
The Potential of MicroRNAs as Clinical Biomarkers to Aid Ovarian Cancer Diagnosis and Treatment

Abstract
:1. Introduction
1.1. Ovarian Cancer: A Heterogenous Disease
1.2. Biomarkers and Ovarian Cancer
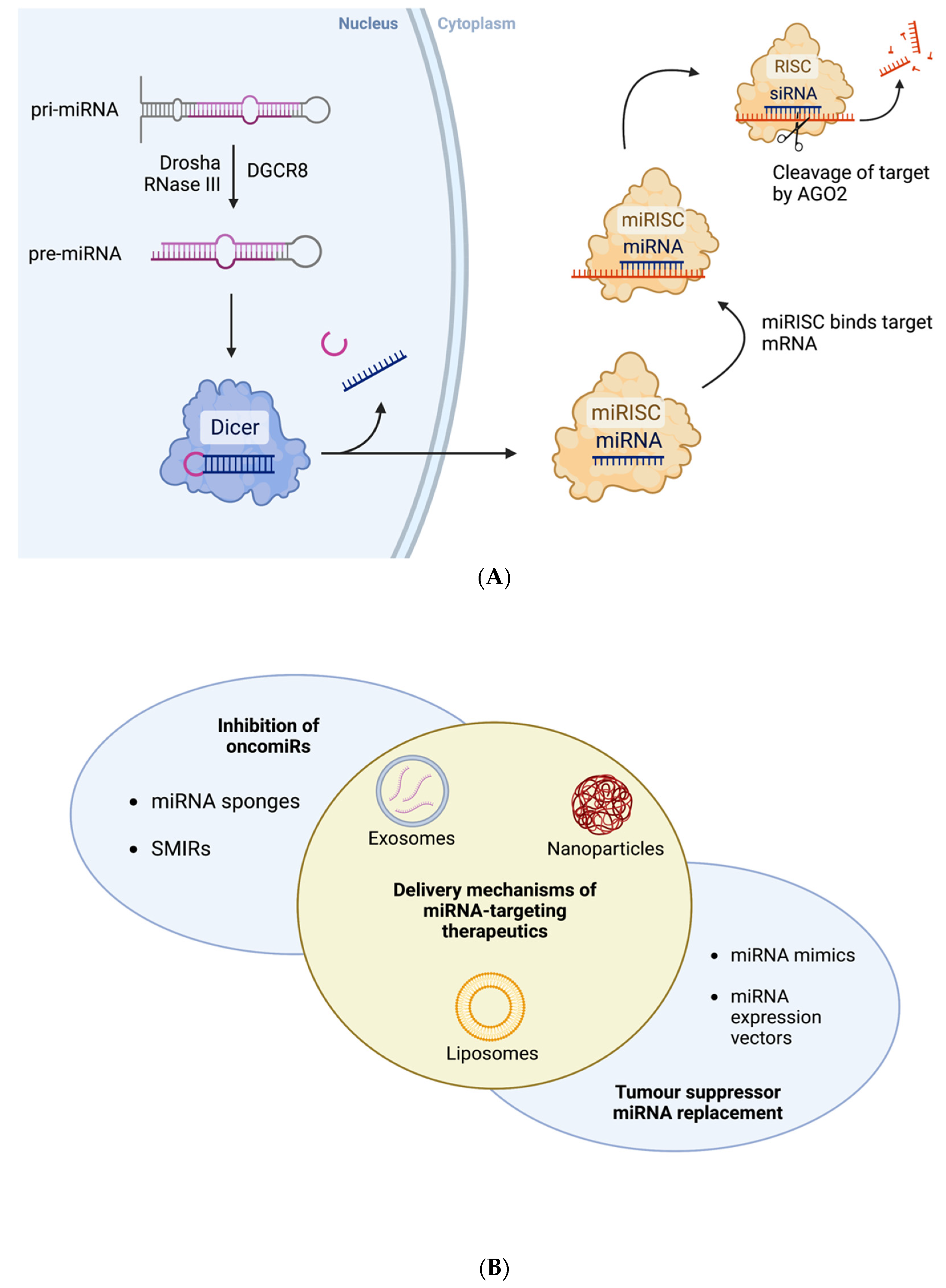
1.3. MicroRNAs as Cancer Biomarkers
1.4. MiRNAs as Therapeutic Agents
1.5. Tumour Suppressor MiRNAs Associated with Ovarian Cancer
1.6. OncomiRs Associated with Ovarian Cancer
1.7. Using MiRNAs to Differentiate between Female Cancers
1.8. Using MiRNAs to Determine Ovarian Cancer Subtype
2. Limitations
3. Future Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atallah, G.A.; Abd Aziz, N.H.; Teik, C.K.; Shafiee, M.N.; Kampan, N.C. New Predictive Biomarkers for Ovarian Cancer. Diagnostics 2021, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Alshamrani, A.A. Roles of microRNAs in Ovarian Cancer Tumorigenesis: Two Decades Later, What Have We Learned? Front. Oncol. 2020, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.; Ralyea, C.; Lockwood, S. Ovarian Cancer: An Integrated Review. Semin. Oncol. Nurs. 2019, 35, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Koshiyama, M.; Matsumura, N.; Konishi, I. Subtypes of Ovarian Cancer and Ovarian Cancer Screening. Diagnostics 2017, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Ireland, N.C.R. Cancer in Ireland 1994–2019: Annual Report of the National Cancer Registry; NCRI: Cork, Ireland, 2021. [Google Scholar]
- Lheureux, S.; Braunstein, M.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef] [Green Version]
- Kossaï, M.; Leary, A.; Scoazec, J.Y.; Genestie, C. Ovarian Cancer: A Heterogeneous Disease. Pathobiology 2018, 85, 41–49. [Google Scholar] [CrossRef]
- Lisio, M.-A.; Fu, L.; Goyeneche, A.; Gao, Z.-H.; Telleria, C. High-Grade Serous Ovarian Cancer: Basic Sciences, Clinical and Therapeutic Standpoints. Int. J. Mol. Sci. 2019, 20, 952. [Google Scholar] [CrossRef] [Green Version]
- Biomarkers Definitions Working Group; Atkinson, A.; Colburn, W.; Degruttola, V.; Demets, D.; Downing, G.; Hoth, D.; Oates, J.; Peck, C.; Schooley, R.; et al. Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar]
- Davey, M.G.; Hynes, S.O.; Kerin, M.J.; Miller, N.; Lowery, A.J. Ki-67 as a Prognostic Biomarker in Invasive Breast Cancer. Cancers 2021, 13, 4455. [Google Scholar] [CrossRef]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Davey, M.G.; Richard, V.; Lowery, A.J.; Kerin, M.J. OncotypeDX© Recurrence Score in BRCA mutation carriers: A systematic review and meta-analysis. Eur. J. Cancer 2021, 154, 209–216. [Google Scholar] [CrossRef]
- McVeigh, T.P.; Kerin, M.J. Clinical use of the Oncotype DX genomic test to guide treatment decisions for patients with invasive breast cancer. Breast Cancer 2017, 9, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlou, M.P.; Diamandis, E.P.; Blasutig, I.M. The Long Journey of Cancer Biomarkers from the Bench to the Clinic. Clin. Chem. 2013, 59, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruthi, S.; Gostout, B.S.; Lindor, N.M. Identification and Management of Women with BRCA Mutations or Hereditary Predisposition for Breast and Ovarian Cancer. Mayo Clin. Proc. 2010, 85, 1111–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.S.; Ren, S.F.; Jiang, W.; Lu, J.Q.; Zhang, X.Y.; Li, X.P.; Cao, R.; Xu, C.J. CA125-Tn ELISA assay improves specificity of pre-operative diagnosis of ovarian cancer among patients with elevated serum CA125 levels. Ann. Transl. Med. 2021, 9, 788. [Google Scholar] [CrossRef]
- Al-Musalhi, K.; Al-Kindi, M.; Ramadhan, F.; Al-Rawahi, T.; Al-Hatali, K.; Mula-Abed, W.A. Validity of Cancer Antigen-125 (CA-125) and Risk of Malignancy Index (RMI) in the Diagnosis of Ovarian Cancer. Oman Med. J. 2015, 30, 428–434. [Google Scholar] [CrossRef]
- Kafali, H.; Artuc, H.; Demir, N. Use of CA125 fluctuation during the menstrual cycle as a tool in the clinical diagnosis of endometriosis; a preliminary report. Eur. J. Obs. Gynecol. Reprod. Biol. 2004, 116, 85–88. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, J.; Cao, Y. Factors associated with serum CA125 level in women without ovarian cancer in the United States: A population-based study. BMC Cancer 2022, 22, 544. [Google Scholar] [CrossRef]
- Gamble, P.; Jaroensri, R.; Wang, H.; Tan, F.; Moran, M.; Brown, T.; Flament-Auvigne, I.; Rakha, E.A.; Toss, M.; Dabbs, D.J. Determining breast cancer biomarker status and associated morphological features using deep learning. Commun. Med. 2021, 1, 14. [Google Scholar] [CrossRef]
- Amuran, G.G.; Eyuboglu, I.P.; Tinay, I.; Akkiprik, M. New Insights in Bladder Cancer Diagnosis: Urinary miRNAs and Proteins. Med. Sci. 2018, 6, 113. [Google Scholar] [CrossRef] [Green Version]
- Zamay, T.N.; Zamay, G.S.; Kolovskaya, O.S.; Zukov, R.A.; Petrova, M.M.; Gargaun, A.; Berezovski, M.V.; Kichkailo, A.S. Current and Prospective Protein Biomarkers of Lung Cancer. Cancers 2017, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Davey, M.G.; Casey, M.C.; McGuire, A.; Waldron, R.M.; Paganga, M.; Holian, E.; Newell, J.; Heneghan, H.M.; McDermott, A.M.; Keane, M.M.; et al. Evaluating the Role of Circulating MicroRNAs to Aid Therapeutic Decision Making for Neoadjuvant Chemotherapy in Breast Cancer—A Prospective, Multicenter Clinical Trial. Ann. Surg. 2022, 276, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Heneghan, H.M.; Miller, N.; Lowery, A.J.; Sweeney, K.J.; Kerin, M.J. MicroRNAs as Novel Biomarkers for Breast Cancer. J. Oncol. 2009, 2009, 950201. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.G.; Davies, M.; Lowery, A.J.; Miller, N.; Kerin, M.J. The Role of MicroRNA as Clinical Biomarkers for Breast Cancer Surgery and Treatment. Int. J. Mol. Sci. 2021, 22, 8290. [Google Scholar] [CrossRef]
- Davey, M.G.; Feeney, G.; Annuk, H.; Paganga, M.; Holian, E.; Lowery, A.J.; Kerin, M.J.; Miller, N. MicroRNA Expression Profiling Predicts Nodal Status and Disease Recurrence in Patients Treated with Curative Intent for Colorectal Cancer. Cancers 2022, 14, 2109. [Google Scholar] [CrossRef]
- Richard, V.; Davey, M.G.; Annuk, H.; Miller, N.; Dwyer, R.M.; Lowery, A.; Kerin, M.J. MicroRNAs in Molecular Classification and Pathogenesis of Breast Tumors. Cancers 2021, 13, 5332. [Google Scholar] [CrossRef]
- Chen, S.N.; Chang, R.; Lin, L.T.; Chern, C.U.; Tsai, H.W.; Wen, Z.H.; Li, Y.H.; Li, C.J.; Tsui, K.H. MicroRNA in Ovarian Cancer: Biology, Pathogenesis, and Therapeutic Opportunities. Int. J. Environ. Res. Public Health 2019, 16, 1510. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [PubMed]
- Pritchard, C.C.; Cheng, H.H.; Tewari, M. MicroRNA profiling: Approaches and considerations. Nat. Rev. Genet. 2012, 13, 358–369. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M.; Kerin, M.J.; Miller, N. Identification and Validation of miRNAs as Endogenous Controls for RQ-PCR in Blood Specimens for Breast Cancer Studies. PLoS ONE 2014, 8, e83718. [Google Scholar] [CrossRef] [Green Version]
- Kheirelseid, E.A.; Chang, K.H.; Newell, J.; Kerin, M.J.; Miller, N. Identification of endogenous control genes for normalisation of real-time quantitative PCR data in colorectal cancer. BMC Mol. Biol. 2010, 11, 12. [Google Scholar]
- Shah, M.Y.; Ferrajoli, A.; Sood, A.K.; Lopez-Berestein, G.; Calin, G.A. microRNA therapeutics in cancer—An emerging concept. EBioMedicine 2016, 12, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Jung, E.-J.; Shah, M.Y.; Lu, C.; Spizzo, R.; Shimizu, M.; Han, H.D.; Ivan, C.; Rossi, S.; Zhang, X.; et al. Therapeutic Synergy between microRNA and siRNA in Ovarian Cancer Treatment. Cancer Discov. 2013, 3, 1302–1315. [Google Scholar]
- Ebert, M.S.; Neilson, J.R.; Sharp, P.A. MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat. Methods 2007, 4, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Monroig Pdel, C.; Chen, L.; Zhang, S.; Calin, G.A. Small molecule compounds targeting miRNAs for cancer therapy. Adv. Drug Deliv. Rev. 2015, 81, 104–116. [Google Scholar]
- Bader, A.G.; Brown, D.; Winkler, M. The Promise of MicroRNA Replacement Therapy. Cancer Res. 2010, 70, 7027–7030. [Google Scholar]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar]
- Wang, C.Z.; Deng, F.; Li, H.; Wang, D.D.; Zhang, W.; Ding, L.; Tang, J.H. MiR-101: A potential therapeutic target of cancers. Am. J. Transl. Res. 2018, 10, 3310–3321. [Google Scholar] [PubMed]
- Wang, J.; Zeng, H.; Li, H.; Chen, T.; Wang, L.; Zhang, K.; Chen, J.; Wang, R.; Li, Q.; Wang, S. MicroRNA-101 Inhibits Growth, Proliferation and Migration and Induces Apoptosis of Breast Cancer Cells by Targeting Sex-Determining Region Y-Box 2. Cell. Physiol. Biochem. 2017, 43, 717–732. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yao, J.; Sun, H.; He, K.; Tong, D.; Song, T.; Huang, C. MicroRNA-101 suppresses progression of lung cancer through the PTEN/AKT signaling pathway by targeting DNA methyltransferase 3A. Oncol. Lett. 2017, 13, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Guo, J.; Yu, L.; Cai, J.; Gui, T.; Tang, H.; Song, L.; Wang, J.; Han, F.; Yang, C.; et al. miR-101 regulates expression of EZH2 and contributes to progression of and cisplatin resistance in epithelial ovarian cancer. Tumor Biol. 2014, 35, 12619–12626. [Google Scholar] [CrossRef]
- Coughlan, A.Y.; Testa, G. Exploiting epigenetic dependencies in ovarian cancer therapy. Int. J. Cancer 2021, 149, 1732–1743. [Google Scholar] [CrossRef]
- Wei, M.; Jin, H.; Yang, S.; Li, Z.; Wang, X.; Li, L.; Jia, Y.; Cui, M. MicroRNA-101 inhibits growth and metastasis of human ovarian cancer cells by targeting PI3K/AKT. Arch. Med. Sci. 2021, 17, 127–134. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chen, J.; He, L.; Stiles, B.L. PTEN: Tumor Suppressor and Metabolic Regulator. Front. Endocrinol. 2018, 9, 338. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ma, H. MiRNA-584 suppresses the progression of ovarian cancer by negatively regulating LPIN1. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1062–1071. [Google Scholar]
- Liu, C.H.; Jing, X.N.; Liu, X.L.; Qin, S.Y.; Liu, M.W.; Hou, C.H. Tumor-suppressor miRNA-27b-5p regulates the growth and metastatic behaviors of ovarian carcinoma cells by targeting CXCL1. J. Ovarian Res. 2020, 13, 92. [Google Scholar] [CrossRef]
- Li, H.; Xu, Y.; Zhao, D. MicroRNA-193b regulates human ovarian cancer cell growth via targeting STMN1. Exp. Ther. Med. 2020, 20, 3310–3315. [Google Scholar] [CrossRef]
- Ysrafil, Y.; Astuti, I.; Anwar, S.L.; Martien, R.; Sumadi, F.A.N.; Wardhana, T.; Haryana, S.M. MicroRNA-155-5p Diminishes in Vitro Ovarian Cancer Cell Viability by Targeting HIF1α Expression. Adv. Pharm. Bull. 2020, 10, 630–637. [Google Scholar] [CrossRef] [PubMed]
- Chen, J. miRNA-195 suppresses cell proliferation of ovarian cancer cell by regulating VEGFR2 and AKT signaling pathways. Mol. Med. Rep. 2018, 18, 1666–1673. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, E.J.; Jeon, M.J. MicroRNAs 125a and 125b inhibit ovarian cancer cells through post-transcriptional inactivation of EIF4EBP1. Oncotarget 2016, 7, 8726–8742. [Google Scholar] [CrossRef] [Green Version]
- Biamonte, F.; Santamaria, G.; Sacco, A.; Perrone, F.M.; di Cello, A.; Battaglia, A.M.; Salatino, A.; di Vito, A.; Aversa, I.; Venturella, R.; et al. MicroRNA let-7g acts as tumor suppressor and predictive biomarker for chemoresistance in human epithelial ovarian cancer. Sci. Rep. 2019, 9, 5668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Li, G.; Liu, S.; Wang, H.; Chen, B. MiR-585-3p suppresses tumor proliferation and migration by directly targeting CAPN9 in high grade serous ovarian cancer. J. Ovarian Res. 2021, 14, 90. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, J.; He, J.; Yi, N. MicroRNA-193a-5p exerts a tumor suppressive role in epithelial ovarian cancer by modulating RBBP6. Mol. Med. Rep. 2021, 24, 582. [Google Scholar] [CrossRef]
- Chirshev, E.; Oberg, K.C.; Ioffe, Y.J.; Unternaehrer, J.J. Let-7 as biomarker, prognostic indicator, and therapy for precision medicine in cancer. Clin. Transl. Med. 2019, 8, 24. [Google Scholar] [CrossRef]
- De Santis, C.; Götte, M. The Role of microRNA Let-7d in Female Malignancies and Diseases of the Female Reproductive Tract. Int. J. Mol. Sci. 2021, 22, 7359. [Google Scholar] [CrossRef]
- Thammaiah, C.K.; Jayaram, S. Role of let-7 family microRNA in breast cancer. Non-Coding RNA Res. 2016, 1, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Perdas, E.; Stawski, R.; Nowak, D.; Zubrzycka, M. The Role of miRNA in Papillary Thyroid Cancer in the Context of miRNA Let-7 Family. Int. J. Mol. Sci. 2016, 17, 909. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Li, J.; Che, G. Prognostic value of let-7 in lung cancer: Systematic review and meta-analysis. Transl. Cancer Res. 2020, 9, 6354–6361. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, R.; Kawada, K.; Sakai, Y. The Molecular Basis and Therapeutic Potential of Let-7 MicroRNAs against Colorectal Cancer. Can. J. Gastroenterol. Hepatol. 2018, 2018, 5769591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, J.; Yang, C.; Yang, Q.; Ding, H.; Jia, J.; Guo, J.; Wang, J.; Wang, Z. Deregulation of let-7e in epithelial ovarian cancer promotes the development of resistance to cisplatin. Oncogenesis 2013, 2, e75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Ouyang, Y.; Che, J.; Li, X.; Zhao, Y.; Yang, K.; Zhao, X.; Chen, Y.; Fan, C.; Yuan, W. Potential Value of miR-221/222 as Diagnostic, Prognostic, and Therapeutic Biomarkers for Diseases. Front. Immunol. 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, Q.; Huang, H.; Li, Y.; Li, L.; Hou, W.; You, Z. Overexpression of miRNA-221 promotes cell proliferation by targeting the apoptotic protease activating factor-1 and indicates a poor prognosis in ovarian cancer. Int. J. Oncol. 2017, 50, 1087–1096. [Google Scholar] [CrossRef] [Green Version]
- Humphries, B.; Yang, C. The microRNA-200 family: Small molecules with novel roles in cancer development, progression and therapy. Oncotarget 2015, 6, 6472–6498. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Su, X.; Zhang, X.; Zhang, Y.; Li, C.; Niu, W.; Liu, C.; Qu, K. miRNA-200a/c as potential biomarker in epithelial ovarian cancer (EOC): Evidence based on miRNA meta-signature and clinical investigations. Oncotarget 2016, 7, 81621–81633. [Google Scholar] [CrossRef]
- Meng, X.; Joosse, S.A.; Müller, V.; Trillsch, F.; Milde-Langosch, K.; Mahner, S.; Geffken, M.; Pantel, K.; Schwarzenbach, H. Diagnostic and prognostic potential of serum miR-7, miR-16, miR-25, miR-93, miR-182, miR-376a and miR-429 in ovarian cancer patients. Br. J. Cancer 2015, 113, 1358–1366. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, M.-J.; Ren, A.-M.; Wu, H.-F.; Han, W.-M.; Tan, R.-Y.; Tu, R.-Q. A Ten-MicroRNA Signature Identified from a Genome-Wide MicroRNA Expression Profiling in Human Epithelial Ovarian Cancer. PLoS ONE 2014, 9, e96472. [Google Scholar]
- Liu, J.; Yoo, J.; Ho, J.Y.; Jung, Y.; Lee, S.; Hur, S.Y.; Choi, Y.J. Plasma-derived exosomal miR-4732-5p is a promising noninvasive diagnostic biomarker for epithelial ovarian cancer. J. Ovarian Res. 2021, 14, 59. [Google Scholar] [CrossRef]
- Wahab, N.A.; Othman, Z.; Nasri, N.W.M.; Mokhtar, M.H.; Ibrahim, S.F.; Hamid, A.A.; Ali, R.A.R.; Mokhtar, N.M. Inhibition of miR-141 and miR-200a Increase DLC-1 and ZEB2 Expression, Enhance Migration and Invasion in Metastatic Serous Ovarian Cancer. Int. J. Environ. Res. Public Health 2020, 17, 2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Choi, M.C.; Jeong, J.Y.; Hwang, S.; Jung, S.G.; Joo, W.D.; Park, H.; Song, S.H.; Lee, C.; Kim, T.H.; et al. Serum exosomal miRNA-145 and miRNA-200c as promising biomarkers for preoperative diagnosis of ovarian carcinomas. J. Cancer 2019, 10, 1958–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Deng, Y.; Liu, J.; Ye, Z.; You, Z.; Yao, S.; He, S. MiR-760 overexpression promotes proliferation in ovarian cancer by downregulation of PHLPP2 expression. Gynecol. Oncol. 2016, 143, 655–663. [Google Scholar] [CrossRef]
- Hirschfeld, M.; Ge, I.; Rücker, G.; Waldschmidt, J.; Mayer, S.; Jäger, M.; Voigt, M.; Kammerer, B.; Nöthling, C.; Berner, K.; et al. Mutually distinguishing microRNA signatures of breast, ovarian and endometrial cancers in vitro. Mol. Med. Rep. 2020, 22, 4048–4060. [Google Scholar] [CrossRef]
- Hui, P.; Gysler, S.M.; Uduman, M.; Togun, T.A.; Prado, D.E.; Brambs, C.E.; Nallur, S.; Schwartz, P.E.; Rutherford, T.J.; Santin, A.D.; et al. MicroRNA signatures discriminate between uterine and ovarian serous carcinomas. Hum. Pathol. 2018, 76, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Harbin, L.M.; Gallion, H.H.; Allison, D.B.; Kolesar, J.M. Next Generation Sequencing and Molecular Biomarkers in Ovarian Cancer—An Opportunity for Targeted Therapy. Diagnostics 2022, 12, 842. [Google Scholar] [CrossRef]
- Agostini, A.; Brunetti, M.; Davidson, B.; Tropé, C.G.; Eriksson, A.G.Z.; Heim, S.; Panagopoulos, I.; Micci, F. The microRNA miR-192/215 family is upregulated in mucinous ovarian carcinomas. Sci. Rep. 2018, 8, 11069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanaihara, N.; Noguchi, Y.; Saito, M.; Takenaka, M.; Takakura, S.; Yamada, K.; Okamoto, A. MicroRNA Gene Expression Signature Driven by miR-9 Overexpression in Ovarian Clear Cell Carcinoma. PLoS ONE 2016, 11, e0162584. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, G.; Wang, G.; Zhao, J.; Wang, B.; Yu, X.; Ding, Y. Profile of differentially expressed miRNAs in high-grade serous carcinoma and clear cell ovarian carcinoma, and the expression of miR-510 in ovarian carcinoma. Mol. Med. Rep. 2015, 12, 8021–8031. [Google Scholar] [CrossRef] [Green Version]
- Lopacinska-Joergensen, J.; Oliveira, D.V.N.P.; Hoegdall, C.K.; Hoegdall, E.V. Identification of Stably Expressed Reference microRNAs in Epithelial Ovarian Cancer. In Vivo 2022, 36, 1059. [Google Scholar] [CrossRef]
- Bignotti, E.; Calza, S.; Tassi, R.A.; Zanotti, L.; Bandiera, E.; Sartori, E.; Odicino, F.E.; Ravaggi, A.; Todeschini, P.; Romani, C. Identification of stably expressed reference small non-coding RNAs for microRNA quantification in high-grade serous ovarian carcinoma tissues. J. Cell. Mol. Med. 2016, 20, 2341–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kura, B.; Parikh, M.; Slezak, J.; Pierce, G.N. The Influence of Diet on MicroRNAs that Impact Cardiovascular Disease. Molecules 2019, 24, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momi, N.; Kaur, S.; Rachagani, S.; Ganti, A.K.; Batra, S.K. Smoking and microRNA dysregulation: A cancerous combination. Trends Mol. Med. 2014, 20, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, J.; Woldhuis, R.R.; Boudewijn, I.M.; van den Berg, A.; Kluiver, J.; Kok, K.; Terpstra, M.M.; Guryev, V.; de Vries, M.; Vermeulen, C.J.; et al. Age-related gene and miRNA expression changes in airways of healthy individuals. Sci. Rep. 2019, 9, 3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Yang, W.; Shi, J.; Zhou, Y.; Yang, J.; Cui, Q.; Zhou, Y. Identification and Analysis of Human Sex-biased MicroRNAs. Genom. Proteom. Bioinform. 2018, 16, 200–211. [Google Scholar] [CrossRef]
- Bast, R.C.; Han, C.Y.; Lu, Z.; Lu, K.H. Next steps in the early detection of ovarian cancer. Commun. Med. 2021, 1, 36. [Google Scholar] [CrossRef]
- Sørensen, C.G.; Karlsson, W.K.; Pommergaard, H.-C.; Burcharth, J.; Rosenberg, J. The diagnostic accuracy of carcinoembryonic antigen to detect colorectal cancer recurrence—A systematic review. Int. J. Surg. 2016, 25, 134–144. [Google Scholar] [CrossRef]
- Talaat, A.; Helmy, M.A.; Saadawy, S.F. Evaluation of miRNA-21 and CA-125 as a promising diagnostic biomarker in patients with ovarian cancer. Egypt. J. Med. Hum. Genet. 2022, 23, 123. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Savolainen, K.; Scaravilli, M.; Ilvesmäki, A.; Staff, S.; Tolonen, T.; Mäenpää, J.U.; Visakorpi, T.; Auranen, A. Expression of the miR-200 family in tumor tissue, plasma and urine of epithelial ovarian cancer patients in comparison to benign counterparts. BMC Res. Notes 2020, 13, 311. [Google Scholar] [CrossRef]
- Rapado-González, Ó.; Majem, B.; Muinelo-Romay, L.; Álvarez-Castro, A.; Santamaría, A.; Gil-Moreno, A.; López-López, R.; Suárez-Cunqueiro, M.M. Human salivary microRNAs in Cancer. J. Cancer 2018, 9, 638–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setti, G.; Pezzi, M.E.; Viani, M.V.; Pertinhez, T.A.; Cassi, D.; Magnoni, C.; Bellini, P.; Musolino, A.; Vescovi, P.; Meleti, M. Salivary MicroRNA for Diagnosis of Cancer and Systemic Diseases: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davey, M.G.; Lowery, A.J.; Miller, N.; Kerin, M.J. MicroRNA Expression Profiles and Breast Cancer Chemotherapy. Int. J. Mol. Sci. 2021, 22, 10812. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, X.; Wang, G.; Wang, B.; Ding, Y.; Zhao, J.; Liu, H.; Cui, S. miR-206 as a prognostic and sensitivity biomarker for platinum chemotherapy in epithelial ovarian cancer. Cancer Cell Int. 2020, 20, 534. [Google Scholar] [CrossRef]
- Zhang, L.-Y.; Yu, J.-Y.; Leng, Y.-L.; Zhu, R.-R.; Liu, H.-X.; Wang, X.-Y.; Yang, T.-T.; Guo, Y.-N.; Tang, J.-L.; Zhang, X.-C. MiR-181c sensitizes ovarian cancer cells to paclitaxel by targeting GRP78 through the PI3K/Akt pathway. Cancer Gene Ther. 2022, 29, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-Y.; Li, P.-L.; Xu, A.; Zhang, X.-C. Involvement of GRP78 in the resistance of ovarian carcinoma cells to paclitaxel. Asian Pac. J. Cancer Prev. 2015, 16, 3517–3522. [Google Scholar] [CrossRef]


{kind=link}
| Cancer Type | Biomarker | Reference |
|---|---|---|
| Breast | ER | [20] |
| PgR | [20] | |
| HER2 | [20] | |
| Ki-67 | [10] | |
| BRCA1/2 mutation | [15] | |
| Colorectal | CEA | [14] |
| Bladder | BTA | [21] |
| NMP22 | [21] | |
| BladderCheck Test | [21] | |
| Lung | EarlyCDT-Lung | [22] |
| CEA | [22] | |
| CYFRA21-1 | [22] | |
| ProGRP | [22] | |
| Prostate | PSA | [14] |
| Ovarian | CA125 | [1] |
| BRCA1/2 mutation | [15] | |
| HE4 | [1] |
| Year | Tissue/Cell Type | N | miRNA | Reference |
|---|---|---|---|---|
| 2021 | SW626, SKOV3, OVACAR3, PA1 | N/A | miR-101 | Wei et al. [47] |
| 2020 | A2780cp, A2780s, SKOV3, CAOV3 | N/A | miR-193b | Li et al. [51] |
| 2020 | Tumour & TAN | 45 | miR-27b-5p | Liu et al. [50] |
| 2020 | Tumour & TAN | 31 | miR-584 | Yang et al. [49] |
| 2020 | SKOV3 | N/A | miR-155-5p | Ysrafil et al. [52] |
| 2018 | Blood, OVCAR3 | 8 | miR-195 | Chen [53] |
| 2016 | Tumour, SKOV3, OVCAR-429 | 20 | miR-125a, miR-125b | Lee et al. [54] |
| 2019 | Tumour & TAN, serum, OVAR3, HEY-A8 | 17 | let-7g | Biamonte et al. [55] |
| 2021 | Tumour & TAN, SKOV3, A2780 | 10 | miR-585-3p | Lu et al. [56] |
| 2021 | Serum | 30 | miR-193a-5p | Zhang et al. [57] |
| Year | Tissue/Cell Type | N | miRNA | Reference |
|---|---|---|---|---|
| 2021 | Plasma EVs | 34 | miR-4732-5p | Liu et al. [71] |
| 2020 | Tumour & TAN, CAOV3, SKOV3 | 11 | miR-141, miR-200a, miR-7, miR-203, miR-18a, miR-93, miR-106a, miR-20a, miR-19a | Wahab et al. [72] |
| 2019 | Serum EVs | 48 | miR-93, miR-145, miR-200c | Kim et al. [73] |
| 2017 | Tumour & TAN, A2780, OVCAR3, SKOV3, 3AO | 63 | miR-221 | Li et al. [66] |
| 2016 | Tumour & TAN, SKOV3, OV56, A2780, COV362, EFO21, OV90 | 145 | miR-760 | Liao et al. [74] |
| 2014 | Tumour & TAN | 487 | miR-96-5p, miR-182, miR-183, miR-141-5p, miR-200a, miR-200b, miR-200c, miR-429 | Wang et al. [70] |
| 2015 | Serum | 180 | miR-7, miR-429 | Meng et al. [69] |
| Study | Differential miRNA Expression Patterns |
|---|---|
| Hirschfeld et al. [75] |
|
| Hui et al. [76] |
|
| Agostini et al. [78] |
|
| Yanaihara et al. [79] |
|
| Zhang et al. [80] |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, M.; Davey, M.G.; Miller, N. The Potential of MicroRNAs as Clinical Biomarkers to Aid Ovarian Cancer Diagnosis and Treatment. Genes 2022, 13, 2054. https://doi.org/10.3390/genes13112054
Davies M, Davey MG, Miller N. The Potential of MicroRNAs as Clinical Biomarkers to Aid Ovarian Cancer Diagnosis and Treatment. Genes. 2022; 13(11):2054. https://doi.org/10.3390/genes13112054
Chicago/Turabian StyleDavies, Molly, Matthew G. Davey, and Nicola Miller. 2022. "The Potential of MicroRNAs as Clinical Biomarkers to Aid Ovarian Cancer Diagnosis and Treatment" Genes 13, no. 11: 2054. https://doi.org/10.3390/genes13112054