

Owl Monkey Alu Insertion Polymorphisms and Aotus Phylogenetics

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lineage-Specific Alu Elements
2.2. Oligonucleotide Primer Design
2.3. DNA Samples
2.4. PCR Amplification
2.5. Sanger Chain Termination DNA Sequencing
2.6. Phylogenetic Analyses
3. Results
3.1. Young Lineage-Specific Alu Elements and PCR
3.2. DNA Sequencing
3.2.1. Red-Necked Species Group (A. nancymaae and A. azarae) and Novel Alu Discovery
3.2.2. Grey-Necked Species Group (A. vociferans and A. trivirgatus)
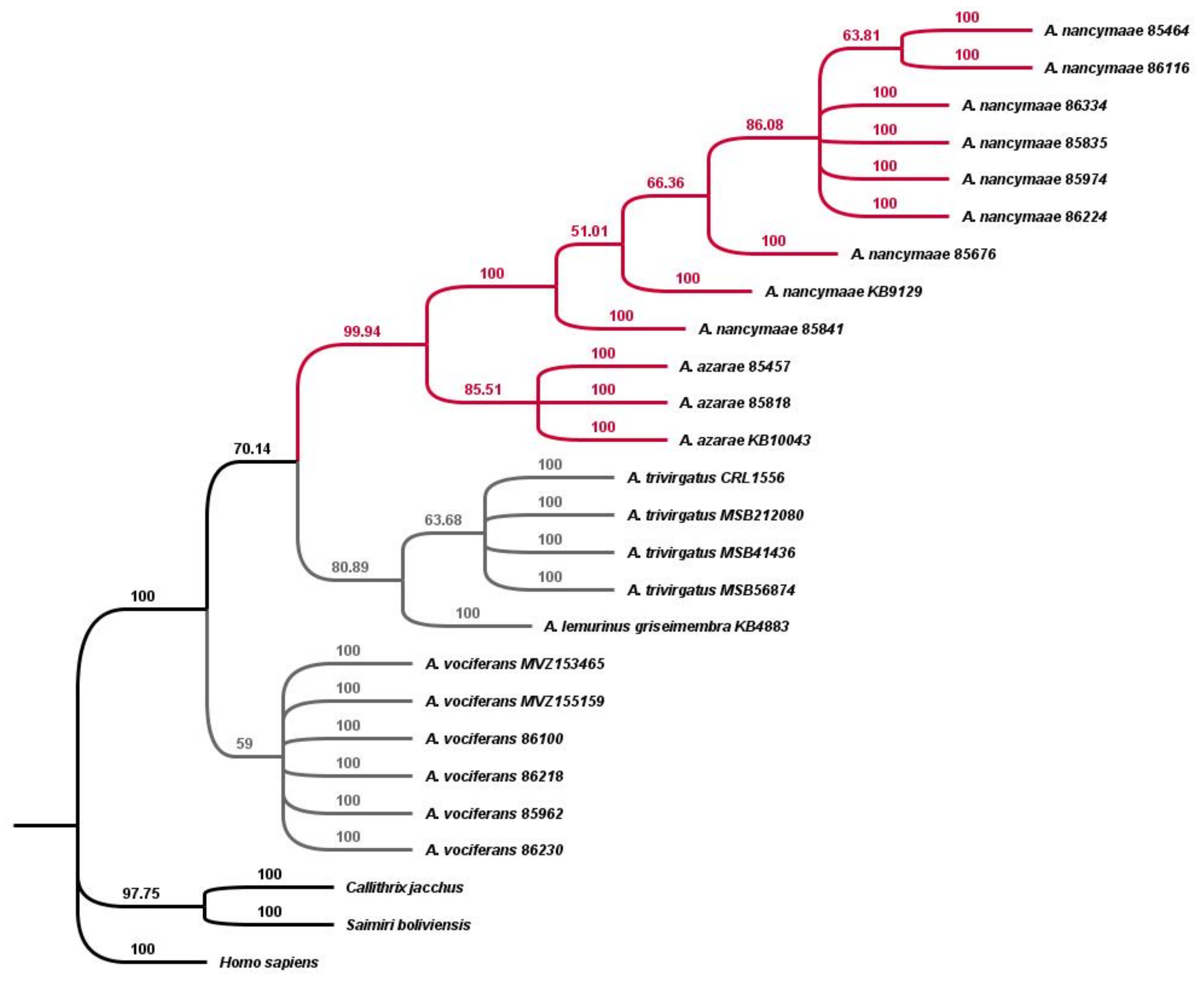
3.3. Maximum Parsimony
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins-Junior, A.M.G.; Sampaio, I.; Silva, A.; Boubli, J.; Hrbek, T.; Farias, I.; Ruiz-García, M.; Schneider, H. Out of the shadows: Multilocus systematics and biogeography of night monkeys suggest a Central Amazonian origin and a very recent widespread southeastward expansion in South America. Mol. Phylogenet. Evol. 2022, 170, 107426. [Google Scholar] [CrossRef] [PubMed]
- Menezes, A.N.; Bonvicino, C.R.; Seuánez, H.N. Identification, classification and evolution of owl monkeys (Aotus, Illiger 1811). BMC Evol. Biol. 2010, 10, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-García, M.; Vásquez, C.; Camargo, E.; Leguizamón, N.; Gálvez, H.; Vallejo, A.; Pinedo, M.; Castellanos-Mora, L.; Shostell, J.; Alvarez, D. Molecular Phylogenetics of Aotus (Platyrrhini, Cebidae). Int. J. Primatol. 2011, 32, 1218. [Google Scholar] [CrossRef]
- Hershkovitz, P. Two new species of night monkeys, genus Aotus (Cebidae, platyrrhini): A preliminary report on Aotus taxonomy. Am. J. Primatol. 1983, 4, 209–243. [Google Scholar] [CrossRef] [PubMed]
- Ashley, M.V.; Vaughn, J.L. Owl monkeys (Aotus) are highly divergent in mitochondrial Cytochromec Oxidase (COII) sequences. Int. J. Primatol. 1995, 16, 793–806. [Google Scholar] [CrossRef]
- Plautz, H.L.; Gonçalves, E.C.; Ferrari, S.F.; Schneider, M.P.; Silva, A. Evolutionary inferences on the diversity of the genus Aotus (Platyrrhini, Cebidae) from mitochondrial cytochrome c oxidase subunit II gene sequences. Mol. Phylogenet. Evol. 2009, 51, 382–387. [Google Scholar] [CrossRef]
- Perelman, P.; Johnson, W.E.; Roos, C.; Seuánez, H.N.; Horvath, J.E.; Moreira, M.A.; Kessing, B.; Pontius, J.; Roelke, M.; Rumpler, Y.; et al. A molecular phylogeny of living primates. PLoS Genet. 2011, 7, e1001342. [Google Scholar] [CrossRef] [Green Version]
- Jameson Kiesling, N.M.; Yi, S.V.; Xu, K.; Gianluca Sperone, F.; Wildman, D.E. The tempo and mode of New World monkey evolution and biogeography in the context of phylogenomic analysis. Mol. Phylogenet. Evol. 2015, 82, 386–399. [Google Scholar] [CrossRef]
- Herrera, S.; Perlaza, B.L.; Bonelo, A.; Arévalo-Herrera, M. Aotus monkeys: Their great value for anti-malaria vaccines and drug testing. Int. J. Parasitol. 2002, 32, 1625–1635. [Google Scholar] [CrossRef]
- Douglas, A.D.; Baldeviano, G.C.; Jin, J.; Miura, K.; Diouf, A.; Zenonos, Z.A.; Ventocilla, J.A.; Silk, S.E.; Marshall, J.M.; Alanine, D.G.W.; et al. A defined mechanistic correlate of protection against Plasmodium falciparum malaria in non-human primates. Nat. Commun. 2019, 10, 1953. [Google Scholar] [CrossRef]
- Ruiz-Garcia, M.; Vasques, C.; Camargo, E.; Castellanos-Mora, L.F.; Galvez, H.; Leguizamon, N.; Shostell, J.M. Molecular Genetics Analysis of mtDNA COII Gene Sequences Shows Illegal Traffic of Night Monkeys (Aotus, Platyrrhini, Primates) in Colombia. J. Primatol. 2013, 2, 107. [Google Scholar] [CrossRef]
- Deininger, P.L.; Moran, J.V.; Batzer, M.A.; Kazazian, H.H., Jr. Mobile elements and mammalian genome evolution. Curr. Opin. Genet. Dev. 2003, 13, 651–658. [Google Scholar] [CrossRef]
- Morrish, T.A.; Gilbert, N.; Myers, J.S.; Vincent, B.J.; Stamato, T.D.; Taccioli, G.E.; Batzer, M.A.; Moran, J.V. DNA repair mediated by endonuclease-independent LINE-1 retrotransposition. Nat. Genet. 2002, 31, 159–165. [Google Scholar] [CrossRef]
- Dewannieux, M.; Esnault, C.; Heidmann, T. LINE-mediated retrotransposition of marked Alu sequences. Nat. Genet. 2003, 35, 41–48. [Google Scholar] [CrossRef]
- Luan, D.D.; Korman, M.H.; Jakubczak, J.L.; Eickbush, T.H. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal target site: A mechanism for non-LTR retrotransposition. Cell 1993, 72, 595–605. [Google Scholar] [CrossRef]
- Baker, J.N.; Walker, J.A.; Denham, M.W.; Loupe, C.D., 3rd; Batzer, M.A. Recently integrated Alu insertions in the squirrel monkey (Saimiri) lineage and application for population analyses. Mob. DNA 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Han, K.; Xing, J.; Kim, H.S.; Rogers, J.; Ryder, O.A.; Disotell, T.; Yue, B.; Batzer, M.A. Phylogeny of the macaques (Cercopithecidae: Macaca) based on Alu elements. Gene 2009, 448, 242–249. [Google Scholar] [CrossRef] [Green Version]
- McLain, A.T.; Meyer, T.J.; Faulk, C.; Herke, S.W.; Oldenburg, J.M.; Bourgeois, M.G.; Abshire, C.F.; Roos, C.; Batzer, M.A. An Alu-based phylogeny of lemurs (infraorder: Lemuriformes). PLoS ONE 2012, 7, e44035. [Google Scholar] [CrossRef]
- Meyer, T.J.; McLain, A.T.; Oldenburg, J.M.; Faulk, C.; Bourgeois, M.G.; Conlin, E.M.; Mootnick, A.R.; de Jong, P.J.; Roos, C.; Carbone, L.; et al. An Alu-based phylogeny of gibbons (hylobatidae). Mol. Biol. Evol. 2012, 29, 3441–3450. [Google Scholar] [CrossRef] [Green Version]
- Perna, N.T.; Batzer, M.A.; Deininger, P.L.; Stoneking, M. Alu insertion polymorphism: A new type of marker for human population studies. Hum. Biol. 1992, 64, 641–648. [Google Scholar]
- Ray, D.A.; Batzer, M.A. Tracking Alu evolution in New World primates. BMC Evol. Biol. 2005, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.H.; Ray, D.A.; Xing, J.; Callinan, P.A.; Myers, J.S.; Hedges, D.J.; Garber, R.K.; Witherspoon, D.J.; Jorde, L.B.; Batzer, M.A. Alu elements and hominid phylogenetics. Proc. Natl. Acad. Sci. USA 2003, 100, 12787–12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steely, C.J.; Walker, J.A.; Jordan, V.E.; Beckstrom, T.O.; McDaniel, C.L.; St Romain, C.P.; Bennett, E.C.; Robichaux, A.; Clement, B.N.; Raveendran, M.; et al. Alu Insertion Polymorphisms as Evidence for Population Structure in Baboons. Genome Biol. Evol. 2017, 9, 2418–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.; Kural, D.; Stromberg, M.P.; Walker, J.A.; Konkel, M.K.; Stutz, A.M.; Urban, A.E.; Grubert, F.; Lam, H.Y.; Lee, W.P.; et al. A comprehensive map of mobile element insertion polymorphisms in humans. PLoS Genet. 2011, 7, e1002236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, J.; Wang, H.; Han, K.; Ray, D.A.; Huang, C.H.; Chemnick, L.G.; Stewart, C.B.; Disotell, T.R.; Ryder, O.A.; Batzer, M.A. A mobile element based phylogeny of Old World monkeys. Mol. Phylogenet. Evol. 2005, 37, 872–880. [Google Scholar] [CrossRef]
- Xing, J.; Wang, H.; Zhang, Y.; Ray, D.A.; Tosi, A.J.; Disotell, T.R.; Batzer, M.A. A mobile element-based evolutionary history of guenons (tribe Cercopithecini). BMC Biol. 2007, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Roos, C.; Zinner, D.; Kubatko, L.S.; Schwarz, C.; Yang, M.; Meyer, D.; Nash, S.D.; Xing, J.; Batzer, M.A.; Brameier, M.; et al. Nuclear versus mitochondrial DNA: Evidence for hybridization in colobine monkeys. BMC Evol. Biol. 2011, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Osterholz, M.; Vermeer, J.; Walter, L.; Roos, C. A PCR-based marker to simply identify Saimiri sciureus and S. boliviensis boliviensis. Am. J. Primatol. 2008, 70, 1177–1180. [Google Scholar] [CrossRef]
- Osterholz, M.; Walter, L.; Roos, C. Retropositional events consolidate the branching order among New World monkey genera. Mol. Phylogenet. Evol. 2009, 50, 507–513. [Google Scholar] [CrossRef]
- Schmitz, J.; Noll, A.; Raabe, C.A.; Churakov, G.; Voss, R.; Kiefmann, M.; Rozhdestvensky, T.; Brosius, J.; Baertsch, R.; Clawson, H.; et al. Genome sequence of the basal haplorrhine primate Tarsius syrichta reveals unusual insertions. Nat. Commun. 2016, 7, 12997. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L. Alu repeats and human genomic diversity. Nat. Rev. Genet. 2002, 3, 370–379. [Google Scholar] [CrossRef]
- Jurka, J.; Smith, T. A fundamental division in the Alu family of repeated sequences. Proc. Natl. Acad. Sci. USA 1988, 85, 4775–4778. [Google Scholar] [CrossRef] [Green Version]
- Bailey, A.D.; Shen, C.K. Sequential insertion of Alu family repeats into specific genomic sites of higher primates. Proc. Natl. Acad. Sci. USA 1993, 90, 7205–7209. [Google Scholar] [CrossRef] [Green Version]
- Britten, R.J.; Baron, W.F.; Stout, D.B.; Davidson, E.H. Sources and evolution of human Alu repeated sequences. Proc. Natl. Acad. Sci. USA 1988, 85, 4770–4774. [Google Scholar] [CrossRef] [Green Version]
- Sawada, I.; Willard, C.; Shen, C.K.; Chapman, B.; Wilson, A.C.; Schmid, C.W. Evolution of Alu family repeats since the divergence of human and chimpanzee. J. Mol. Evol. 1985, 22, 316–322. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L. A human-specific subfamily of Alu sequences. Genomics 1991, 9, 481–487. [Google Scholar] [CrossRef]
- Batzer, M.A.; Deininger, P.L.; Hellmann-Blumberg, U.; Jurka, J.; Labuda, D.; Rubin, C.M.; Schmid, C.W.; Zietkiewicz, E.; Zuckerkandl, E. Standardized nomenclature for Alu repeats. J. Mol. Evol. 1996, 42, 3–6. [Google Scholar] [CrossRef]
- Jurka, J. Sequence patterns indicate an enzymatic involvement in integration of mammalian retroposons. Proc. Natl. Acad. Sci. USA 1997, 94, 1872–1877. [Google Scholar] [CrossRef] [Green Version]
- Kapitonov, V.; Jurka, J. The age of Alu subfamilies. J. Mol. Evol. 1996, 42, 59–65. [Google Scholar] [CrossRef]
- Konkel, M.K.; Walker, J.A.; Batzer, M.A. LINEs and SINEs of Primate Evolution. Evol. Anthropol. 2010, 19, 236–249. [Google Scholar] [CrossRef] [Green Version]
- Worley, K.C.; Warren, W.C.; Rogers, J.; Locke, D.; Muzny, D.M.; Mardis, E.R.; Weinstock, G.M.; Tardif, S.D.; Aagaard, K.M.; Archidiacono, N.; et al. The common marmoset genome provides insight into primate biology and evolution. Nat. Genet. 2014, 46, 850–857. [Google Scholar] [CrossRef]
- Baker, J.N.; Walker, J.A.; Vanchiere, J.A.; Phillippe, K.R.; St Romain, C.P.; Gonzalez-Quiroga, P.; Denham, M.W.; Mierl, J.R.; Konkel, M.K.; Batzer, M.A. Evolution of Alu Subfamily Structure in the Saimiri Lineage of New World Monkeys. Genome Biol. Evol. 2017, 9, 2365–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storer, J.M.; Walker, J.A.; Jordan, V.E.; Batzer, M.A. Sensitivity of the polyDetect computational pipeline for phylogenetic analyses. Anal. Biochem. 2020, 593, 113516. [Google Scholar] [CrossRef] [PubMed]
- Storer, J.M.; Walker, J.A.; Rockwell, C.E.; Mores, G.; Beckstrom, T.O.; Orkin, J.D.; Melin, A.D.; Phillips, K.A.; Roos, C.; Batzer, M.A. Recently Integrated Alu Elements in Capuchin Monkeys: A Resource for Cebus/Sapajus Genomics. Genes 2022, 13, 572. [Google Scholar] [CrossRef]
- Storer, J. Characterization and Amplification of Retrotransposable Elements Platy-1 and Alu in the Cebidae Lineage of Platyrrhine Primates. Doctoral Dissertations, Louisiana State University, Baton Rouge, LA, USA, 2019. 5053. [Google Scholar]
- Martins, A.M., Jr.; Amorim, N.; Carneiro, J.C.; de Mello Affonso, P.R.; Sampaio, I.; Schneider, H. Alu elements and the phylogeny of capuchin (Cebus and Sapajus) monkeys. Am. J. Primatol. 2015, 77, 368–375. [Google Scholar] [CrossRef]
- Ray, D.A.; Xing, J.; Hedges, D.J.; Hall, M.A.; Laborde, M.E.; Anders, B.A.; White, B.R.; Stoilova, N.; Fowlkes, J.D.; Landry, K.E.; et al. Alu insertion loci and platyrrhine primate phylogeny. Mol. Phylogenet. Evol. 2005, 35, 117–126. [Google Scholar] [CrossRef]
- Smit, A.F.A.; Hubley, R.; Green, P. 2013–2015, RepeatMasker Open-4.0. 2015. Available online: http://www.repeatmasker.org (accessed on 19 October 2022).
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef]
- Konkel, M.K.; Walker, J.A.; Hotard, A.B.; Ranck, M.C.; Fontenot, C.C.; Storer, J.; Stewart, C.; Marth, G.T.; Batzer, M.A. Sequence Analysis and Characterization of Active Human Alu Subfamilies Based on the 1000 Genomes Pilot Project. Genome Biol. Evol. 2015, 7, 2608–2622. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony; Version 4.0a147; Sinauer Associates, Inc.: Sunderland, MA, USA, 2003. [Google Scholar]
- Rogers, J.; Raveendran, M.; Harris, R.A.; Mailund, T.; Leppala, K.; Athanasiadis, G.; Schierup, M.H.; Cheng, J.; Munch, K.; Walker, J.A.; et al. The comparative genomics and complex population history of Papio baboons. Sci. Adv. 2019, 5, eaau6947. [Google Scholar] [CrossRef] [Green Version]
- Hirai, H.; Hirai, Y.; Morimoto, M.; Kaneko, A.; Kamanaka, Y.; Koga, A. Night Monkey Hybrids Exhibit De Novo Genomic and Karyotypic Alterations: The First Such Case in Primates. Genome Biol. Evol. 2017, 9, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Storer, J.M.; Walker, J.A.; Rewerts, L.C.; Brown, M.A.; Beckstrom, T.O.; Herke, S.W.; Roos, C.; Batzer, M.A. Owl Monkey Alu Insertion Polymorphisms and Aotus Phylogenetics. Genes 2022, 13, 2069. https://doi.org/10.3390/genes13112069
Storer JM, Walker JA, Rewerts LC, Brown MA, Beckstrom TO, Herke SW, Roos C, Batzer MA. Owl Monkey Alu Insertion Polymorphisms and Aotus Phylogenetics. Genes. 2022; 13(11):2069. https://doi.org/10.3390/genes13112069
Chicago/Turabian StyleStorer, Jessica M., Jerilyn A. Walker, Lydia C. Rewerts, Morgan A. Brown, Thomas O. Beckstrom, Scott W. Herke, Christian Roos, and Mark A. Batzer. 2022. "Owl Monkey Alu Insertion Polymorphisms and Aotus Phylogenetics" Genes 13, no. 11: 2069. https://doi.org/10.3390/genes13112069