
Transcriptome Analysis of the Ovaries of Taihe Black-Bone Silky Fowls at Different Egg-Laying Stages

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Sample Collection
2.2. Ethical Statement
2.3. Library Construction and Sequencing of the Transcriptome
2.4. Bioinformatics Analysis of the Transcriptome
2.5. Differential Expression Analysis
2.6. Analysis of the GO and KEGG Pathways
3. Results
3.1. Sequencing Results and Reading Mapping
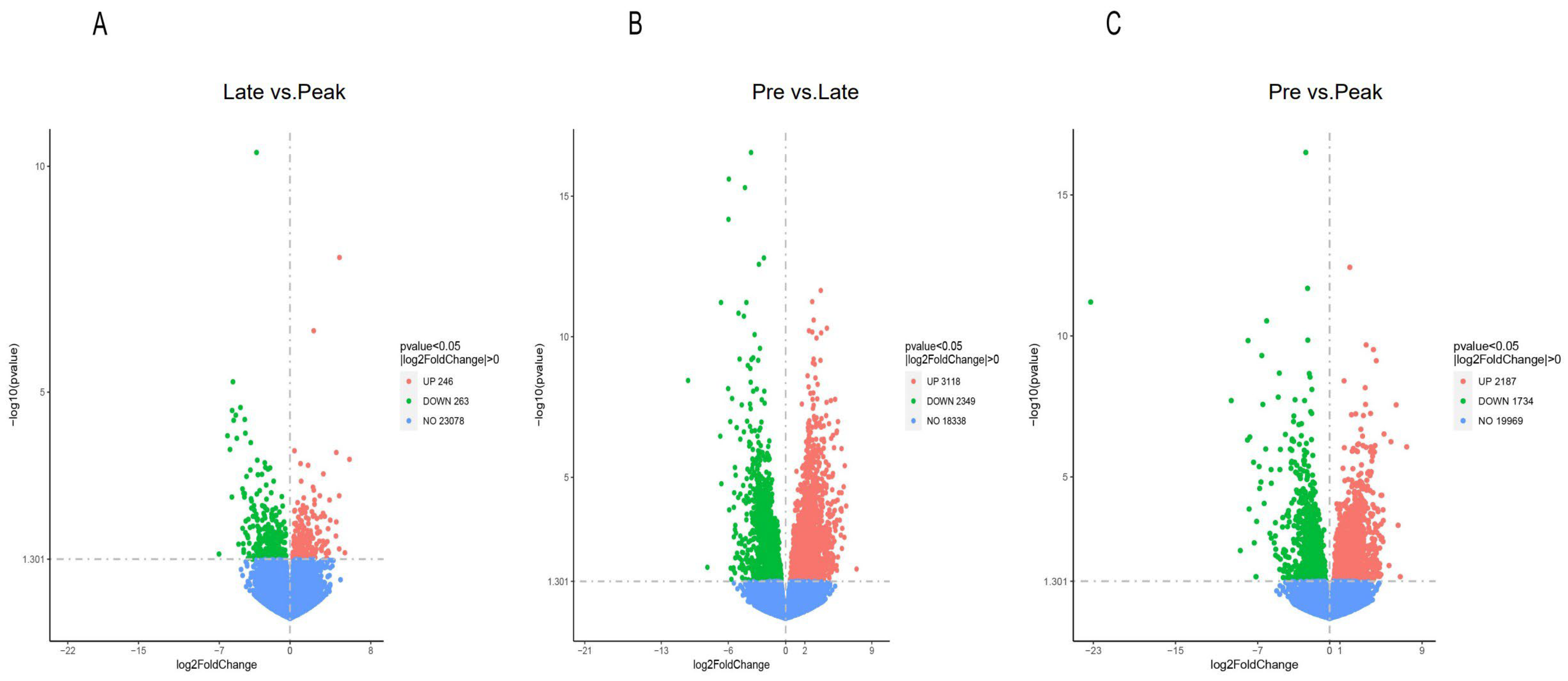
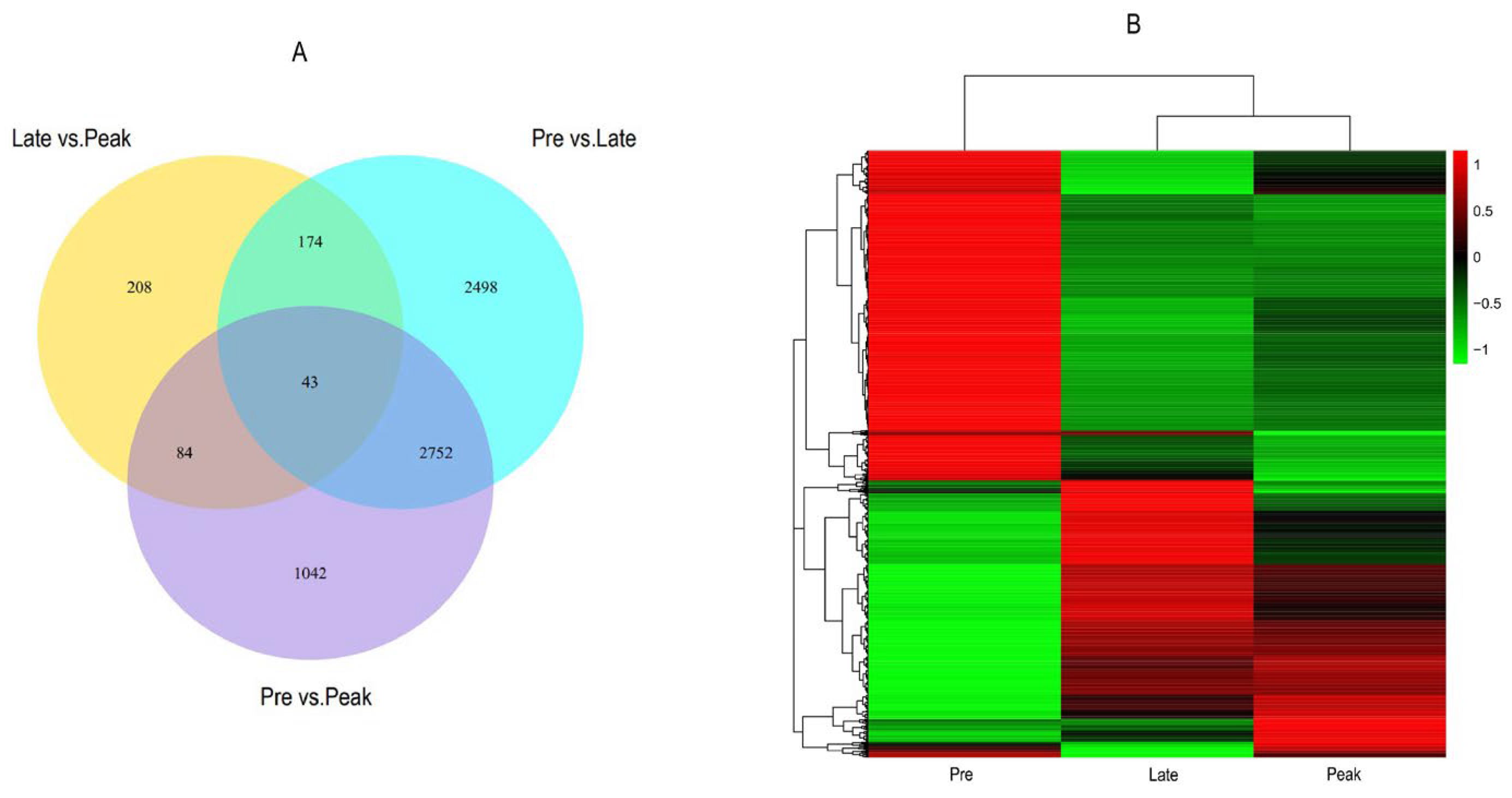
3.2. Differential Expression Genes
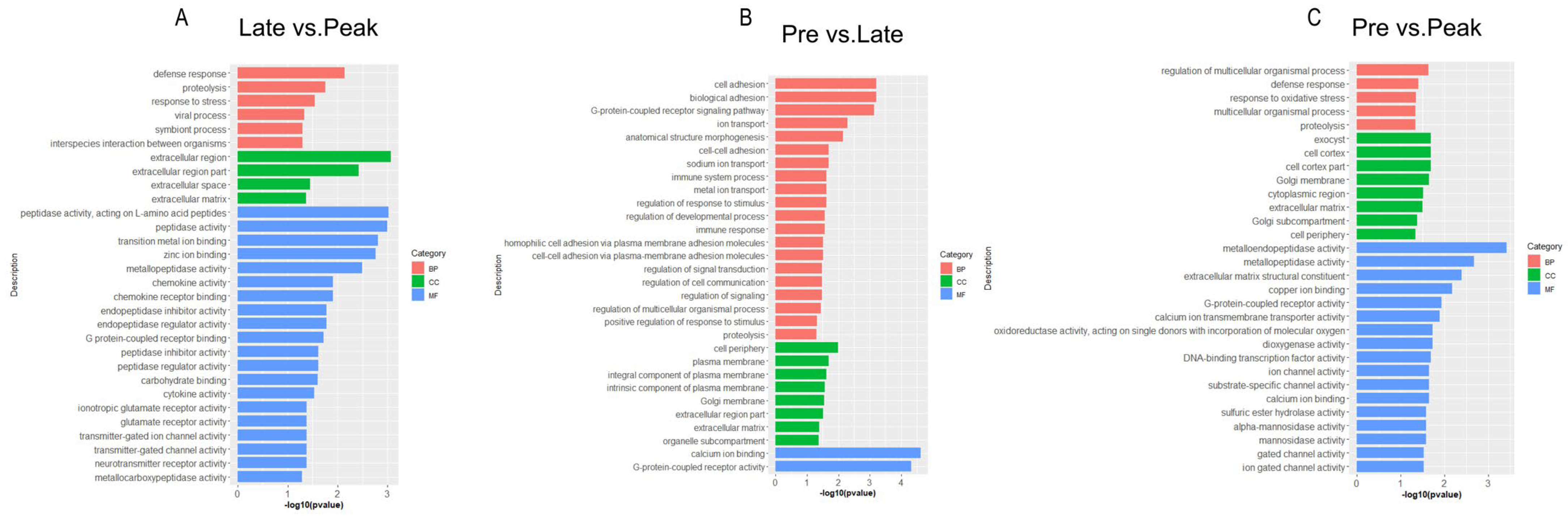
3.3. Gene Ontology Enrichment Analysis
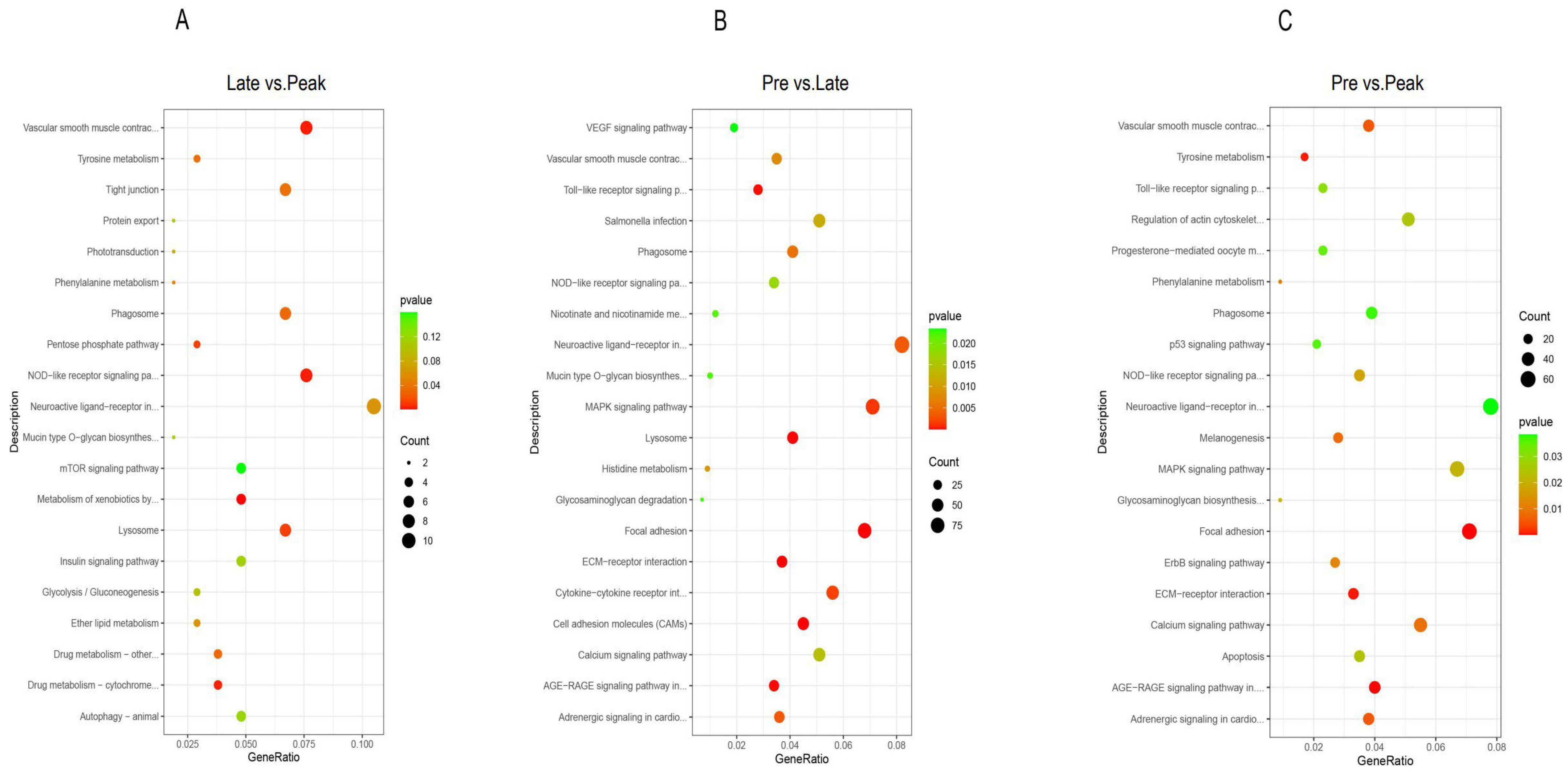
3.4. KEGG Pathway Analysis
4. Discussion
4.1. Analysis of DEGs
4.2. Analysis of GO and KEGG
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mu, R.; Yu, Y.Y.; Gegen, T.; Wen, D.; Wang, F.; Chen, Z.; Xu, W.B. Transcriptome analysis of ovary tissues from low- and high-yielding Changshun green-shell laying hens. BMC Genom. 2021, 22, 349. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Guan, L.F.; Ge, L.Y.; Liu, G.Y.; Bai, Y.J.; Liu, X.L. Nanopore-based full-length transcriptome sequencing of Muscovy duck (Cairina moschata) ovary. Poult. Sci. 2021, 100, 101246. [Google Scholar] [CrossRef] [PubMed]
- Ghanem, K.; Johnson, A.L. Response of hen pre-recruitment ovarian follicles to follicle stimulating hormone, in vivo. Gen. Comp. Endocrinol. 2019, 270, 41–47. [Google Scholar] [CrossRef]
- Sun, X.; Chen, X.X.; Zhao, J.H.; Ma, C.; Yan, C.C.; Liswaniso, S.; Xu, R.F.; Qin, N. Transcriptome comparative analysis of ovarian follicles reveals the key genes and signaling pathways implicated in hen egg production. BMC Genom. 2021, 22, 899. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Chen, B.L.; Zhu, Q.; Xu, Z.X.; Ning, C.Y.; Yin, H.D.; Wang, Y.; Zhao, X.L.; Fan, X.L.; Yang, M.Y.; et al. Transcriptome analysis reveals differentially expressed genes associated with high rates of egg production in chicken hypothalamic-pituitary-ovarian axis. Sci. Rep. 2020, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Finotello, F.; Di Camillo, B. Measuring differential gene expression with RNA-seq: Challenges and strategies for data analysis. Brief. Funct. Genom. 2015, 14, 130–142. [Google Scholar] [CrossRef]
- Tao, Z.; Song, W.; Zhu, C.; Xu, W.; Liu, H.; Zhang, S.; Li, H.F. Comparative transcriptomic analysis of high and low egg-producing duck ovaries. Poult. Sci. 2017, 96, 4378–4388. [Google Scholar] [CrossRef]
- Mi, S.; Shang, K.; Jia, W.; Zhang, C.H.; Li, X.; Fan, Y.Q.; Wang, H. Characterization and discrimination of Taihe black-boned silky fowl (Gallus gallus domesticus Brisson) muscles using LC/MS-based lipidomics. Food Res. Int. 2018, 109, 187–195. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Hu, Z.; Liu, J.; Cao, J.; Zhang, H.; Liu, X. Ovarian transcriptomic analysis of black Muscovy duck at the early, peak and late egg-laying stages. Gene 2021, 777, 145449. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, P.; Cong, G.; Liu, M.; Shi, S.; Shao, D.; Tan, B. Comparative transcriptomic analysis of ovaries from high and low egg-laying Lingyun black-bone chickens. Vet. Med. Sci. 2021, 7, 1867–1880. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.M.; Chiang, J.C.; Lin, Y.C.; Lin, Y.N.; Chuang, P.Y.; Chang, Y.C.; Chen, C.C.; Wu, K.Y.; Hsieh, J.C.; Chen, S.K.; et al. Lysophosphatidic acid receptor LPA3 prevents oxidative stress and cellular senescence in Hutchinson-Gilford progeria syndrome. Aging Cell 2020, 19, e13064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diao, H.L.; Li, R.; El Zowalaty, A.E.; Xiao, S.; Zhao, F.; Dudley, E.A.; Ye, X.Q. Deletion of Lysophosphatidic Acid Receptor 3 (Lpar3) Disrupts Fine Local Balance of Progesterone and Estrogen Signaling in Mouse Uterus During Implantation. Biol. Reprod. 2015, 93, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.C.; Hu, S.Q.; Ouyang, Q.Y.; Wu, T.H.; Lu, Y.; Hu, J.W.; Hu, B.; Li, L.; Wang, J.W. Comparative transcriptome analysis identifies crucial candidate genes and pathways in the hypothalamic-pituitary-gonadal axis during external genitalia development of male geese. BMC Genom. 2022, 23, 136. [Google Scholar] [CrossRef]
- Gokul, C.; Parida, S.; Singh, T.U.; Panigrahi, M.; Suhas, K.S.; Soni, K.; Kesavan, M.; Srivastava, V.; Shyam Kumar, T.S.; Mahobiya, A. Lysophosphatidic acid enhances PGE2 to PGF2alpha ratio and nitric oxide level in nonpregnant buffalo uterus. Theriogenology 2018, 120, 47–55. [Google Scholar] [CrossRef]
- Ye, X.Q.; Hama, K.; Contos, J.J.A.; Anliker, B.; Inoue, A.; Skinner, M.K.; Suzuki, H.; Amano, T.; Kennedy, G.; Arai, H.; et al. LPA3-mediated lysophosphatidic acid signalling in embryo implantation and spacing. Nature 2005, 435, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Brunnert, D.; Sztachelska, M.; Bornkessel, F.; Treder, N.; Wolczynski, S.; Goyal, P.; Zygmunt, M. Lysophosphatidic acid and sphingosine 1-phosphate metabolic pathways and their receptors are differentially regulated during decidualization of human endometrial stromal cells. Mol. Hum. Reprod. 2014, 20, 1016–1025. [Google Scholar] [CrossRef]
- Klemencic, M.; Novinec, M.; Maier, S.; Hartmann, U.; Lenarcic, B. The Heparin-Binding Activity of Secreted Modular Calcium-Binding Protein 1 (SMOC-1) Modulates Its Cell Adhesion Properties. PLoS ONE 2013, 8, e56839. [Google Scholar] [CrossRef] [Green Version]
- Pazin, D.E.; Albrecht, K.H. Developmental Expression of Smoc1 and Smoc2 Suggests Potential Roles in Fetal Gonad and Reproductive Tract Differentiation. Dev. Dyn. 2009, 238, 2877–2890. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Nii, T.; Isobe, N.; Yoshimura, Y. Effects of the routine multiple vaccinations on the expression of innate immune molecules and induction of histone modification in ovarian cells of layer chicks. Poult. Sci. 2019, 98, 5127–5136. [Google Scholar] [CrossRef]
- Grande, G.; Barrachina, F.; Soler-Ventura, A.; Jodar, M.; Mancini, F.; Marana, R.; Chiloiro, S.; Pontecorvi, A.; Oliva, R.; Milardi, D. The Role of Testosterone in Spermatogenesis: Lessons from Proteome Profiling of Human Spermatozoa in Testosterone Deficiency. Front. Endocrinol. 2022, 13, 11. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadou, M.; Avdi, M.; Theodoridis, A.; Michailidis, G. Temporal changes in the expression of avian beta-defensins in the chicken vagina during sexual maturation and Salmonella infection. Vet. Res. Commun. 2013, 37, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadou, M.; Theodoridis, A.; Michailidis, G. Effects of sexual maturation and Salmonella infection on the expression of avian beta-defensin genes in the chicken testis. Vet. Res. Commun. 2014, 38, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Mok, H.P.; Zhuang, J. Secreted modular calcium-binding proteins in pathophysiological processes and embryonic development. Chin. Med. J. 2019, 132, 2476–2484. [Google Scholar] [CrossRef] [PubMed]
- Piltonen, T.T. Polycystic ovary syndrome: Endometrial markers. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 37, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Young, C.H.; Rothfuss, H.M.; Gard, P.F.; Muth, A.; Thompson, P.R.; Ashley, R.L.; Cherrington, B.D. Citrullination regulates the expression of insulin-like growth factor-binding protein 1 (IGFBP1) in ovine uterine luminal epithelial cells. Reproduction 2017, 153, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citrinovitz, A.C.M.; Langer, L.; Strowitzki, T.; Germeyer, A. Resveratrol enhances decidualization of human endometrial stromal cells. Reproduction 2020, 159, 453–463. [Google Scholar] [CrossRef]
- Young, C.H.; Snow, B.; DeVore, S.B.; Mohandass, A.; Nemmara, V.V.; Thompson, P.R.; Thyagarajan, B.; Navratil, A.M.; Cherrington, B.D. Progesterone stimulates histone citrullination to increase IGFBP1 expression in uterine cells. Reproduction 2021, 162, 117–127. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.L.; Zhao, L. A Potential Three-Gene-Based Diagnostic Signature for Hypertension in Pregnancy. Int. J. Gen. Med. 2021, 14, 6847–6856. [Google Scholar] [CrossRef]
- Delmeire, D.; Flamez, D.; Hinke, S.A.; Cali, J.J.; Pipeleers, D.; Schuit, F. Type VIII adenylyl cyclase in rat beta cells: Coincidence signal detector/generator for glucose and GLP-1. Diabetologia 2003, 46, 1383–1393. [Google Scholar] [CrossRef]
- Waddleton, D.; Wu, W.Z.; Feng, Y.; Thompson, C.; Wu, M.; Zhou, Y.P.; Howard, A.; Thornberry, N.; Li, J.; Mancini, J.A. Phosphodiesterase 3 and 4 comprise the major cAMP metabolizing enzymes responsible for insulin secretion in INS-1 (832/13) cells and rat islets. Biochem. Pharmacol. 2008, 76, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.Q.; Jing, Z.X.; Pan, C.G.; Lin, Z.; Zhen, Z.; Hou, L.P.; Dong, Z.D. The progestin norethindrone alters growth, reproductive histology and gene expression in zebrafish (Danio rerio). Chemosphere 2020, 242, 11. [Google Scholar] [CrossRef]
- Bello, S.F.; Xu, H.P.; Guo, L.J.; Li, K.; Zheng, M.; Xu, Y.B.; Zhang, S.Y.; Bekele, E.J.; Bahareldin, A.A.; Zhu, W.J.; et al. Hypothalamic and ovarian transcriptome profiling reveals potential candidate genes in low and high egg production of white Muscovy ducks (Cairina moschata). Poult. Sci. 2021, 100, 13. [Google Scholar] [CrossRef] [PubMed]
- He, H.Y.; Liu, L.L.; Chen, B.; Xiao, H.X.; Liu, W.J. Study on lactation performance and development of KASP marker for milk traits in Xinjiang donkey (Equus asinus). Anim. Biotechnol. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Belli, M.; Shimasaki, S. Molecular Aspects and Clinical Relevance of GDF9 and BMP15 in Ovarian Function. In Ovarian Cycle; Litwack, G., Ed.; Vitamins and Hormones; Elsevier Academic Press Inc.: San Diego, CA, USA, 2018; Volume 107, pp. 317–348. [Google Scholar]
- Simpson, C.M.; Robertson, D.M.; Al-Musawi, S.L.; Heath, D.A.; McNatty, K.P.; Ritter, L.J.; Mottershead, D.G.; Gilchrist, R.B.; Harrison, C.A.; Stanton, P.G. Aberrant GDF9 Expression and Activation Are Associated With Common Human Ovarian Disorders. J. Clin. Endocrinol. Metab. 2014, 99, E615–E624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karagul, M.I.; Aktas, S.; Yilmaz, B.C.; Yilmaz, M.; Temel, G.O. GDF9 and BMP15 Expressions and Fine Structure Changes during Folliculogenesis in Polycystic Ovary Syndrome. Balk. Med. J. 2018, 35, 43–54. [Google Scholar] [CrossRef]
- Myers, M.; Mansouri-Attia, N.; James, R.; Peng, J.; Pangas, S.A. GDF9 Modulates the Reproductive and Tumor Phenotype of Female Inha-Null Mice. Biol. Reprod. 2013, 88, 6. [Google Scholar] [CrossRef] [Green Version]
- Fang, E.; Zhang, H.; Yin, W.L.; Wang, C.Y.; Liu, Y.L.; Li, Y.Z.; Wang, L.L.; Wu, Y.; Zhang, R.Z.; Zou, C.D.; et al. CPEB3 deficiency in mice affect ovarian follicle development and causes premature ovarian insufficiency. Cell Death Dis. 2022, 13, 9. [Google Scholar]
- Stocker, W.A.; Walton, K.L.; Richani, D.; Chan, K.L.; Beilby, K.H.; Finger, B.J.; Green, M.P.; Gilchrist, R.B.; Harrison, C.A. A variant of human growth differentiation factor-9 that improves oocyte developmental competence. J. Biol. Chem. 2020, 295, 7981–7991. [Google Scholar] [CrossRef]
- Cong, Y.Y.; Wu, H.B.; Bian, X.J.; Xie, Q.; Lyu, Q.F.; Cui, J.Q.; Suo, L.; Kuang, Y.P. Ptk2bdeletion improves mice folliculogenesis and fecundity via inhibiting follicle loss mediated by Erk pathway. J. Cell. Physiol. 2021, 236, 1043–1053. [Google Scholar] [CrossRef]
- Meng, X.Q.; Dai, Y.Y.; Jing, L.D.; Bai, J.; Liu, S.Z.; Zheng, K.G.; Pan, J. Subcellular localization of proline-rich tyrosine kinase 2 during oocyte fertilization and early-embryo development in mice. J. Reprod. Dev. 2016, 62, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Kinsey, W.H. PYK2: A calcium-sensitive protein tyrosine kinase activated in response to fertilization of the zebrafish oocyte. Dev. Biol. 2013, 373, 130–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brukman, N.G.; Nunez, S.Y.; Molina, L.D.P.; Buffone, M.G.; Darszon, A.; Cuasnicu, P.S.; Da Ros, V.G. Tyrosine phosphorylation signaling regulates Ca2+ entry by affecting intracellular pH during human sperm capacitation. J. Cell. Physiol. 2019, 234, 5276–5288. [Google Scholar] [CrossRef] [PubMed]
- Talaulikar, V.S.; Manyonda, I. Progesterone and progesterone receptor modulators in the management of symptomatic uterine fibroids. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 165, 135–140. [Google Scholar] [CrossRef]
- Cha, J.Y.; Sun, X.F.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef]
- Kim, H.I.; Kim, T.H.; Yoo, J.Y.; Young, S.L.; Lessey, B.A.; Ku, B.J.; Jeong, J.W. ARID1A and PGR proteins interact in the endometrium and reveal a positive correlation in endometriosis. Biochem. Biophys. Res. Commun. 2021, 550, 151–157. [Google Scholar] [CrossRef]
- Akison, L.K.; Robker, R.L. The Critical Roles of Progesterone Receptor (PGR) in Ovulation, Oocyte Developmental Competence and Oviductal Transport in Mammalian Reproduction. Reprod. Domest. Anim. 2012, 47, 288–296. [Google Scholar]
- Akison, L.K.; Robertson, S.A.; Gonzalez, M.B.; Richards, J.S.; Smith, C.W.; Russell, D.L.; Robker, R.L. Regulation of the ovarian inflammatory response at ovulation by nuclear progesterone receptor. Am. J. Reprod. Immunol. 2018, 79, 12. [Google Scholar] [CrossRef]
- Yaghobi, Z.; Movassaghpour, A.; Talebi, M.; Shadbad, M.A.; Hajiasgharzadeh, K.; Pourvahdani, S.; Baradaran, B. The role of CD44 in cancer chemoresistance: A concise review. Eur. J. Pharmacol. 2021, 903, 174147. [Google Scholar] [CrossRef]
- Mattheolabakis, G.; Milane, L.; Singh, A.; Amiji, M.M. Hyaluronic acid targeting of CD44 for cancer therapy: From receptor biology to nanomedicine. J. Drug Target. 2015, 23, 605–618. [Google Scholar] [CrossRef]
- Paravati, R.; De Mello, N.; Onyido, E.K.; Francis, L.W.; Brusehafer, K.; Younas, K.; Spencer-Harty, S.; Conlan, R.S.; Gonzalez, D.; Margarit, L. Differential regulation of osteopontin and CD44 correlates with infertility status in PCOS patients. J. Mol. Med. 2020, 98, 1713–1725. [Google Scholar] [CrossRef] [PubMed]
- Chaen, T.; Konno, T.; Egashira, M.; Bai, R.L.; Nomura, N.; Nomura, S.; Hirota, Y.; Sakurai, T.; Imakawa, K. Estrogen-Dependent Uterine Secretion of Osteopontin Activates Blastocyst Adhesion Competence. PLoS ONE 2012, 7, e48933. [Google Scholar] [CrossRef] [PubMed]
- Raheem, K.A.; Marei, W.F.; Mifsud, K.; Khalid, M.; Wathes, D.C.; Fouladi-Nashta, A.A. Regulation of the hyaluronan system in ovine endometrium by ovarian steroids. Reproduction 2013, 145, 491–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.; Pandey, S.; Parmar, M.S.; Somal, A.; Paul, A.; Panda, B.S.K.; Bhat, I.A.; Baiju, I.; Bharti, M.K.; Saikumar, G.; et al. Impact of oocyte-secreted factors on its developmental competence in buffalo. Zygote 2017, 25, 313–320. [Google Scholar] [CrossRef]
- Liu, C.; Rodriguez, K.F.; Brown, P.R.; Yao, H.H.C. Reproductive, Physiological, and Molecular Outcomes in Female Mice Deficient in Dhh and Ihh. Endocrinology 2018, 159, 2563–2575. [Google Scholar] [CrossRef] [Green Version]
- Johansson, H.K.L.; Taxvig, C.; Olsen, G.P.M.; Svingen, T. Effects of the Hedgehog Signaling Inhibitor Itraconazole on Developing Rat Ovaries. Toxicol. Sci. 2021, 182, 60–69. [Google Scholar] [CrossRef]
- Lee, K.; Jeong, J.; Kwak, I.; Yu, C.T.; Lanske, B.; Soegiarto, D.W.; Toftgard, R.; Tsai, M.J.; Tsai, S.; Lydon, J.P.; et al. Indian hedgehog is a major mediator of progesterone signaling in the mouse uterus. Nat. Genet. 2006, 38, 1204–1209. [Google Scholar] [CrossRef]
- Welt, C.; Sidis, Y.; Keutmann, H.; Schneyer, A. Activins, inhibins, and follistatins: From endocrinology to signaling. A paradigm for the new millennium. Exp. Biol. Med. 2002, 227, 724–752. [Google Scholar] [CrossRef]
- Sirotkin, A.V. Cytokines: Signalling molecules controlling ovarian functions. Int. J. Biochem. Cell Biol. 2011, 43, 857–861. [Google Scholar]
- Walton, K.L.; Goney, M.P.; Peppas, Z.; Stringer, J.M.; Winship, A.; Hutt, K.; Goodchild, G.; Maskey, S.; Chan, K.L.; Brule, E.; et al. Inhibin Inactivation in Female Mice Leads to Elevated FSH Levels, Ovarian Overstimulation, and Pregnancy Loss. Endocrinology 2022, 163, 13. [Google Scholar] [CrossRef]
- Cui, Z.F.; Liu, L.B.; Zhao, X.L.; Ran, J.S.; Wang, Y.; Yin, H.D.; Li, D.Y.; Zhu, Q. Analysis of Expression and Single Nucleotide Polymorphisms of INHA Gene Associated with Reproductive Traits in Chickens. Biomed Res. Int. 2019, 2019, 8572837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gong, Y.Z. Transcription of CYP19A1 is directly regulated by SF-1 in the theca cells of ovary follicles in chicken. Gen. Comp. Endocrinol. 2017, 247, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cheng, L.; Li, X.L.; Tang, S.X.; Zhang, X.X.; Gong, Y.Z. Transcriptional regulation of CYP19A1 expression in chickens: ESR1, ESR2 and NR5A2 form a functional network. Gen. Comp. Endocrinol. 2022, 315, 12. [Google Scholar] [CrossRef]
- Bhagwat, S.R.; Hajela, K.; Kumar, A. Proteolysis to Identify Protease Substrates: Cleave to Decipher. Proteomics 2018, 18, e1800011. [Google Scholar] [CrossRef]
- Chen, B.L.; Chang, H.M.; Zhang, Z.G.; Cao, Y.X.; Leung, P.C.K. ALK4-SMAD3/4 mediates the effects of activin A on the upregulation of PAI-1 in human granulosa lutein cells. Mol. Cell. Endocrinol. 2020, 505, 10. [Google Scholar] [CrossRef] [PubMed]
- Robker, R.L.; Hennebold, J.D.; Russell, D.L. Coordination of Ovulation and Oocyte Maturation: A Good Egg at the Right Time. Endocrinology 2018, 159, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Slingerland, J.M. Links between oestrogen receptor activation and proteolysis: Relevance to hormone-regulated cancer therapy. Nat. Rev. Cancer 2014, 14, 26–38. [Google Scholar]
- Bentov, Y.; Jurisicova, A.; Kenigsberg, S.; Casper, R.F. What maintains the high intra-follicular estradiol concentration in pre-ovulatory follicles? J. Assist. Reprod. Genet. 2016, 33, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Crespo, D.; Goetz, F.W.; Planas, J.V. Luteinizing hormone induces ovulation via tumor necrosis factor alpha-dependent increases in prostaglandin F2alpha in a nonmammalian vertebrate. Sci. Rep. 2015, 5, 14210. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Kulus, J.; Kulus, M.; Kranc, W.; Jopek, K.; Zdun, M.; Jozkowiak, M.; Jaskowski, J.M.; Piotrowska-Kempisty, H.; Bukowska, D.; Antosik, P.; et al. Transcriptomic Profile of New Gene Markers Encoding Proteins Responsible for Structure of Porcine Ovarian Granulosa Cells. Biology 2021, 10, 1214. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.T.; Xiao, C.; Yang, Z.L.; Deng, J.X.; Yang, X.R. Grade follicles transcriptional profiling analysis in different laying stages in chicken. BMC Genom. 2022, 23, 492. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Ma, Y.F.; Zhao, D.; Mi, Y.L.; Zhang, C.Q. Transcriptome profiling analysis of underlying regulation of growing follicle development in the chicken. Poult. Sci. 2020, 99, 2861–2872. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Muraoka, M.; Kato, Y.; Saga, Y. Decoding the transcriptome of pre-granulosa cells during the formation of primordial follicles in the mouse. Biol. Reprod. 2021, 105, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, L.S.; Yoneda, M.; Zhao, M.; Yingsung, W.; Yoshida, N.; Kitagawa, Y.; Kawamura, K.; Suzuki, T.; Kimata, K. Defect in SHAP-hyaluronan complex causes severe female infertility—A study by inactivation of the bikunin gene in mice. J. Biol. Chem. 2001, 276, 7693–7696. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.W.; Khalil, R.A. Evolving mechanisms of vascular smooth muscle contraction highlight key targets in vascular disease. Biochem. Pharmacol. 2018, 153, 91–122. [Google Scholar] [CrossRef]
- Pang, Y.F.; Thomas, P. Role of mPR alpha (PAQR7) in progesterone-induced Ca2+ decrease in human vascular smooth muscle cells. J. Mol. Endocrinol. 2019, 63, 199–213. [Google Scholar] [CrossRef]
- Alqudah, M.; Al-Shboul, O.; Al Dwairi, A.; Al-U’Dat, D.G.; Alqudah, A. Progesterone inhibitory role on gastrointestinal motility. Physiol. Res. 2022, 71, 193–198. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, L.; Zeng, T.; Du, X.; Tao, Z.; Li, G.; Zhong, S.; Wen, J.; Zhou, C.; Xu, X. Transcriptome analyses of potential regulators of pre- and post-ovulatory follicles in the pigeon (Columba livia). Reprod. Fertil. Dev. 2022, 34, 689–697. [Google Scholar] [CrossRef]
- Mai, Y.; Peng, S.; Li, H.; Gao, Y.; Lai, Z. NOD-like receptor signaling pathway activation: A potential mechanism underlying negative effects of benzo(alpha)pyrene on zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 240, 108935. [Google Scholar] [CrossRef]
- Saxena, M.; Yeretssian, G. NOD-Like Receptors: Master Regulators of Inflammation and Cancer. Front. Immunol. 2014, 5, 32. [Google Scholar]
- Jiang, M.; Wang, W.; Zhang, J.; Wang, C.; Bi, Y.; Li, P.; Yang, S.; Li, J.; Xu, Y.T.; Wang, T. Protective Effects and Possible Mechanisms of Actions of Bushen Cuyun Recipe on Diminished Ovarian Reserve Induced by Cyclophosphamide in Rats. Front. Pharmacol. 2020, 11, 546. [Google Scholar] [CrossRef] [PubMed]
- Martyniak, M.; Zglejc-Waszak, K.; Franczak, A.; Kotwica, G. Transcriptomic analysis of the oviduct of pigs during the peri-conceptional period. Anim. Reprod. Sci. 2018, 197, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Li, P.; Huang, S.; Lkhagvagarav, P.; Zhu, M.; Liang, C.; Jia, C. Identification of key genes and molecular mechanisms associated with low egg production of broiler breeder hens in ad libitum. BMC Genom. 2019, 20, 408. [Google Scholar]
- Candelaria, J.I.; Rabaglino, M.B.; Denicol, A.C. Ovarian preantral follicles are responsive to FSH as early as the primary stage of development. J. Endocrinol. 2020, 247, 153–168. [Google Scholar] [CrossRef]
- Esencan, E.; Jiang, Z.; Wang, T.; Zhang, M.; Soylemez-Imamoglu, G.; Seli, E. Impaired Mitochondrial Stress Response due to CLPP Deletion Is Associated with Altered Mitochondrial Dynamics and Increased Apoptosis in Cumulus Cells. Reprod. Sci. 2020, 27, 621–630. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Total Map | Q20 | Q30 | GC pct |
|---|---|---|---|---|---|---|
| Pre_1 | 45,325,812 | 41,798,666 | 38,517,440 (92.15%) | 97.3% | 92.85% | 50.26% |
| Pre_2 | 52,044,340 | 48,173,986 | 43,445,903 (90.19%) | 96.89% | 92.24% | 48.89% |
| Pre_3 | 44,553,380 | 40,271,656 | 36,994,662 (91.86%) | 97.18% | 92.71% | 50.83% |
| Pre_4 | 45,781,184 | 41,751,192 | 37,993,982 (91.0%) | 96.95% | 92.27% | 48.16% |
| Peak_1 | 47,304,680 | 43,650,052 | 39,559,715 (90.63%) | 96.95% | 92.21% | 50.97% |
| Peak_2 | 43,973,992 | 40,910,080 | 37,503,629 (91.67%) | 97.21% | 92.76% | 50.4% |
| Peak_3 | 46,869,756 | 43,034,208 | 39,548,737 (91.9%) | 97.21% | 92.76% | 50.44% |
| Peak_4 | 49,189,518 | 45,234,226 | 41,466,481 (91.67%) | 97.14% | 92.61% | 50.54% |
| Late_1 | 45,141,060 | 41,969,306 | 38,109,595 (90.8%) | 96.99% | 92.34% | 52.02% |
| Late_2 | 44,914,334 | 41,107,798 | 37,444,801 (91.09%) | 97.18% | 92.71% | 51.4% |
| Late_3 | 43,823,110 | 40,425,332 | 36,598,028 (90.53%) | 97.1% | 92.62% | 51.44% |
| Late_4 | 47,357,330 | 43,244,028 | 39,397,767 (91.11%) | 97.09% | 92.5% | 50.8% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, X.; Huang, X.; Wang, J.; Zhang, H.; Zhou, W.; Xu, C.; Huang, Y.; Tan, Y.; Yin, Z. Transcriptome Analysis of the Ovaries of Taihe Black-Bone Silky Fowls at Different Egg-Laying Stages. Genes 2022, 13, 2066. https://doi.org/10.3390/genes13112066
Xiang X, Huang X, Wang J, Zhang H, Zhou W, Xu C, Huang Y, Tan Y, Yin Z. Transcriptome Analysis of the Ovaries of Taihe Black-Bone Silky Fowls at Different Egg-Laying Stages. Genes. 2022; 13(11):2066. https://doi.org/10.3390/genes13112066
Chicago/Turabian StyleXiang, Xin, Xuan Huang, Jianfeng Wang, Haiyang Zhang, Wei Zhou, Chunhui Xu, Yunyan Huang, Yuting Tan, and Zhaozheng Yin. 2022. "Transcriptome Analysis of the Ovaries of Taihe Black-Bone Silky Fowls at Different Egg-Laying Stages" Genes 13, no. 11: 2066. https://doi.org/10.3390/genes13112066