
Orchid NAC Transcription Factors: A Focused Analysis of CUPULIFORMIS Genes


Abstract
:1. Introduction
2. Materials and Methods
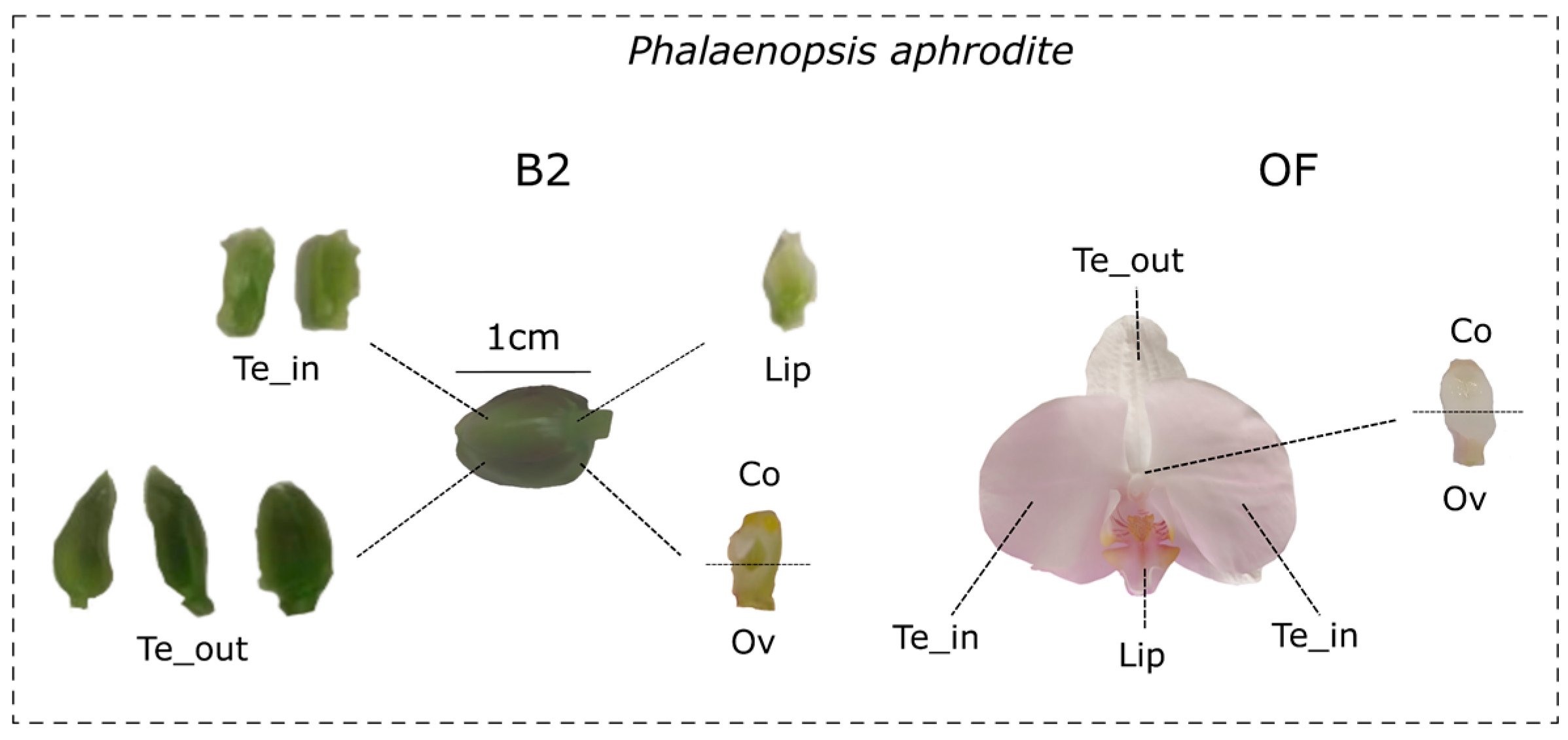
2.1. Plant Material
2.2. Identification, Phylogeny, and Conserved Motif Analysis of the Orchid NAC Genes
2.3. PCR Amplification and Sequencing of the CUP Genes of O. italica
2.4. PCR Amplification and Sequencing of P. aphrodite CUP Transcripts
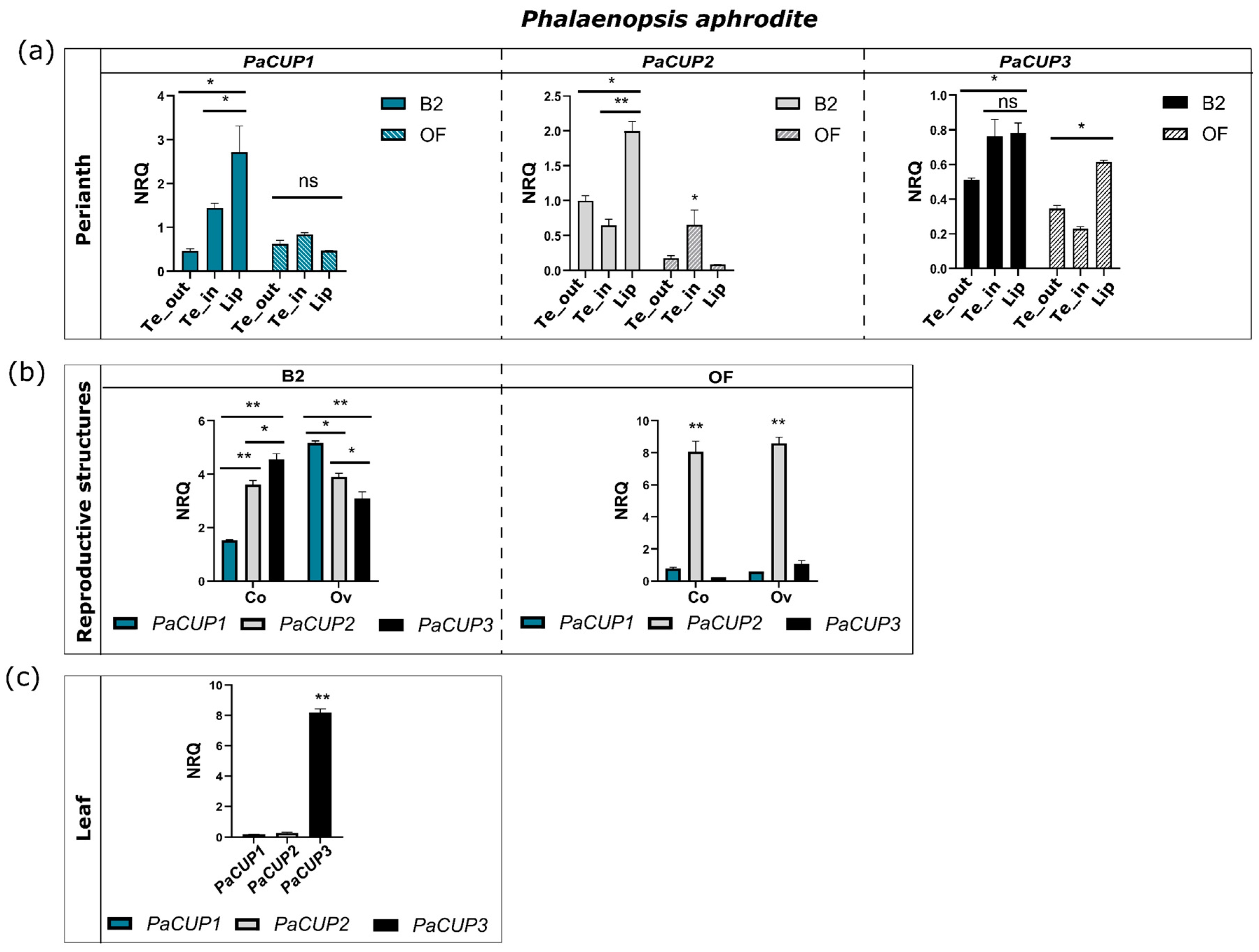
2.5. Expression Analysis
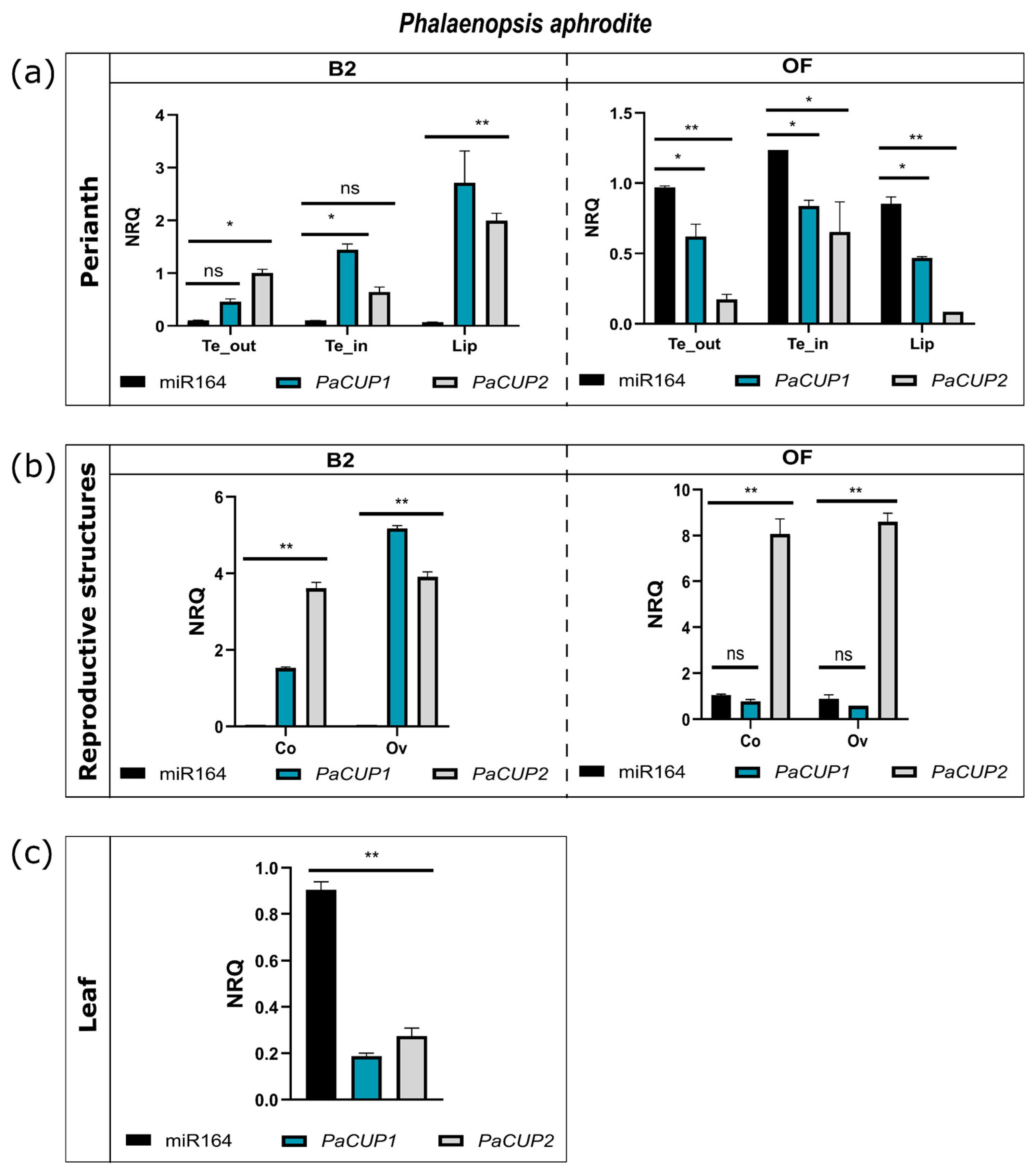
2.6. miR164 Analysis
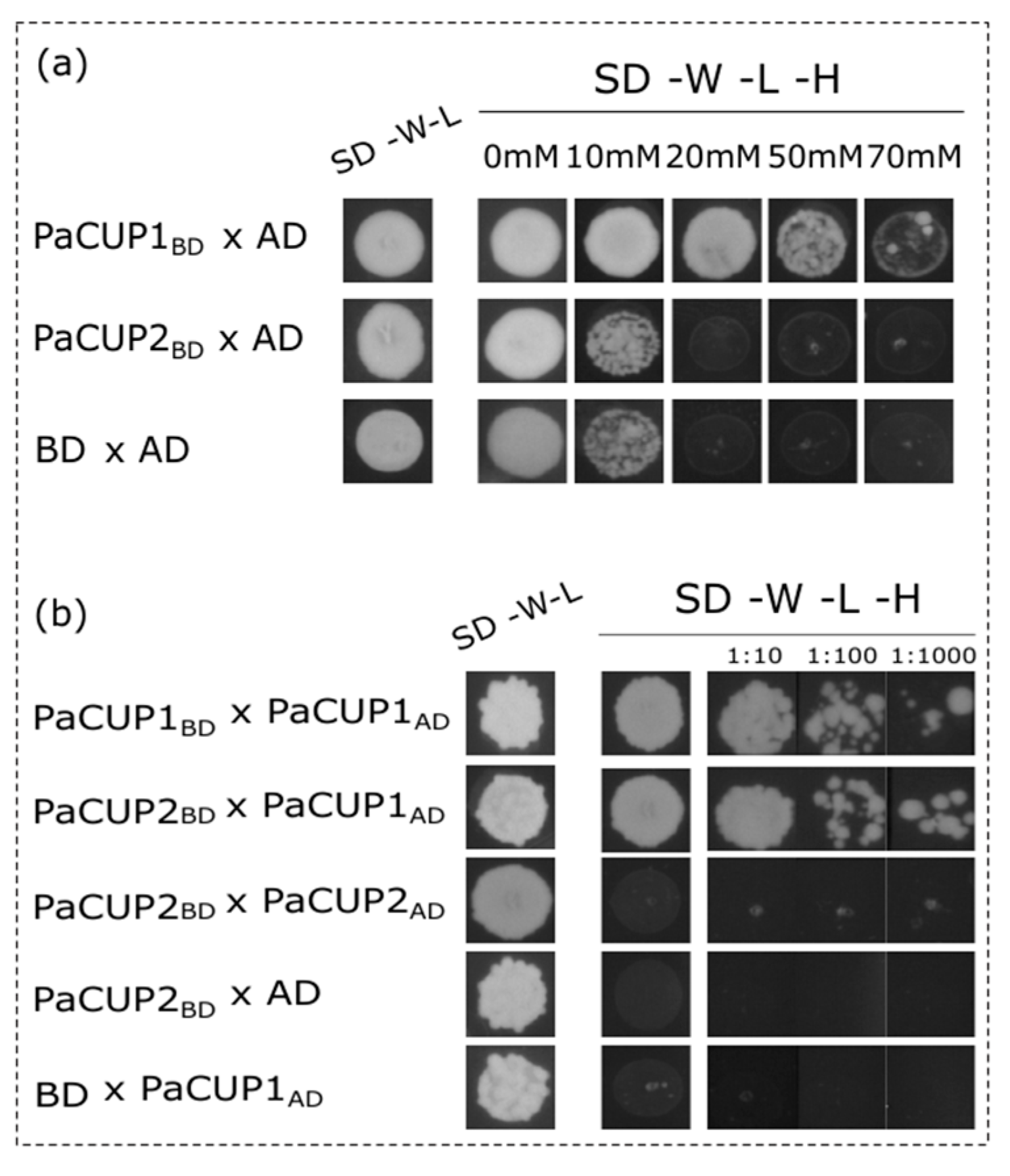
2.7. Yeast Two-Hybrid Analysis
3. Results and Discussion
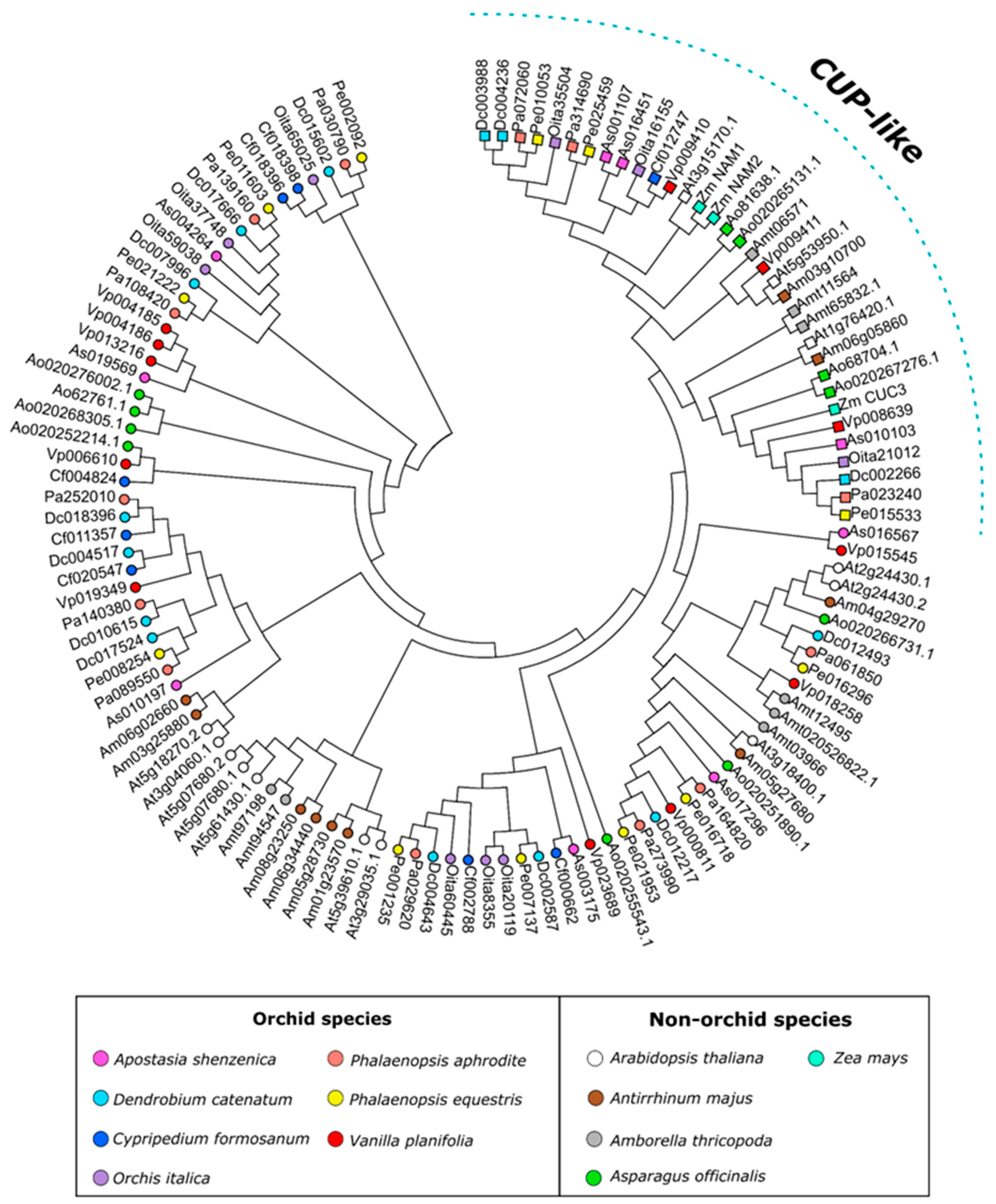
3.1. The NAC Transcription Factors of Orchids
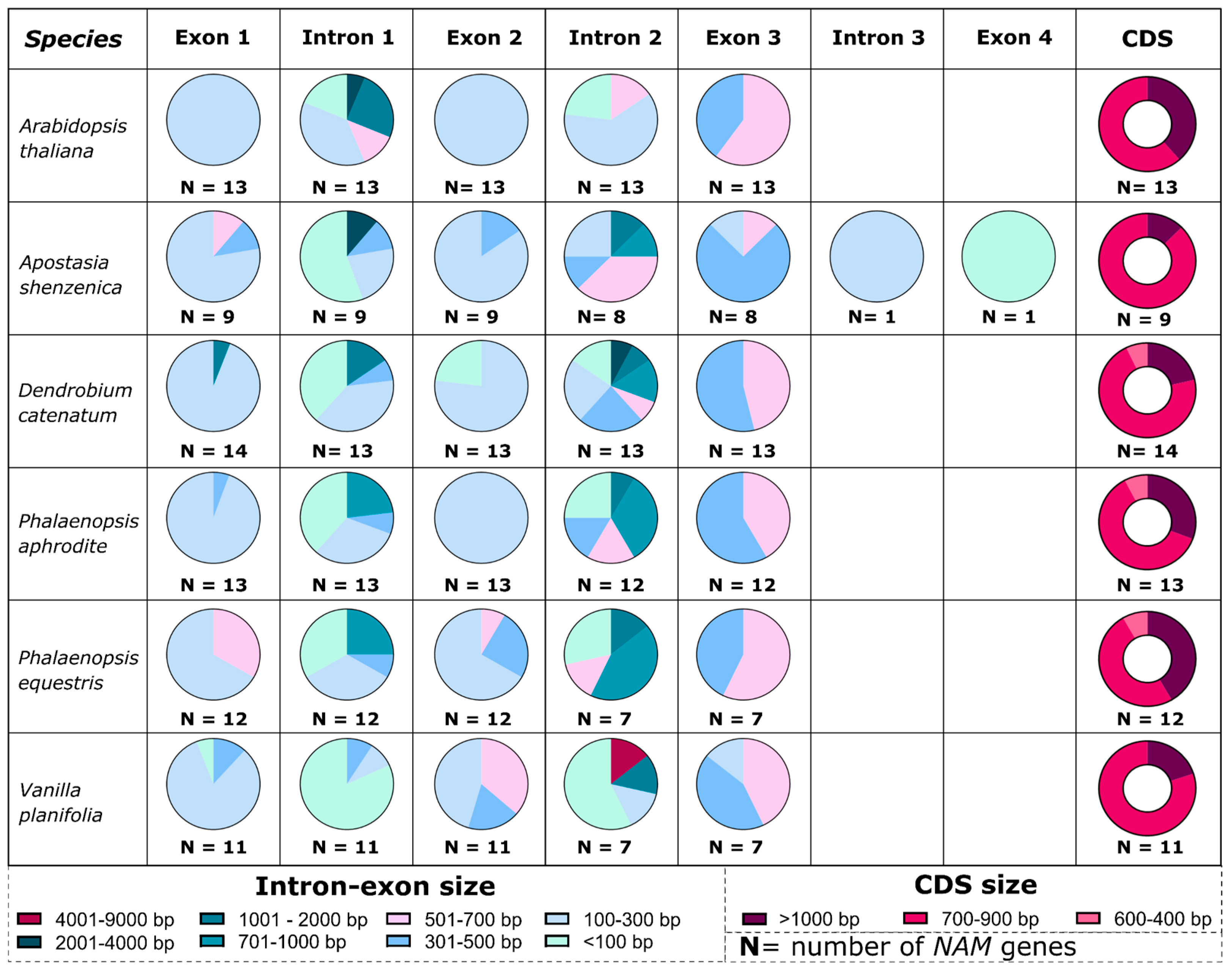
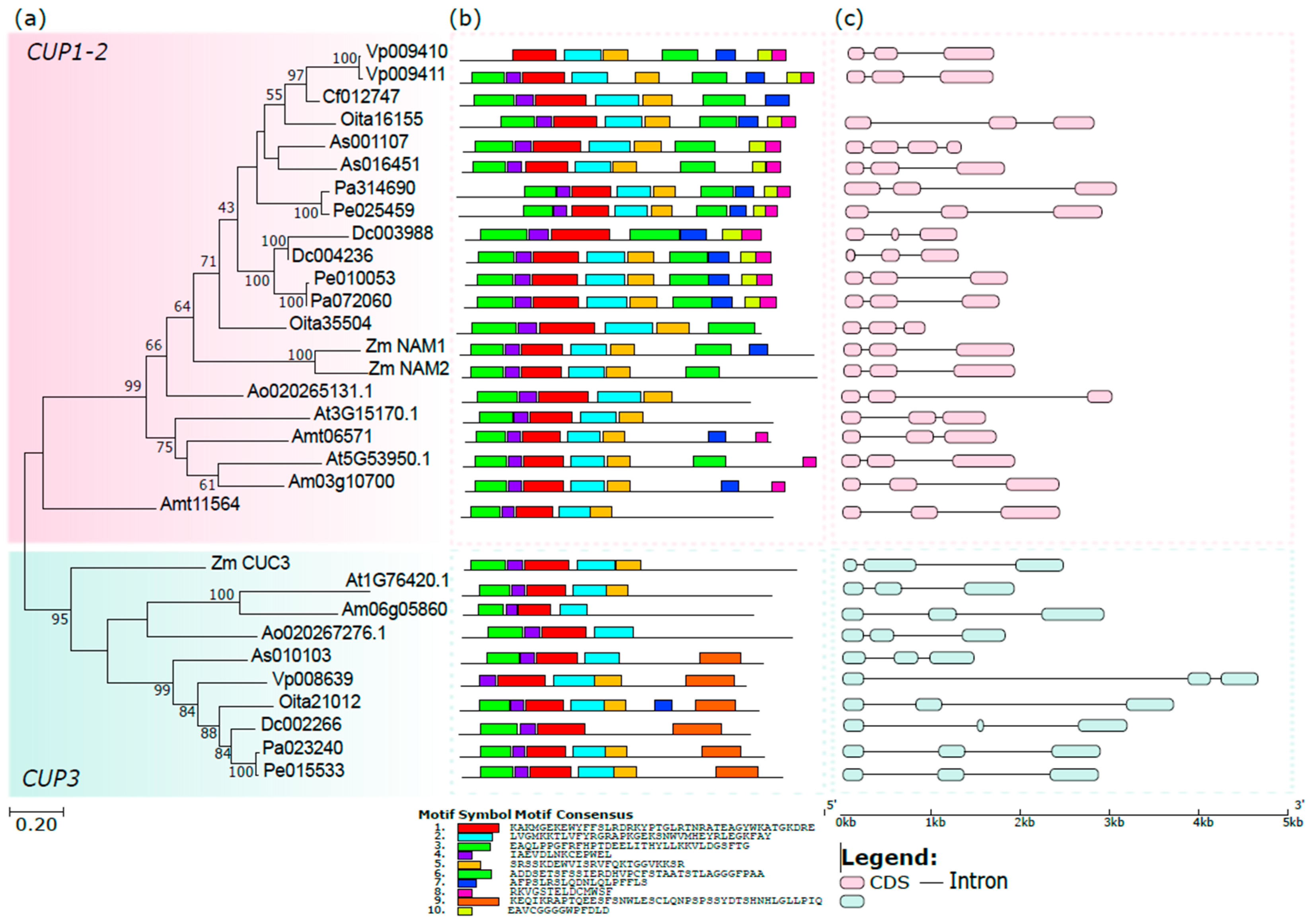
3.2. The Orchid CUP Genes: Phylogeny, Structure, and Expression Pattern
3.3. Expression Pattern of miR164
3.4. PaCUP1–PaCUP2 Protein Interaction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pérez-Rodríguez, P.; Riaño-Pachón, D.M.; Corrêa, L.G.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: Updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2010, 38, D822–D827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sablowski, R.W.; Meyerowitz, E.M. A Homolog of NO APICAL MERISTEM Is an Immediate Target of the Floral Homeotic Genes APETALA3/PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The No Apical Meristem Gene of Petunia Is Required for Pattern Formation in Embryos and Flowers and Is Expressed at Meristem and Primordia Boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Takada, S.; Hibara, K.; Ishida, T.; Tasaka, M. The CUP-SHAPED COTYLEDON1 gene of Arabidopsis regulates shoot apical meristem formation. Development 2001, 128, 1127–1135. [Google Scholar] [CrossRef]
- Vroemen, C.W.; Mordhorst, A.P.; Albrecht, C.; Kwaaitaal, M.A.C.J.; de Vries, S.C. The CUP-SHAPED COTYLEDON3 Gene Is Required for Boundary and Shoot Meristem Formation in Arabidopsis. Plant Cell 2003, 15, 1563–1577. [Google Scholar] [CrossRef]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.-H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weir, I.; Lu, J.; Cook, H.; Causier, B.; Schwarz-Sommer, Z.; Davies, B. CUPULIFORMIS establishes lateral organ boundaries in Antirrhinum. Development 2004, 131, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Collinge, M.; Boller, T. Differential induction of two potato genes, Stprx2 and StNAC, in response to infection by Phytophthora infestans and to wounding. Plant Mol. Biol. 2001, 46, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Qu, F.; Morris, T.J. HRT Gene Function Requires Interaction between a NAC Protein and Viral Capsid Protein to Confer Resistance to Turnip Crinkle Virus. Plant Cell 2000, 12, 1917–1926. [Google Scholar] [CrossRef]
- Xie, Q.; Sanz-Burgos, A.P.; Guo, H.; García, J.A.; Gutiérrez, C. GRAB proteins, novel members of the NAC domain family, isolated by their interaction with a geminivirus protein. Plant Mol. Biol. 1999, 39, 647–656. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.-S.P.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef]
- Hegedus, D.; Yu, M.; Baldwin, D.; Gruber, M.; Sharpe, A.; Parkin, I.; Whitwill, S.; Lydiate, D. Molecular characterization of Brassica napus NAC domain transcriptional activators induced in response to biotic and abiotic stress. Plant Mol. Biol. 2003, 53, 383–397. [Google Scholar] [CrossRef]
- Tran, L.-S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and Functional Analysis of Arabidopsis Stress-Inducible NAC Transcription Factors That Bind to a Drought-Responsive cis-Element in the early responsive to dehydration stress 1 Promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebocho, A.B.; Kennaway, J.R.; Bangham, J.A.; Coen, E. Formation and Shaping of the Antirrhinum Flower through Modulation of the CUP Boundary Gene. Curr. Biol. 2017, 27, 2610–2622.e3. [Google Scholar] [CrossRef] [Green Version]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive Analysis of NAC Family Genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Rushton, P.J.; Bokowiec, M.T.; Han, S.; Zhang, H.; Brannock, J.F.; Chen, X.; Laudeman, T.W.; Timko, M.P. Tobacco Transcription Factors: Novel Insights into Transcriptional Regulation in the Solanaceae. Plant Physiol. 2008, 147, 280–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, M.; Hsieh, T.-F.; Kim, S.Y.; Thomas, T.L. Molecular characterization of AtNAM: A member of the Arabidopsis NAC domain superfamily. Plant Mol. Biol. 2002, 50, 237–248. [Google Scholar] [CrossRef]
- Ernst, H.A.; Olsen, A.N.; Larsen, S.; Leggio, L.L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Johansson, E.; Larsen, S.; Skriver, K. Preliminary crystallographic analysis of the NAC domain of ANAC, a member of the plant-specific NAC transcription factor family. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 112–115. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.E.; Barley, R.; Curtis, M.; Arroyo, J.M.; Dunham, M.; Hudson, A.; Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis. Nature 2000, 408, 967–971. [Google Scholar] [CrossRef] [Green Version]
- Daimon, Y.; Takabe, K.; Tasaka, M. The CUP-SHAPED COTYLEDON genes promote adventitious shoot formation on calli. Plant Cell Physiol. 2003, 44, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibara, K.-I.; Takada, S.; Tasaka, M. CUC1 gene activates the expression of SAM-related genes to induce adventitious shoot formation. Plant J. 2003, 36, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Luo, D.; Carpenter, R.; Copsey, L.; Vincent, C.; Clark, J.; Coen, E. Control of Organ Asymmetry in Flowers of Antirrhinum. Cell 1999, 99, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam, H.; Marguerettaz, M.; Qadri, R.; Adroher, B.; Richaud, F.; Collin, M.; Thuillet, A.-C.; Vigouroux, Y.; Laufs, P.; Tregear, J.W.; et al. Divergent Expression Patterns of miR164 and CUP-SHAPED COTYLEDON Genes in Palms and Other Monocots: Implication for the Evolution of Meristem Function in Angiosperms. Mol. Biol. Evol. 2011, 28, 1439–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laufs, P.; Peaucelle, A.; Morin, H.; Traas, J. MicroRNA regulation of the CUC genes is required for boundary size control in Arabidopsis meristems. Development 2004, 131, 4311–4322. [Google Scholar] [CrossRef] [Green Version]
- Mallory, A.C.; Dugas, D.; Bartel, D.P.; Bartel, B. MicroRNA Regulation of NAC-Domain Targets Is Required for Proper Formation and Separation of Adjacent Embryonic, Vegetative, and Floral Organs. Curr. Biol. 2004, 14, 1035–1046. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.-W.; Tian, X.-H.; Ma, G.-H.; Li, Z.-X. Isolation and Functional Analysis of ZmLTP3, a Homologue to Arabidopsis LTP. Int. J. Mol. Sci. 2013, 14, 5025–5035. [Google Scholar] [CrossRef]
- Larue, C.T.; Wen, J.; Walker, J.C. A microRNA-transcription factor module regulates lateral organ size and patterning in Arabidopsis. Plant J. 2009, 58, 450–463. [Google Scholar] [CrossRef]
- Aceto, S.; Gaudio, L. The MADS and the Beauty: Genes Involved in the Development of Orchid Flowers. Curr. Genom. 2011, 12, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, S.; Widmer, A. Orchid diversity: An evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Rudall, P.J.; Bateman, R.M. Roles of synorganisation, zygomorphy and heterotopy in floral evolution: The gynostemium and labellum of orchids and other lilioid monocots. Biol. Rev. 2002, 77, 403–441. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Liu, X.; Vanneste, K.; Proost, S.; Tsai, W.-C.; Liu, K.-W.; Chen, L.-J.; He, Y.; Xu, Q.; Bian, C.; et al. The genome sequence of the orchid Phalaenopsis equestris. Nat. Genet. 2015, 47, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirks-Mulder, A.; Butôt, R.; van Schaik, P.; Wijnands, J.W.P.M.; Berg, R.V.D.; Krol, L.; Doebar, S.; van Kooperen, K.; de Boer, H.; Kramer, E.M.; et al. Exploring the evolutionary origin of floral organs of Erycina pusilla, an emerging orchid model system. BMC Evol. Biol. 2017, 17, 89. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.-F.; Hsu, W.-H.; Lee, Y.-I.; Mao, W.-T.; Yang, J.-Y.; Li, J.-Y.; Yang, C.-H. Model for perianth formation in orchids. Nat. Plants 2015, 1, 15046. [Google Scholar] [CrossRef]
- Mondragón-Palomino, M.; Theißen, G. Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes. Ann. Bot. 2009, 104, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Mondragón-Palomino, M.; Theißen, G. Conserved differential expression of paralogous DEFICIENS- and GLOBOSA-like MADS-box genes in the flowers of Orchidaceae: Refining the ‘orchid code’. Plant J. 2011, 66, 1008–1019. [Google Scholar] [CrossRef]
- Pan, Z.-J.; Cheng, C.-C.; Tsai, W.-C.; Chung, M.-C.; Chen, W.-H.; Hu, J.-M.; Chen, H.-H. The Duplicated B-class MADS-Box Genes Display Dualistic Characters in Orchid Floral Organ Identity and Growth. Plant Cell Physiol. 2011, 52, 1515–1531. [Google Scholar] [CrossRef]
- Valoroso, M.C.; Censullo, M.C.; Aceto, S. The MADS-box genes expressed in the inflorescence of Orchis italica (Orchidaceae). PLoS ONE 2019, 14, e0213185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.-Q.; Liu, K.-W.; Li, Z.; Lohaus, R.; Hsiao, Y.-Y.; Niu, S.-C.; Wang, J.-Y.; Lin, Y.-C.; Xu, Q.; Chen, L.-J.; et al. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.-Q.; Xu, Q.; Bian, C.; Tsai, W.-C.; Yeh, C.-M.; Liu, K.-W.; Yoshida, K.; Zhang, L.-S.; Chang, S.-B.; Chen, F.; et al. The Dendrobium catenatum Lindl. genome sequence provides insights into polysaccharide synthase, floral development and adaptive evolution. Sci. Rep. 2016, 6, 19029. [Google Scholar] [CrossRef] [Green Version]
- Valoroso, M.C.; De Paolo, S.; Iazzetti, G.; Aceto, S. Transcriptome-Wide Identification and Expression Analysis of DIVARICATA- and RADIALIS-Like Genes of the Mediterranean Orchid Orchis italica. Genome Biol. Evol. 2017, 9, 1418–1431. [Google Scholar] [CrossRef] [Green Version]
- Valoroso, M.C.; Sobral, R.; Saccone, G.; Salvemini, M.; Costa, M.M.R.; Aceto, S. Evolutionary Conservation of the Orchid MYB Transcription Factors DIV, RAD, and DRIF. Front. Plant Sci. 2019, 10, 1359. [Google Scholar] [CrossRef]
- De Paolo, S.; Gaudio, L.; Aceto, S. Analysis of the TCP genes expressed in the inflorescence of the orchid Orchis italica. Sci. Rep. 2015, 5, 16265. [Google Scholar] [CrossRef] [Green Version]
- Lucibelli, F.; Valoroso, M.C.; Theißen, G.; Nolden, S.; Mondragon-Palomino, M.; Aceto, S. Extending the Toolkit for Beauty: Differential Co-Expression of DROOPING LEAF-Like and Class B MADS-Box Genes during Phalaenopsis Flower Development. Int. J. Mol. Sci. 2021, 22, 7025. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-T.; Chen, W.-C.; Chen, C.-Y.; Ho, H.-Y.; Yeh, C.-H.; Kuo, Y.-T.; Su, C.-L.; Yen, S.-H.; Hsueh, H.-Y.; Yeh, J.-H.; et al. Chromosome-level assembly, genetic and physical mapping of Phalaenopsis aphrodite genome provides new insights into species adaptation and resources for orchid breeding. Plant Biotechnol. J. 2018, 16, 2027–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paolo, S.; Salvemini, M.; Gaudio, L.; Aceto, S. De Novo Transcriptome Assembly from Inflorescence of Orchis italica: Analysis of Coding and Non-Coding Transcripts. PLoS ONE 2014, 9, e102155. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Resende, M.F.R., Jr.; Bombarely, A.; Brym, M.; Bassil, E.; Chambers, A.H. Genomics-based diversity analysis of Vanilla species using a Vanilla planifolia draft genome and Genotyping-By-Sequencing. Sci. Rep. 2019, 9, 3416. [Google Scholar] [CrossRef]
- Chao, Y.-T.; Yen, S.-H.; Yeh, J.-H.; Chen, W.-C.; Shih, M.-C. Orchidstra 2.0—A Transcriptomics Resource for the Orchid Family. Plant Cell Physiol. 2017, 58, e9. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.-C.; Fu, C.-H.; Hsiao, Y.-Y.; Huang, Y.-M.; Chen, L.-J.; Wang, M.; Liu, Z.-J.; Chen, H.-H. OrchidBase 2.0: Comprehensive Collection of Orchidaceae Floral Transcriptomes. Plant Cell Physiol. 2013, 54, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bel, M.; Diels, T.; Vancaester, E.; Kreft, Ł.; Botzki, A.; Van De Peer, Y.; Coppens, F.; Vandepoele, K. PLAZA 4.0: An integrative resource for functional, evolutionary and comparative plant genomics. Nucleic Acids Res. 2018, 46, D1190–D1196. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohany, O.; Gentles, A.J.; Hankus, L.; Jurka, J. Annotation, submission and screening of repetitive elements in Repbase: RepbaseSubmitter and Censor. BMC Bioinform. 2006, 7, 474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J. Isolation of plant DNA from fresh tissue. Focus 1999, 12, 13–15. [Google Scholar]
- Acri-Nunes-Miranda, R.; Mondragón-Palomino, M. Expression of paralogous SEP-, FUL-, AG- and STK-like MADS-box genes in wild-type and peloric Phalaenopsis flowers. Front. Plant Sci. 2014, 5, 76. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [Green Version]
- Gietz, R.D.; Schiestl, R.H.; Willems, A.R.; Woods, R.A. Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 1995, 11, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Wang, M.; Miao, Y.; Ni, M.; Bibi, N.; Yuan, S.; Li, F.; Wang, X. Molecular Evolution and Expansion Analysis of the NAC Transcription Factor in Zea mays. PLoS ONE 2014, 9, e111837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuchi, K.; Ueguchi-Tanaka, M.; Yoshida, K.T.; Nagato, Y.; Matsusoka, M.; Hirano, H.Y. Molecular analysis of the NAC gene family in rice. Mol. Genet. Genom. 2000, 262, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Albert, V.A.; Carretero-Paulet, L. A genome to unveil the mysteries of orchids. Nat. Genet. 2015, 47, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lan, S.; Yin, W.-L.; Liu, Z.-J. Genome-Wide Identification and Expression Pattern Analysis of KNOX Gene Family in Orchidaceae. Front. Plant Sci. 2022, 13, 901089. [Google Scholar] [CrossRef] [PubMed]
- Salemme, M.; Sica, M.; Gaudio, L.; Aceto, S. The OitaAG and OitaSTK genes of the orchid Orchis italica: A comparative analysis with other C- and D-class MADS-box genes. Mol. Biol. Rep. 2013, 40, 3523–3535. [Google Scholar] [CrossRef]
- Salemme, M.; Sica, M.; Iazzetti, G.; Gaudio, L.; Aceto, S. The AP2-Like Gene OitaAP2 Is Alternatively Spliced and Differentially Expressed in Inflorescence and Vegetative Tissues of the Orchid Orchis italica. PLoS ONE 2013, 8, e77454. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, R.; Werr, W. Pattern Formation in the Monocot Embryo as Revealed by NAMand CUC3 Orthologues from Zea mays L. Plant Mol. Biol. 2005, 58, 669–685. [Google Scholar] [CrossRef] [PubMed]
- Kamiuchi, Y.; Yamamoto, K.; Furutani, M.; Tasaka, M.; Aida, M. The CUC1 and CUC2 genes promote carpel margin meristem formation during Arabidopsis gynoecium development. Front. Plant Sci. 2014, 5, 165. [Google Scholar] [CrossRef] [Green Version]
- Blein, T.; Pulido, A.; Vialette-Guiraud, A.; Nikovics, K.; Morin, H.; Hay, A.; Johansen, I.E.; Tsiantis, M.; Laufs, P. A Conserved Molecular Framework for Compound Leaf Development. Science 2008, 322, 1835–1839. [Google Scholar] [CrossRef] [Green Version]
- Vialette-Guiraud, A.C.M.; Adam, H.; Finet, C.; Jasinski, S.; Jouannic, S.; Scutt, C.P. Insights from ANA-grade angiosperms into the early evolution of CUP-SHAPED COTYLEDON genes. Ann. Bot. 2011, 107, 1511–1519. [Google Scholar] [CrossRef]
- Maugarny, A.; Gonçalves, B.; Arnaud, N.; Laufs, P. Chapter 15—CUC Transcription Factors: To the Meristem and Beyond. In Plant Transcription Factors; Academic Press: Cambridge, MA, USA, 2016; pp. 229–247. [Google Scholar] [CrossRef]
- Hasson, A.; Plessis, A.; Blein, T.; Adroher, B.; Grigg, S.; Tsiantis, M.; Boudaoud, A.; Damerval, C.; Laufs, P. Evolution and Diverse Roles of the CUP-SHAPED COTYLEDON Genes in Arabidopsis Leaf Development. Plant Cell 2011, 23, 54–68. [Google Scholar] [CrossRef] [Green Version]
- Citerne, H.; Jabbour, F.; Nadot, S.; Damerval, C. The Evolution of Floral Symmetry. Adv. Bot. Res. 2010, 10, 85–137. [Google Scholar] [CrossRef]
- Corley, S.B.; Carpenter, R.; Copsey, L.; Coen, E. Floral asymmetry involves an interplay between TCP and MYB transcription factors in Antirrhinum. Proc. Natl. Acad. Sci. USA 2005, 102, 5068–5073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimundo, J.; Sobral, R.; Bailey, P.; Azevedo, H.; Galego, L.; Almeida, J.; Coen, E.; Costa, M.M.R. A subcellular tug of war involving three MYB-like proteins underlies a molecular antagonism in Antirrhinum flower asymmetry. Plant J. 2013, 75, 527–538. [Google Scholar] [CrossRef]
- Baker, C.C.; Sieber, P.; Wellmer, F.; Meyerowitz, E.M. The early extra petals1 Mutant Uncovers a Role for MicroRNA miR164c in Regulating Petal Number in Arabidopsis. Curr. Biol. 2005, 15, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikovics, K.; Blein, T.; Peaucelle, A.; Ishida, T.; Morin, H.; Aida, M.; Laufs, P. The Balance between the MIR164A and CUC2 Genes Controls Leaf Margin Serration in Arabidopsis. Plant Cell 2006, 18, 2929–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, B.; Hasson, A.; Belcram, K.; Cortizo, M.; Morin, H.; Nikovics, K.; Vialette-Guiraud, A.; Takeda, S.; Aida, M.; Laufs, P.; et al. A conserved role for CUP-SHAPED COTYLEDON genes during ovule development. Plant J. 2015, 83, 732–742. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application | Gene | Forward (5’-3’) | Reverse (5’-3’) |
|---|---|---|---|
| gDNA | Oita16155 | TCCTCCTCCTGACAACCCCT | GCTGTAGATCTCCCGGTCTT |
| Oita16155 | ATGAGTACAGGTTGGAGGGC | GCACCGCTTTCCTTCATCTTC | |
| Oita35504 | CCCTCACTCTTCTGGCATGG | GGCATGTGATCCGTCTCGAT | |
| Oita21012 | ACGTCACCTGTCTTGTTCGG | TTGCGGTCTCTTAGGCTGAA | |
| Oita21012 | AGACTTCTCCTTCCGACACG | AGGTGTCTGCAGTTTGGTGG | |
| Oita21012 | GCTGTAGATCTCCCGGTCTT | ||
| cDNA | Pa_CUP1 | ACAAATCCCATACGCTTCGC | AACATGCAGTCCAACTCCGT |
| Pa_CUP2 | CTTCTAACTATCCCGGCCCC | ATCTCACACTCTGTCACCGG | |
| Pa_CUP3 | GGCTTGAATCCTGCTTACAA | GGTGCTCCTCCTTGGATTGG | |
| qPCR | Pa_CUP1 | ACAAATCCCATACGCTTCGC | AACATGCAGTCCAACTCCGT |
| Pa_CUP2 | CTTCTAACTATCCCGGCCCC | ATCTCACACTCTGTCACCGG | |
| Pa_CUP3 | GGCTTGAATCCTGCTTACAA | GGTGCTCCTCCTTGGATTGG | |
| Pa18S | TTAGGCCACGGAAGTTTGAG | ACACTTCACCGGACCATTCAA | |
| miRNA | miRNA164_Stloop | GCGGCGTGGAGAAGCAGG | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCGCACGTGC |
| Stloop_18S | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCACTTC | ||
| Stloop_UNIV | GTGCAGGGTCCGAGGT | ||
| Y2H | Pa_CUP1 | AGGATCCAAATGGAGAACTTCAGCCACCAT | AACTGCAGGATGATGTCATCCACTTAAAATCGT |
| Pa_CUP2 | AGGATCCAAATGGAGAACTTCAGTCAGCATTTCGAC | AACTGCAGTGGAAGCACAAAACATAATTAGCTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valoroso, M.C.; Lucibelli, F.; Aceto, S. Orchid NAC Transcription Factors: A Focused Analysis of CUPULIFORMIS Genes. Genes 2022, 13, 2293. https://doi.org/10.3390/genes13122293
Valoroso MC, Lucibelli F, Aceto S. Orchid NAC Transcription Factors: A Focused Analysis of CUPULIFORMIS Genes. Genes. 2022; 13(12):2293. https://doi.org/10.3390/genes13122293
Chicago/Turabian StyleValoroso, Maria Carmen, Francesca Lucibelli, and Serena Aceto. 2022. "Orchid NAC Transcription Factors: A Focused Analysis of CUPULIFORMIS Genes" Genes 13, no. 12: 2293. https://doi.org/10.3390/genes13122293