

Oridonin Protects against Myocardial Ischemia–Reperfusion Injury by Inhibiting GSDMD-Mediated Pyroptosis
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and H/R Treatment of Neonatal Rat Cardiomyocytes (NRCMs)
2.2. Mice and I/R Treatment
2.3. Evans Blue/TTC Staining
2.4. CCK-8 Assay
2.5. Western Blot Analysis
2.6. Hematoxylin and Eosin (H&E) Staining
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Immunohistochemistry
2.9. TdT-Mediated DUTP Nick End Labeling (TUNEL) Assay
2.10. Immunofluorescence
2.11. Statistical Analysis
3. Results
3.1. Oridonin Reduced Infarct Size and Improved the Morphology of Cardiomyocytes
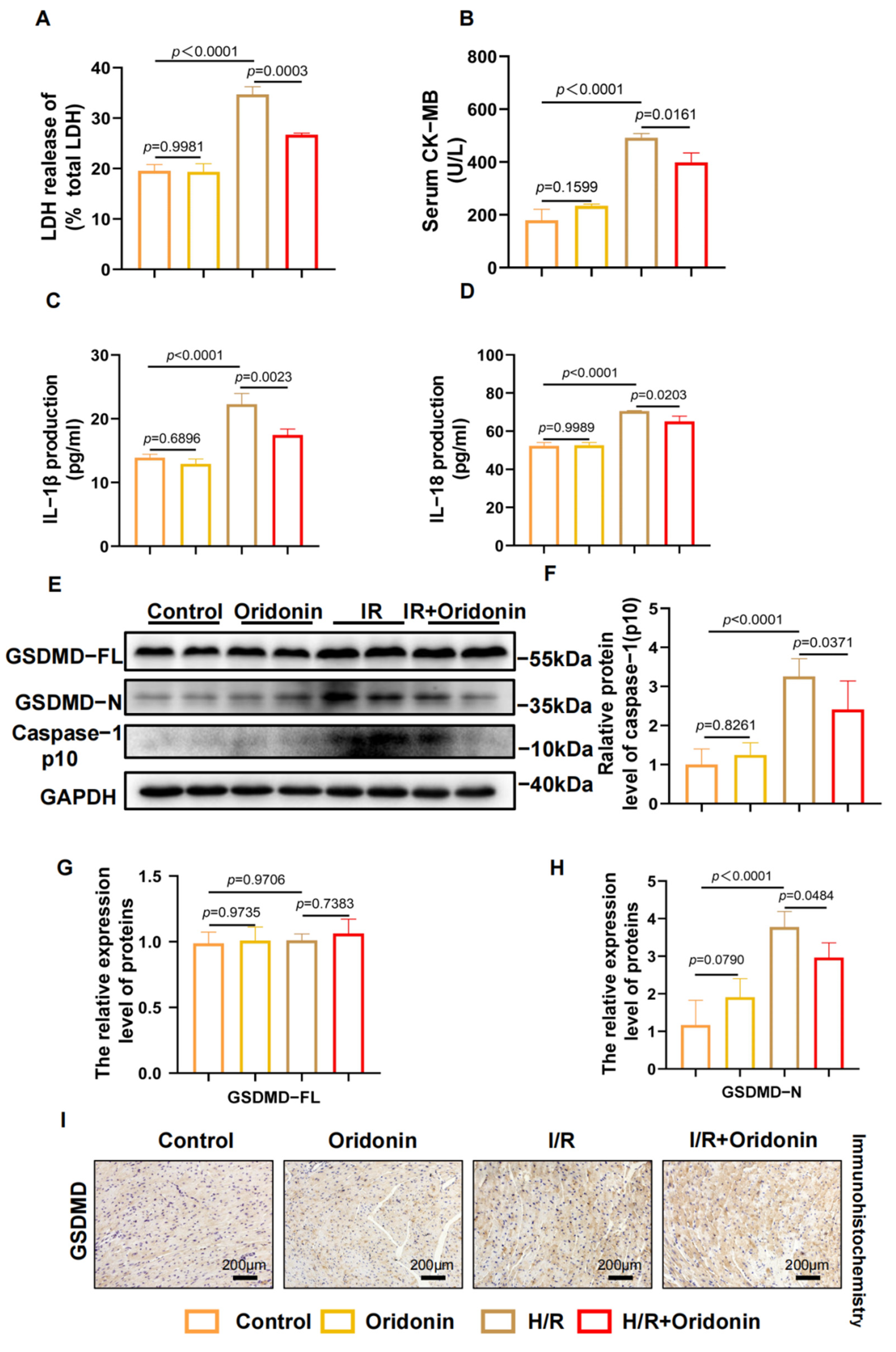
3.2. Oridonin Reduces IR-Induced Myocardial Pyroptosis In Vivo
3.3. Oridonin Protected Cardiomyocytes from H/R-Induced Pyroptosis
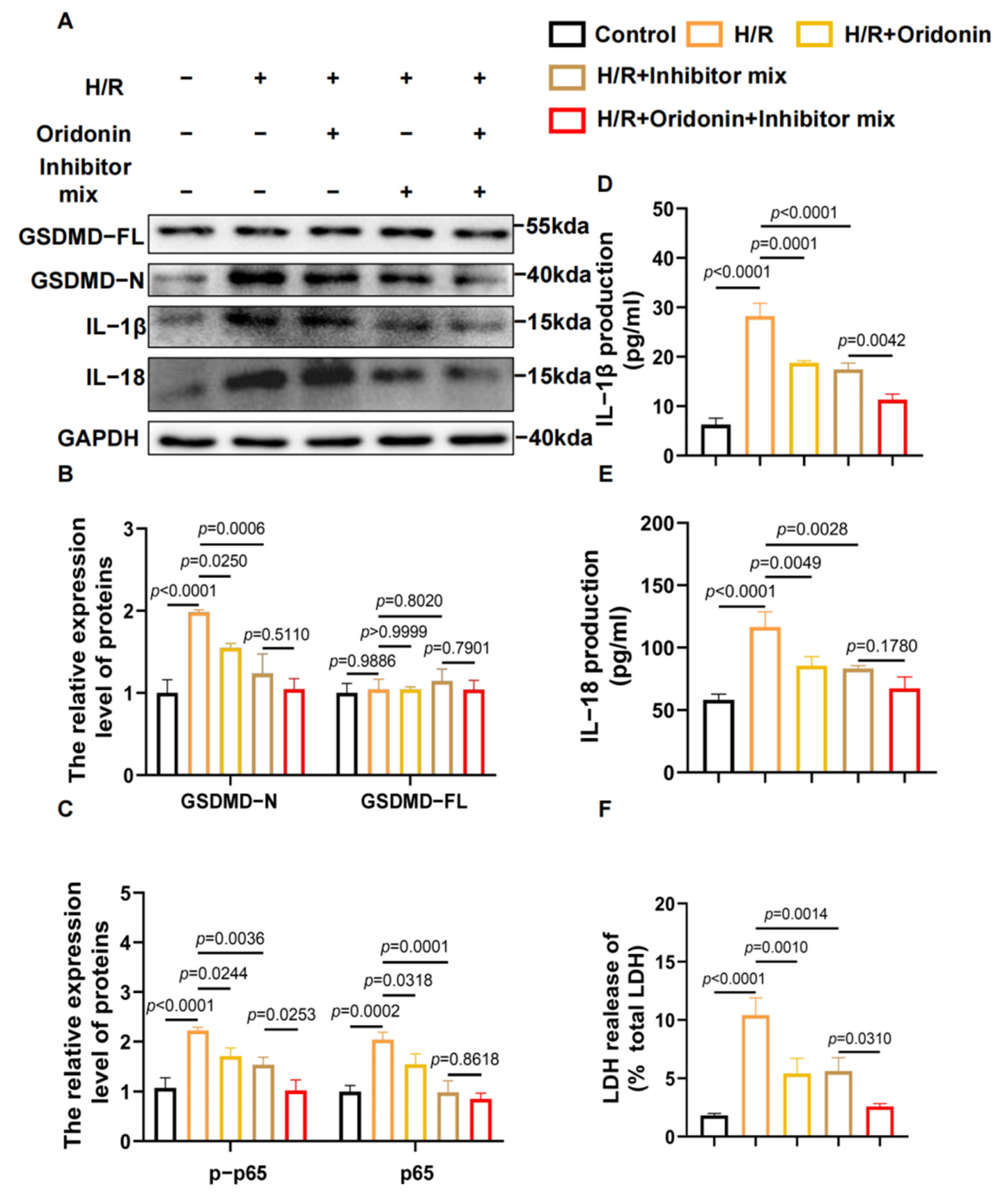
3.4. Oridonin Regulates GSDMD-Mediated Pyroptosis via the NF-κB/NLRP3 Signaling Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| H/R | hypoxia/reoxygenation |
| I/R | ischemia/reperfusion |
| NRCMs | neonatal rat cardiomyocytes |
| CCK | cell counting kit |
| TUNEL | terminal-deoxynucleotidyl transferase-mediated nick end labeling |
| INF | infarct size |
| AAR | area at risk |
| LV | left ventricle |
| Ori | oridonin |
| LDH | lactate dehydrogenase |
| CK-MB | creatine kinase isoenzyme |
| GSDMD-FL | gasdermin D-full length |
| GSDMD-N | gasdermin D-N domain |
References
- Mechanic, O.J.; Gavin, M.; Grossman, S.A. Acute Myocardial Infarction; StatPearls: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29083808 (accessed on 15 March 2022).
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. Available online: https://www.ncbi.nlm.nih.gov/pubmed/23281415 (accessed on 15 March 2022). [CrossRef] [PubMed] [Green Version]
- Ibanez, B.; Heusch, G.; Ovize, M.; Van de Werf, F. Evolving therapies for myocardial ischemia/reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1454–1471. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25857912 (accessed on 15 March 2022). [CrossRef] [PubMed] [Green Version]
- Wallert, M.; Ziegler, M.; Wang, X.; Maluenda, A.; Xu, X.; Yap, M.L.; Witt, R.; Giles, C.; Kluge, S.; Hortmann, M.; et al. alpha-Tocopherol preserves cardiac function by reducing oxidative stress and inflammation in ischemia/reperfusion injury. Redox Biol. 2019, 26, 101292. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31419755 (accessed on 15 March 2022). [CrossRef] [PubMed]
- Mezzaroma, E.; Toldo, S.; Farkas, D.; Seropian, I.M.; Van Tassell, B.W.; Salloum, F.N.; Kannan, H.R.; Menna, A.C.; Voelkel, N.F.; Abbate, A. The inflammasome promotes adverse cardiac remodeling following acute myocardial infarction in the mouse. Proc. Natl. Acad. Sci. USA 2011, 108, 19725–19730. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22106299 (accessed on 15 March 2022). [CrossRef] [Green Version]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26375003 (accessed on 15 March 2022). [CrossRef]
- Zhang, D.; Qian, J.; Zhang, P.; Li, H.; Shen, H.; Li, X.; Chen, G. Gasdermin D serves as a key executioner of pyroptosis in experimental cerebral ischemia and reperfusion model both in vivo and in vitro. J. Neurosci. Res. 2019, 97, 645–660. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30600840 (accessed on 15 March 2022). [CrossRef]
- Prabhu, S.D.; Frangogiannis, N.G. The Biological Basis for Cardiac Repair After Myocardial Infarction: From Inflammation to Fibrosis. Circ. Res. 2016, 119, 91–112. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27340270 (accessed on 15 March 2022). [CrossRef]
- Lu, C.; Chen, C.; Chen, A.; Wu, Y.; Wen, J.; Huang, F.; Zeng, Z. Oridonin Attenuates Myocardial Ischemia/Reperfusion Injury via Downregulating Oxidative Stress and NLRP3 Inflammasome Pathway in Mice. Evid. Based Complement. Altern. Med. 2020, 2020, 7395187. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32565873 (accessed on 15 March 2022). [CrossRef]
- Li, X.; Zhang, C.T.; Ma, W.; Xie, X.; Huang, Q. Oridonin: A Review of Its Pharmacology, Pharmacokinetics and Toxicity. Front. Pharmacol. 2021, 12, 645824. Available online: https://www.ncbi.nlm.nih.gov/pubmed/34295243 (accessed on 15 March 2022). [CrossRef]
- Gao, R.F.; Li, X.; Xiang, H.Y.; Yang, H.; Lv, C.Y.; Sun, X.L.; Chen, H.Z.; Gao, Y.; Yang, J.S.; Luo, W.; et al. The covalent NLRP3-inflammasome inhibitor oridonin relieves myocardial infarction induced myocardial fibrosis and cardiac remodeling in mice. Int. Immunopharmacol. 2021, 90, 107133. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33168408 (accessed on 15 March 2022). [CrossRef]
- Wang, S.; Yang, H.; Yu, L.; Jin, J.; Qian, L.; Zhao, H.; Xu, Y.; Zhu, X. Oridonin attenuates Abeta1-42-induced neuroinflammation and inhibits NF-kappaB pathway. PLoS ONE 2014, 9, e104745. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25121593 (accessed on 15 March 2022).
- Liu, D.; Qin, H.; Yang, B.; Du, B.; Yun, X. Oridonin ameliorates carbon tetrachloride-induced liver fibrosis in mice through inhibition of the NLRP3 inflammasome. Drug Dev. Res. 2020, 81, 526–533. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32219880 (accessed on 15 March 2022). [CrossRef] [PubMed] [Green Version]
- Shao, B.Z.; Xu, Z.Q.; Han, B.Z.; Su, D.F.; Liu, C. NLRP3 inflammasome and its inhibitors: A review. Front Pharmacol. 2015, 6, 262. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26594174 (accessed on 15 March 2022). [CrossRef] [Green Version]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-kappaB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30464573 (accessed on 15 March 2022). [CrossRef] [PubMed] [Green Version]
- Guo, X.; Hong, S.; He, H.; Zeng, Y.; Chen, Y.; Mo, X.; Li, J.; Li, L.; Steinmetz, R.; Liu, Q. NFkappaB promotes oxidative stress-induced necrosis and ischemia/reperfusion injury by inhibiting Nrf2-ARE pathway. Free Radic. Biol. Med. 2020, 159, 125–135. Available online: https://www.ncbi.nlm.nih.gov/pubmed/32745764 (accessed on 15 March 2022). [CrossRef]
- Nennig, S.E.; Schank, J.R. The Role of NFkB in Drug Addiction: Beyond Inflammation. Alcohol. Alcohol. 2017, 52, 172–179. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28043969 (accessed on 15 March 2022). [CrossRef]
- He, H.; Jiang, H.; Chen, Y.; Ye, J.; Wang, A.; Wang, C.; Liu, Q.; Liang, G.; Deng, X.; Jiang, W.; et al. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat. Commun. 2018, 9, 2550. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29959312 (accessed on 15 March 2022). [CrossRef] [Green Version]
- Triantafilou, K. Enigmatic inflammasomes. Immunology 2021, 162, 249–251. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33590488 (accessed on 15 March 2022). [CrossRef]
- He, Y.; Hara, H.; Nunez, G. Mechanism and Regulation of NLRP3 Inflammasome Activation. Trends Biochem. Sci. 2016, 41, 1012–1021. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27669650 (accessed on 15 March 2022). [CrossRef] [Green Version]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. Available online: https://www.ncbi.nlm.nih.gov/pubmed/27383986 (accessed on 15 March 2022). [CrossRef] [Green Version]
- Man, S.M.; Kanneganti, T.D. Gasdermin D: The long-awaited executioner of pyroptosis. Cell Res. 2015, 25, 1183–1184. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26482951 (accessed on 15 March 2022). [CrossRef] [PubMed]
- Jia, Y.; Tong, Y.; Min, L.; Li, Y.; Cheng, Y. Protective effects of oridonin against cerebral ischemia/reperfusion injury by inhibiting the NLRP3 inflammasome activation. Tissue Cell 2021, 71, 101514. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33676236 (accessed on 15 March 2022). [CrossRef] [PubMed]
- Zhao, X.; Liu, Y.; Wang, L.; Yan, C.; Liu, H.; Zhang, W.; Zhao, H.; Cheng, C.; Chen, Z.; Xu, T.; et al. Oridonin attenuates hind limb ischemia-reperfusion injury by modulating Nrf2-mediated oxidative stress and NLRP3-mediated inflammation. J. Ethnopharmacol. 2022, 292, 115206. Available online: https://www.ncbi.nlm.nih.gov/pubmed/35301099 (accessed on 15 March 2022). [CrossRef] [PubMed]
- Fu, M.; Xie, D.; Sun, Y.; Pan, Y.; Zhang, Y.; Chen, X.; Shi, Y.; Deng, S.; Cheng, B. Exosomes derived from MSC pre-treated with oridonin alleviates myocardial IR injury by suppressing apoptosis via regulating autophagy activation. J. Cell. Mol. Med. 2021, 25, 5486–5496. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33955654 (accessed on 15 March 2022). [CrossRef] [PubMed]
- Tan, R.Z.; Yan, Y.; Yu, Y.; Diao, H.; Zhong, X.; Lin, X.; Liao, Y.Y.; Wang, L. Renoprotective Effect of Oridonin in a Mouse Model of Acute Kidney Injury via Suppression of Macrophage Involved Inflammation. Biol. Pharm. Bull. 2021, 44, 714–723. Available online: https://www.ncbi.nlm.nih.gov/pubmed/33952827 (accessed on 15 March 2022). [CrossRef]
- Zhang, J.; Zhou, Y.; Sun, Y.; Yan, H.; Han, W.; Wang, X.; Wang, K.; Wei, B.; Xu, X. Beneficial effects of oridonin on myocardial ischemia/reperfusion injury: Insight gained by metabolomic approaches. Eur. J. Pharmacol. 2019, 861, 172587. Available online: https://www.ncbi.nlm.nih.gov/pubmed/31377155 (accessed on 15 March 2022). [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, J.; Lai, X.; Fan, X.; Ye, B.; Zhong, L.; Zhang, Y.; Shao, R.; Shi, S.; Huang, W.; Su, L.; et al. Oridonin Protects against Myocardial Ischemia–Reperfusion Injury by Inhibiting GSDMD-Mediated Pyroptosis. Genes 2022, 13, 2133. https://doi.org/10.3390/genes13112133
Lin J, Lai X, Fan X, Ye B, Zhong L, Zhang Y, Shao R, Shi S, Huang W, Su L, et al. Oridonin Protects against Myocardial Ischemia–Reperfusion Injury by Inhibiting GSDMD-Mediated Pyroptosis. Genes. 2022; 13(11):2133. https://doi.org/10.3390/genes13112133
Chicago/Turabian StyleLin, Jiahui, Xianhui Lai, Xiaoxi Fan, Bozhi Ye, Lingfeng Zhong, Yucong Zhang, Ruiyin Shao, Si Shi, Weijian Huang, Lan Su, and et al. 2022. "Oridonin Protects against Myocardial Ischemia–Reperfusion Injury by Inhibiting GSDMD-Mediated Pyroptosis" Genes 13, no. 11: 2133. https://doi.org/10.3390/genes13112133