; CAB International: Wallingford, UK, 1992; 1038p. [Google Scholar]
- Inda, L.A.; Segarra-Moragues, J.G.; Müller, J.; Peterson, P.M.; Catalán, P. Dated historical biogeography of the temperate Loliinae (Poaceae, Pooideae) grasses in the northern and southern hemispheres. Mol. Phylogenet. Evol. 2008, 46, 932–957. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T. Festuca. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Stancik, D. Las especies del género Festuca (Poaceae) en Colombia. Darwiniana 2003, 41, 93–153. [Google Scholar]
- Jørgensen, P.M.; Nee, M.H.; Beck, S.G.; Arrázola, S.; Saldias, M. Catálogo de las Plantas Vasculares de Bolivia; Missouri Botanical Garden Press: St. Louis, MO, USA, 2014; pp. 1064–1066. [Google Scholar]
- Mercado, M.; Meneses, R.I.; Delgado, D. Festuca soratana. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds.; Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 216–217. [Google Scholar]
- Meneses, R.I.; Mercado, M.; Delgado, D. Festuca cochabambana. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds.; Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 214–215. [Google Scholar]
- Meneses, R.I.; Mercado, M.; Delgado, D. Festuca potosiana. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds.; Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 81–82. [Google Scholar]
- Mercado, M.; Meneses, R.I. Festuca buchtienii. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds.; Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 33–34. [Google Scholar]
- Meneses, R.I.; Mercado, M.; Delgado, D. Festuca boliviana. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds. Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 78–79. [Google Scholar]
- Mercado, M.; Meneses, R.I.; Delgado, D. Festuca petersonii. In Libro Rojo de la Flora Amenazada de Bolivia, 1st ed.; Navarro, G., Arrázola, S., Atahuachi, M., De La Barra, N., Mercado, M., Eds.; Ministerio de Medio Ambiente y Agua. Rumbol SRL: La Paz, Bolivia, 2012; Volume 1, pp. 80–81. [Google Scholar]
- Campero, J.R. Llama (Lama glama L.) and Guanaco (Lama guanicoe M.): General perspective. In FAO-ICAR Seminar on Camelids: Current Status of Genetic Resources, Recording and Production Systems in African, Asian and American Camelids; Cardellino, R., Rosati, A., Mosconi, C., Eds.; ICAR: Sousse, Tunisia, 2004; pp. 11–18. [Google Scholar]
- Alzerreca, H. (Ed.) Campos Nativos de Pastoreo de Bolivia. In XV Reunión Nacional de Asociación Boliviana de Producción Animal (ABOPA); ABOPA: Oruro, Bolivia, 2004; pp. 217–229. [Google Scholar]
- Alzérreca, H.; Lara, R. Evaluación de praderas nativas en el altiplano central y oeste del Departamento de Oruro. Primera Reunión Nacional en Praderas Nativas de Bolivia. In Programa de Auto-Desarrollo Campesino; Corporación Desarrollo de Oruro (PAC, CORDEOR): Oruro, Bolivia, 1988; pp. 3–11. [Google Scholar]
- Monteiro, J.A.F.; Hiltbrunner, E.; Körner, C. Functional morphology and microclimate of Festuca orthophylla, the dominant tall tussock grass in the Andean Altiplano. J. Flora-Morphol. Distrib. Funct. Ecol. Plant. 2011, 206, 387–396. [Google Scholar] [CrossRef]
- Genin, D.; Alzérreca, H. Campos nativos de pastoreo y producción animal en la puna semiárida y árida andina. Sécheresse 2006, 17, 265–274. [Google Scholar]
- Mercado, M.; Arrázola, S.; Gutiérrez, F.; Ramírez, K.; Gonzales, J.; Atahuachi, M.; Vargas, N.J.; Burgos, J.; Ovando, K.; Campos, H.; et al. Guía Ilustrada de Especies Forrajeras Nativas de la Zona Andina en Bolivia. Proyecto Conservación y Manejo Sostenible de la Biodiversidad de los Recursos Genéticos Forrajeros de la Zona Andina de Bolivia; UMSS–FCAPFyV–FCyT–CISTEL–BASFOR–CBG–CIUF-CUD: Cochabamba, Bolivia, 2013. [Google Scholar]
- Merlo, F.E.; Ku, J.C.; Condori, R.; Pérez, L.; Albarracín, A. Efecto de la edad y época de año sobre el rendimiento y composición química del pastizal chilliwar Festuca dolychophylla en Tiahuanaco, Bolivia. In Proceedings of the XXII Reunión Nacional de Asociación Boliviana de Producción Animal (ABOPA), Santa Cruz, Bolivia, 11–13 October 2018; Meneses, R., Ed.; pp. 120–127. [Google Scholar]
- Genin, D.; Villca, Z.; Abasto, P. Diet selection and utilization by llama and sheep in a high altitude-arid rangeland of Bolivia. J. Range Manag. 1994, 47, 245–248. [Google Scholar] [CrossRef]
- Zapata, A. Las forrajeras nativas preferidas por los camélidos. Rev. Suma Tama 2005, 1, 1–20. [Google Scholar]
- Shahabzadeh, Z.; Mohammadi, R.; Darvishzadeh, R.; Jaffari, M. Genetic structure and diversity analysis of tall fescue populations by EST–SSR and ISSR markers. Mol. Biol. Rep. 2020, 47, 655–669. [Google Scholar] [CrossRef]
- Fjellheim, S.; Rognli, O.A. Genetic diversity within and among Nordic meadow fescue (Festuca pratensis Huds.) cultivars determined on the basis of AFLP markers. Crop. Sci. 2005, 45, 2081–2086. [Google Scholar] [CrossRef]
- Hand, M.L.; Cogan, N.O.; Forster, J.W. Molecular characterization and interpretation of genetic diversity within globally distributed germplasm collection of tall fescue (Festuca arundinacea Schreb.) and meadow fescue (F. pratensis Huds.). Theor. Appl. Genet. 2012, 124, 1127–1137. [Google Scholar] [CrossRef]
- Fasih, Z.; Farshadfar, M.; Safari, H. Genetic diversity evaluation of within and between populations for Festuca arundinacea by SSR markers. Int. J. Agric. Crop. Sci. 2013, 5, 1468–1472. [Google Scholar]
- Türpe, A.M. Species of the genus Festuca in Argentina (exception Patagonia). Darwiniana 1969, 15, 191–281. [Google Scholar]
- Renvoize, S.A. Gramineas de Bolivia; Royal Botanic Garden: Kew, UK, 1998. [Google Scholar] [CrossRef]
- Stancik, D. New records of the genus Festuca L. (Poaceae) for Colombia and Venezuela. Caldasia 2001, 23, 337–339. [Google Scholar]
- Stancik, D.; Peterson, P. A revision of Festuca (Poaceae: Loliinae) in South American Páramos. Contrib. United States Natl. Herb. 2007, 56, 1–184. [Google Scholar]
- Ospina, J.C.; Aliscioni, S.S.; Denham, S.S. A revision of Festuca (Loliinae, Pooideae, Poaceae) in Chile. Phytotaxa 2015, 223, 1–66. [Google Scholar] [CrossRef]
- Mamani, C. Producción de Forrajes y Leguminosas en el Altiplano Boliviano. Resumen de Experiencias en Seis Años de Trabajo Entre el Centro Experimental Agropecuario Condoriri e Instituciones del Fundo Universitario “La Violeta”, Proyecto AgroLeg (CIAT-CIF-CIFP-SEFO); Meneses, R., Barrientos, E., Eds.; Centro Experimental Agropecuario Condoriri: Cochabamba, Bolivia, 2003. [Google Scholar]
- Mamani-Linares, W.; Gallo, C.; Pulido, R. Composición Botánica y Contenido Nutricional de Pasturas Nativas en Periodo Seco en el Altiplano. Documento Presentado en el XXXVIII Congreso Sociedad Chilena de Producción Animal; Libro de resúmenes, Instituto de Ciencia Animal, Facultad Ciencias Veterinarias, Universidad Austral de Chile: Valdivia, Chile, 2013. [Google Scholar]
- Ustariz, K.; Geleta, M.; Hovmalm, H.P.; Gutierrez, F.; Beltrán, J.A.R.; Ortiz, R. Mineral composition and nutritive value of Festuca ecotypes’ originated from the highland region of Bolivia and cultivars from Argentina. Aust. J. Crop. Sci. 2019, 1, 1650–1658. [Google Scholar] [CrossRef]
- Genin, D.; Tichit, M. Degradability of Andean range forages in llamas and sheep. J. Range Manag. 1997, 50, 381–385. [Google Scholar] [CrossRef]
- Kölliker, R.; Stadelmann, F.J.; Reidy, B.; Nösberger, J. Genetic variability of forage grass cultivars: A comparison of Festuca pratensis Huds., Lolium perenne L., and Dactylis glomerata L. Euphytica 1999, 106, 261–270. [Google Scholar] [CrossRef]
- Goldstein, D.; Schlötterer, C. Microsatellites: Evolution and Application, 1st ed.; Oxford University Press Inc.: New York, NY, USA, 1999. [Google Scholar]
- Saha, M.C.; Mian, M.R.; Eujayl, I.; Zwonitzer, J.C.; Wang, L.; May, G.D. Tall fescue EST-SSR markers with transferability across several grass species. Theor. Appl. Genet. 2004, 109, 783–791. [Google Scholar] [CrossRef]
- Thiel, T.; Michalek, W.; Varshney, R.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theor. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef]
- Peakall, R.; Gilmore, S.; Keys, W.; Morgante, M.; Rafalski, A. Cross-species amplification of soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume genera: Implications for the transferability of SSRs in plants. Mol. Biol. Evol. 1998, 15, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaitan-Solis, E.; Duque, M.C.; Edwards, K.J.; Tohme, J. Microsatellite repeats in common bean (Phaseolus vulgaris). Crop. Sci. 2002, 42, 2128–2136. [Google Scholar] [CrossRef]
- Saha, M.C.; Cooper, J.D.; Mian, M.R.; Chekhovskiy, K.; May, G.D. Tall fescue genomic SSR markers: Development and transferability across multiple grass species. Theor. Appl. Genet. 2006, 113, 1449–1458. [Google Scholar] [CrossRef]
- Elazreg, H.; Ghariani, S.; Chtourou-Ghorbel, N.; Chakroun, M.; Trifi-Farah, N. SSRs transferability and genetic diversity of Tunisian Festuca arundinacea and Lolium perenne. Bioch. Syst. Ecol. 2011, 39, 79–87. [Google Scholar] [CrossRef]
- Teshome, A.; Bryngelsson, T.; Dagne, K.; Geleta, M. Assessment of genetic diversity in Ethiopian field pea (Pisum sativum L.) accessions with newly developed EST-SSR markers. BMC Genet. 2015, 16, 102. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Rustgi, S.; Sharma, S.; Singh, R.; Kumar, N.; Balyan, H.S. Transferable EST-SSR markers for the study of polymorphism and genetic diversity in bread wheat. Mol. Genet. Genom. 2003, 270, 315–323. [Google Scholar] [CrossRef]
- Zhou, Q.; Luo, D.; Ma, L.; Xie, W.; Wang, Y.; Wang, Y.; Liu, Z. Development and cross-species transferability of EST-SSR markers in Siberian wildrye (Elymus sibiricus L.) using Illumina sequencing. Sci. Rep. 2016, 6, 20549. [Google Scholar] [CrossRef] [Green Version]
- Pashley, C.H.; Ellis, J.R.; McCauley, D.E.; Burke, J.M. EST databases as a source for molecular markers: Lessons from Helianthus. J. Hered. 2006, 97, 381–388. [Google Scholar] [CrossRef]
- Ustariz, K. Building Knowledge for a Forage Breeding Program on Native Festuca Species in the Highlands of Bolivia. Acta Universitatis Agriculturae Sueciae. Ph.D. Thesis, Swedish University of Agricultural Sciences, Alnarp, Sweden, 2019. [Google Scholar]
- Navarro, G.; Maldonado, M. Geografía Ecológica de Bolivia: Vegetación y Ambientes Acuáticos; Centro de Ecología Simón I. Patiño, Departamento de Difusión: Cochabamba, Bolivia, 2002; 719p. [Google Scholar]
- Martins, W.S.; César, D.; Lucas, S.; de Souza, N.K.F.; Bertioli, D.J. WebSat a web software for microsatellite marker development. Bioinformation 2009, 3, 282–283. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3 new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Version 1.32: Microsoft Windows–Based Freeware for Population Genetic Analysis, Quick User Guide; Center for International Forestry Research, University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sneath, P.H.A.; Sokal, R.R. Numerical Taxonomy; Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Varshney, R.K.; Graner, A.; Sorrells, M.E. Genic microsatellite markers in plants: Features and applications. Trends Biotechnol. 2005, 23, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Mian, M.A.R.; Saha, M.C.; Hopkins, A.A.; Wang, Z.Y. Use of tall fescue EST-SSR markers in phylogenetic analysis of cool-season forage grasses. Genome 2005, 48, 637–647. [Google Scholar] [CrossRef]
- Gadissa, F.; Tesfaye, K.; Dagne, K.; Geleta, M. Genetic diversity and population structure analyses of Plectranthus edulis (Vatke) Agnew collections from diverse agro-ecologies in Ethiopia using newly developed EST-SSRs marker system. BMC Genet. 2018, 19, 1. [Google Scholar] [CrossRef] [Green Version]
- Serbessa, T.B.; Dagne, W.K.; Teshome, G.A.; Geleta, D.M.; Tesfaye, G.K. Analyses of genetic diversity and population structure of anchote (Coccinia abyssinica (Lam.) Cogn.) using newly developed EST-SSR markers. Genet. Resour. Crop. Evol. 2021, 68, 2337–2350. [Google Scholar] [CrossRef]
- Peter-Schmid, M.K.I.; Boller, B.; Kölliker, R. Habitat and management affect genetic structure of Festuca pratensis but not Lolium multiflorum ecotype populations. Plant Breed. 2008, 127, 510–517. [Google Scholar] [CrossRef]
- Chombe, D.; Bekele, E.; Bryngelsson, T.; Teshome, A.; Geleta, M. Genetic structure and relationships within and between cultivated and wild korarima [Aframomum corrorima (Braun) P.C.M. Jansen] in Ethiopia as revealed by simple sequence repeat (SSR) markers. BMC Genet. 2017, 18, 72. [Google Scholar] [CrossRef]
- Stancik, D.; Peterson, P. A Revision of Festuca (Poaceae: Loliinae) in South American Paramos; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2007. [Google Scholar]



{kind=link}
{kind=link}
| Locus Name | Primer Sequence (5′–3′) | Repeat Motif | Expected Size | Observed Size Range | PIC |
|---|---|---|---|---|---|
| FES04 a | F: GTTGAGTGGAGTTGAGGTCACA | (TA) 15 | 191 | 168–180 | 0.22 |
| R:AAAGGAACGAGGAAACACTACG | |||||
| FES09 a | F: CCGGTGGGACCAGACTTA | (GC) 6 | 240 | 246 | N/A |
| R: CATGCACCCAGTTGTTGG | |||||
| FES13 a | F: GCAATCTTTCCTTGCATCATCT | (TGC) 7 | 163 | 143–183 | 0.55 |
| R: AGCAGCCTCAACAACTCCAG | |||||
| FES14 a | F: GCCCACACAACAAATGCAAC | (GT) 6 | 212 | 210–218 | 0.09 |
| R: CTCTCACTCTGCAGGTCGAT | |||||
| FES24 a | F: CTTACACCAGGAAACGGAAGAT | (CAG) 7 | 251 | 234–255 | 0.30 |
| R: TGGAGAAGCAAAATGTGAAGTG | |||||
| NFA036 b | F: AGAGGAAGAGCGAAAGAGCA | (GCC) 6 | 200 | 184–195 | 0.02 |
| R: CCCTGGTACTCGTGGATGTT | |||||
| NFA094 b | F: AGCTGAACTATGAGGCATGTCA | (GCA) 6 | 247 | 234–261 | 0.04 |
| R: ATCCCTTTCCAGCATTTACCTC | |||||
| NFA126 b | F: ACTACGTCTGCGAGTTCATTTG | (TA) 11 | 207 | 194 | N/A |
| R: GATCCACCGTTAGGAGAGTGTC | |||||
| NFA136 b | F: TGTGCAAGCAGAGACCTACACT | (TGTT) 9 | 237 | 195–259 | 0.09 |
| R: CAGCTGTGCTCCATTATCTGAG | |||||
| NFA142 b | F: CTTTGGACAAGGCAATGGAAT | (CAG) 7 | 229 | 210–255 | 0.52 |
| R: GTTGTTCTTCTGCGGGTAGTC | |||||
| NFA147 b | F: TGCAGTCGGTTAAGATCAAGAA | (CTG) 7 | 215 | 201–209 | 0.03 |
| R: AGTTGCAGTGAAGGTGCTGAAC | |||||
| NFA150 b | F: TGCAGTCGGTTAAGATCAAGAA | (CTG) 7 | 186 | 170–181 | 0.02 |
| R: GCAGAGCAATGGAGAGGTC |
| Locus | Na | Ne | H | I | Ht | Hs | Gst |
|---|---|---|---|---|---|---|---|
| FES04 | 6 | 2.13 | 0.53 | 0.95 | 0.53 | 0.25 | 0.53 |
| FES13 | 8 | 2.94 | 0.66 | 1.32 | 0.66 | 0.63 | 0.05 |
| FES14 | 4 | 1.15 | 0.13 | 0.31 | 0.13 | 0.11 | 0.16 |
| FES24 | 8 | 2.83 | 0.65 | 1.33 | 0.65 | 0.32 | 0.51 |
| NFA036 | 3 | 1.03 | 0.03 | 0.08 | 0.03 | 0.02 | 0.10 |
| NFA094 | 8 | 1.64 | 0.39 | 0.92 | 0.39 | 0.37 | 0.05 |
| NFA136 | 8 | 2.06 | 0.51 | 0.89 | 0.52 | 0.09 | 0.83 |
| NFA142 | 12 | 3.29 | 0.70 | 1.41 | 0.70 | 0.60 | 0.14 |
| NFA147 | 3 | 1.04 | 0.04 | 0.12 | 0.04 | 0.04 | 0.11 |
| NFA150 | 4 | 1.94 | 0.49 | 0.72 | 0.49 | 0.03 | 0.94 |
| Mean | 6.4 | 2.00 | 0.41 | 0.80 | 0.41 | 0.25 | 0.41 |
| St. Dev. | 2.9 | 0.82 | 0.26 | 0.49 | 0.07 | 0.05 |
| Allele | Locus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| NFA036 | NFA147 | FES14 | NFA150 | FES04 | FES13 | FES24 | NFA094 | NFA136 | NFA142 | |
| A | 0.002 a | 0.010 | 0.010 | 0.011 | 0.304 | 0.029 | 0.327 | 0.007 | 0.019 | 0.003 |
| B | 0.987 | 0.979 | 0.032 | 0.380 | 0.004 | 0.046 | 0.036 | 0.040 | 0.015 | 0.002 |
| C | 0.012 | 0.011 | 0.933 | 0.609 | 0.612 | 0.431 | 0.014 | 0.051 | 0.011 | 0.029 |
| D | 0.024 | 0.001 b | 0.021 | 0.084 | 0.047 | 0.059 | 0.345 | 0.187 | ||
| E | 0.051 | 0.022 | 0.489 | 0.041 | 0.604 | 0.049 | ||||
| F | 0.009 | 0.005 | 0.026 | 0.024 | 0.002 | 0.432 | ||||
| G | 0.004 | 0.054 | 0.775 | 0.001 c | 0.007 h | |||||
| H | 0.379 | 0.007 | 0.003 | 0.002 d | 0.281 e | |||||
| I | 0.003 | |||||||||
| J | 0.001 f | |||||||||
| K | 0.003 g | |||||||||
| L | 0.004 | |||||||||
| Pop | NPL | PPL | Na | Ne | H | I | Pop | NPL | PPL | Na | Ne | H | I | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 8 | 80 | 2.5 | 1.4 | 0.22 | 0.4 | 23 | 4 | 40 | 1.8 | 1.4 | 0.18 | 0.31 | |
| 2 | 5 | 50 | 2.4 | 1.4 | 0.21 | 0.38 | 24 | 5 | 50 | 2.3 | 1.4 | 0.22 | 0.39 | |
| 3 | 5 | 50 | 2.7 | 1.6 | 0.23 | 0.44 | 25 | 5 | 50 | 2.4 | 1.6 | 0.24 | 0.44 | |
| 4 | 7 | 70 | 3.2 | 1.7 | 0.26 | 0.53 | 26 | 5 | 50 | 2.4 | 1.6 | 0.24 | 0.44 | |
| 5 | 9 | 90 | 3.1 | 1.8 | 0.33 | 0.63 | 27 | 6 | 60 | 2.6 | 1.5 | 0.22 | 0.41 | |
| 6 | 8 | 80 | 3.1 | 1.8 | 0.33 | 0.61 | 28 | 6 | 60 | 2.5 | 1.5 | 0.22 | 0.41 | |
| 7 | 7 | 70 | 2.8 | 1.6 | 0.28 | 0.52 | 29 | 6 | 60 | 3 | 1.4 | 0.24 | 0.47 | |
| 8 | 9 | 90 | 3.1 | 1.7 | 0.33 | 0.6 | 30 | 5 | 50 | 2.2 | 1.3 | 0.18 | 0.32 | |
| 9 | 6 | 60 | 2.4 | 1.6 | 0.27 | 0.48 | 31 | 6 | 60 | 2.4 | 1.4 | 0.21 | 0.38 | |
| 10 | 5 | 50 | 2.6 | 1.5 | 0.22 | 0.41 | 32 | 5 | 50 | 2.2 | 1.4 | 0.21 | 0.38 | |
| 11 | 7 | 70 | 2.8 | 1.5 | 0.24 | 0.47 | 33 | 6 | 60 | 2 | 1.4 | 0.21 | 0.36 | |
| 12 | 8 | 80 | 2.4 | 1.7 | 0.31 | 0.53 | 34 | 8 | 80 | 3.3 | 1.7 | 0.33 | 0.63 | |
| 13 | 5 | 50 | 2.9 | 1.5 | 0.25 | 0.47 | 35 | 5 | 50 | 2.3 | 1.5 | 0.2 | 0.38 | |
| 14 | 6 | 60 | 2.6 | 1.5 | 0.24 | 0.46 | 36 | 7 | 70 | 2.1 | 1.6 | 0.27 | 0.45 | |
| 15 | 4 | 40 | 2.4 | 1.4 | 0.2 | 0.37 | 37 | 7 | 70 | 2.6 | 1.7 | 0.3 | 0.54 | |
| 16 | 5 | 50 | 2 | 1.3 | 0.17 | 0.29 | 38 | 7 | 70 | 2.6 | 1.5 | 0.25 | 0.46 | |
| 17 | 5 | 50 | 2.1 | 1.4 | 0.17 | 0.31 | 39 | 6 | 60 | 2.3 | 1.7 | 0.29 | 0.49 | |
| 18 | 8 | 80 | 3.6 | 1.7 | 0.31 | 0.63 | 40 | 6 | 60 | 2.4 | 1.4 | 0.22 | 0.4 | |
| 19 | 8 | 80 | 3.3 | 1.7 | 0.28 | 0.58 | 41 | 5 | 50 | 2.8 | 1.6 | 0.27 | 0.51 | |
| 20 | 6 | 60 | 2.6 | 1.5 | 0.24 | 0.44 | 42 | 6 | 60 | 2.3 | 1.6 | 0.26 | 0.46 | |
| 21 | 5 | 50 | 2.6 | 1.5 | 0.23 | 0.42 | 43 | 6 | 60 | 2 | 1.4 | 0.22 | 0.36 | |
| 22 | 5 | 50 | 2.5 | 1.5 | 0.22 | 0.42 | Mean | 61.2 | 2.6 | 1.5 | 0.24 | 0.45 |
| Source of Variation | DF * | Sum of Squares | Variance Components | % Age of Variation | Fixation Indices | p-Value |
|---|---|---|---|---|---|---|
| (a) Without grouping | ||||||
| Among populations | 42 | 28.647 | −0.43735 Va | −7.96 | FST: −0.07962 | 1.00 |
| Within populations | 473 | 2805.000 | 5.93023 Vb | 107.96 | ||
| Total | 515 | 2833.647 | 5.49289 | |||
| (b) By grouping the populations into two biogeographic provinces | ||||||
| Among groups | 1 | 0.960 | 0.00121 Va | 0.02 | FCT: 0.00022 | 0.24 |
| Among populations within groups | 41 | 27.688 | −0.43791 Vb | −7.97 | FST: −0.07949 | 1.00 |
| Within populations | 473 | 2805.000 | 5.93023 Vc | 107.95 | FSC: −0.07973 | 1.00 |
| Total | 515 | 2833.647 | 5.49354 | |||
| (c) By grouping the populations into four departments | ||||||
| Among groups | 3 | 3.614 | 0.00452 Va | 0.08 | FCT: 0.00082 | 0.01 |
| Among populations within groups | 39 | 25.033 | −0.44070 Vb | −8.02 | FST: −0.07939 | 1.00 |
| Within populations | 473 | 2805.000 | 5.93023 Vc | 107.94 | FSC: −0.08028 | 1.00 |
| Total | 515 | 2833.647 | 5.49406 | |||
| (d) By grouping the populations into two altitudinal ranges | ||||||
| Among groups | 1 | 0.240 | −0.00178 Va | −0.03 | FCT: −0.00032 | 0.88 |
| Among populations within groups | 41 | 28.407 | −0.43645 Vb | −7.95 | FST: −0.07979 | 1.00 |
| Within populations | 473 | 2805.000 | 5.93023 Vc | 107.98 | FSC: −0.07944 | 1.00 |
| Total | 515 | 2833.647 | 5.49201 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ustariz, K.; Geleta, M.; Persson Hovmalm, H.; Ortiz, R. Analysis of Genetic Diversity of Fescue Populations from the Highlands of Bolivia Using EST-SSR Markers. Genes 2022, 13, 2311. https://doi.org/10.3390/genes13122311
Ustariz K, Geleta M, Persson Hovmalm H, Ortiz R. Analysis of Genetic Diversity of Fescue Populations from the Highlands of Bolivia Using EST-SSR Markers. Genes. 2022; 13(12):2311. https://doi.org/10.3390/genes13122311
Chicago/Turabian StyleUstariz, Karina, Mulatu Geleta, Helena Persson Hovmalm, and Rodomiro Ortiz. 2022. "Analysis of Genetic Diversity of Fescue Populations from the Highlands of Bolivia Using EST-SSR Markers" Genes 13, no. 12: 2311. https://doi.org/10.3390/genes13122311