
A Novel Module Promotes Horizontal Gene Transfer in Azorhizobium caulinodans ORS571
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Growth Conditions
2.2. Bioinformatics Analyses
2.3. Molecular Biology Techniques
2.4. Measure HGT Frequency of ICEAc
2.5. Measuring Transcriptional and Translational Expression of Genes
2.6. Bacterial One-Hybrid Assay
2.7. Expression and Purification of His-Tagged RihR
2.8. Electrophoretic Mobility Gel Shift Assay (EMSA)
2.9. Quantitative Real-Time PCR Analysis
2.10. Statistical Analysis
3. Results
3.1. Identification of rgfs Involved in ICEAc Transfer
3.2. Genomic Context of rgf4 Genes in Different Strains
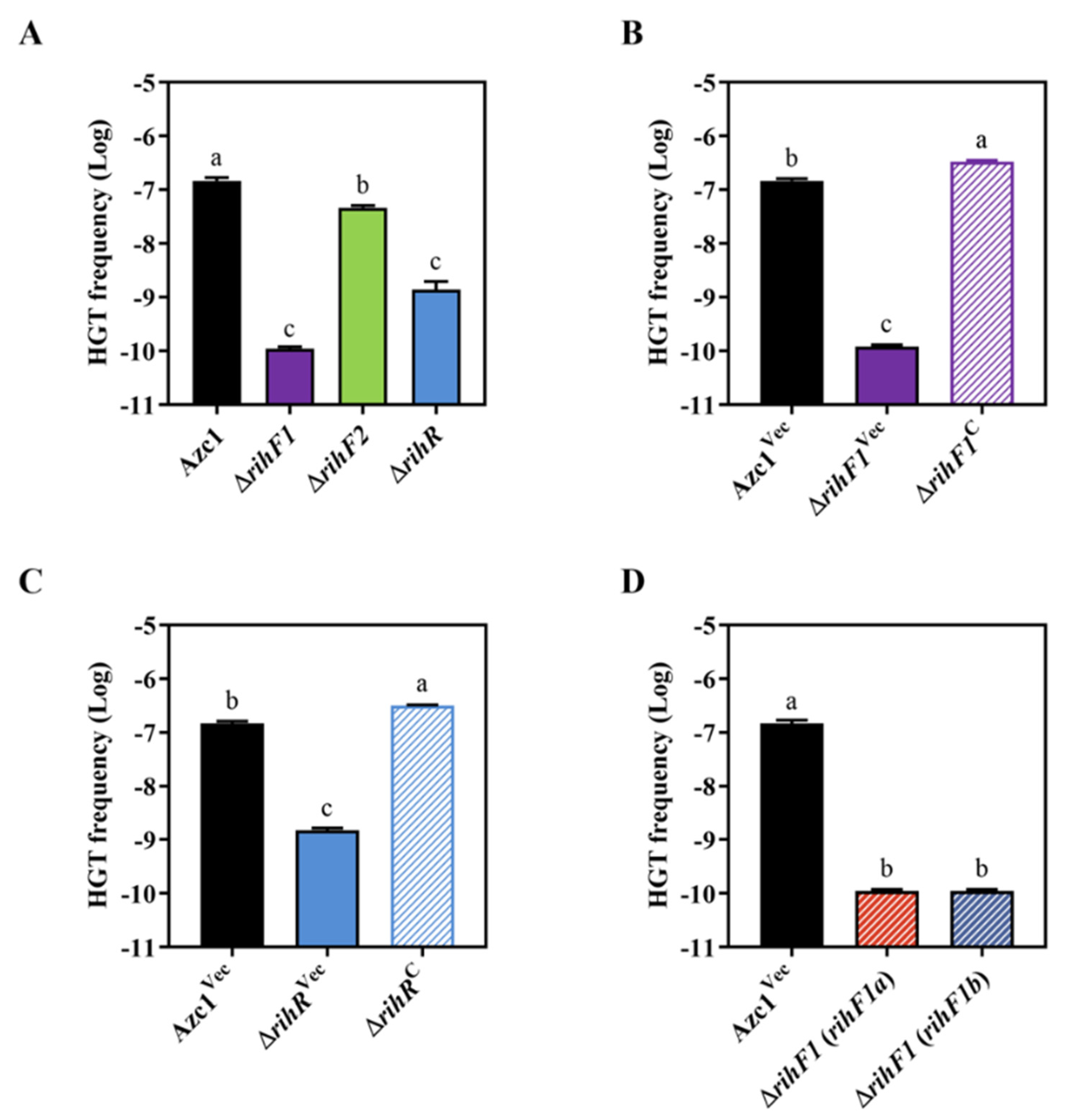
3.3. rihF1 and rihR Contribute to the Conjugation Frequency of ICEAc
3.4. ICEAc HGT Is Influenced by rihF1 and rihR as Two Indipendent Pathways
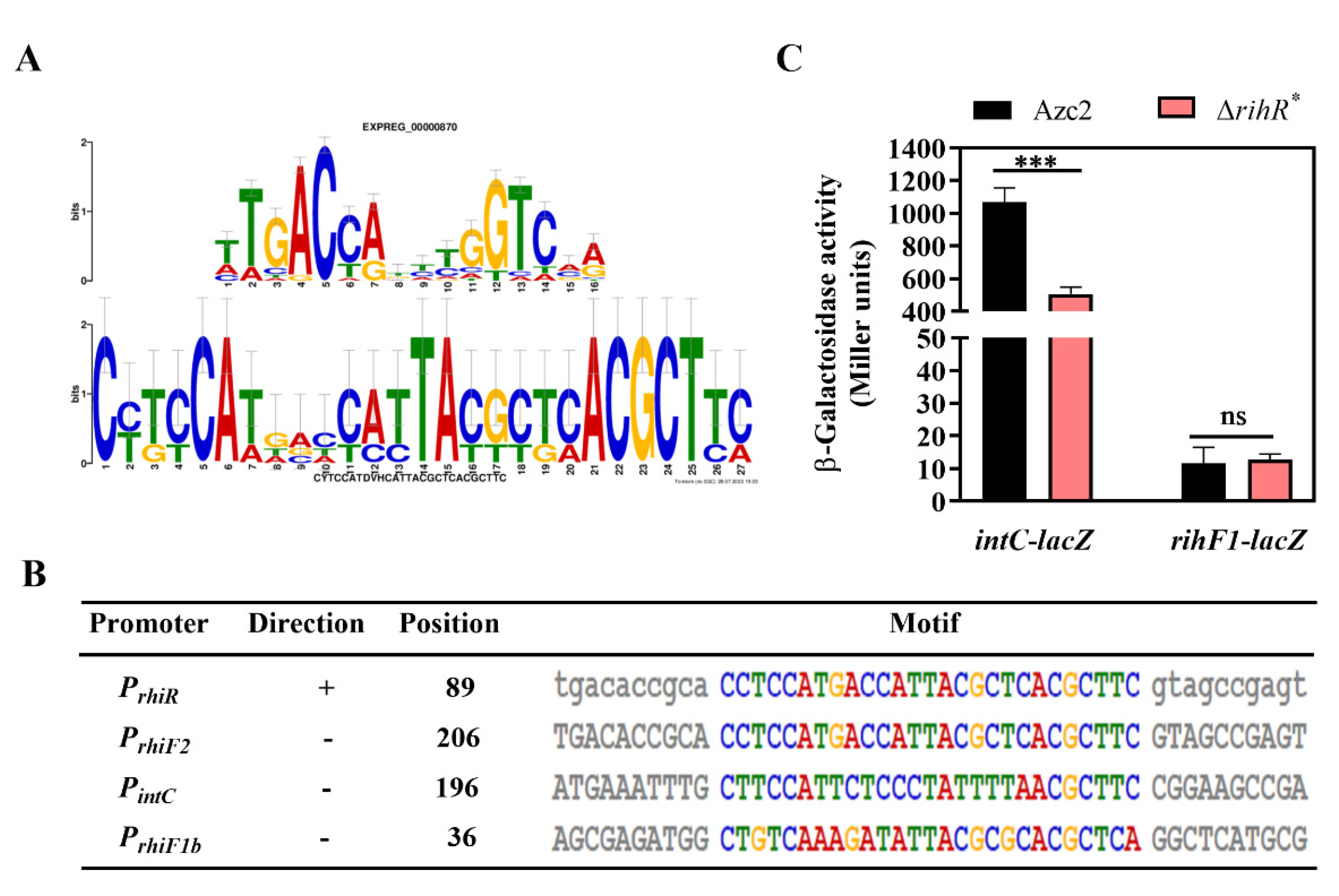
3.5. RihR Directly Binds to the Promoter Region of IntC
3.6. Regulation of rihR Is Independent of AhaR
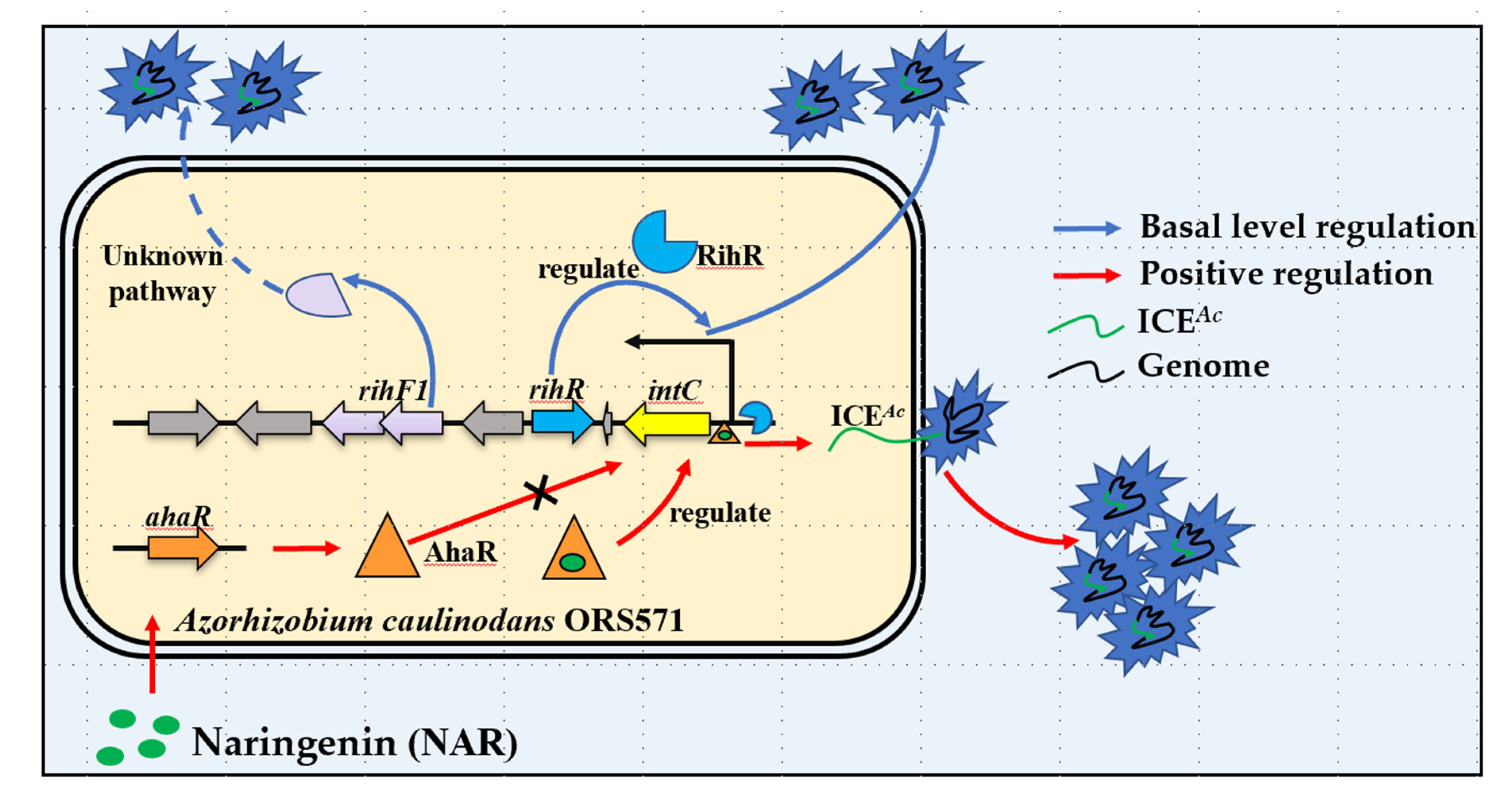
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heinemann, J.A.; Sprague, G.F., Jr. Bacterial conjugative plasmids mobilize DNA transfer between bacteria and yeast. Nature 1989, 340, 205–209. [Google Scholar] [CrossRef]
- Chen, I.; Dubnau, D. DNA uptake during bacterial transformation. Nat. Rev. Microbiol. 2004, 2, 241–249. [Google Scholar] [CrossRef]
- Goh, S. Phage Transduction. Methods Mol. Biol. 2016, 1476, 177–185. [Google Scholar]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot spots of horizontal gene transfer (HGT) in aquatic environments, with a focus on a new HGT mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Dubey, G.; Ben-Yehuda, S. Intercellular nanotubes mediate bacterial communication. Cell 2011, 144, 590–600. [Google Scholar] [CrossRef] [Green Version]
- Ochman, H.; Lawrence, J.G.; Groisman, E.A. Lateral gene transfer and the nature of bacterial innovation. Nature 2000, 405, 299–304. [Google Scholar] [CrossRef]
- Dagan, T.; Artzy-Randrup, Y.; Martin, W. Modular networks and cumulative impact of lateral transfer in prokaryote genome evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 10039–10044. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.K.; Meyer, T.E. Evolutionary analysis by whole-genome comparisons. J. Bacteriol. 2002, 184, 2260–2272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Ren, X.; Mason, A.S.; Liu, H.; Xiao, M.; Li, J.; Fu, D. Horizontal gene transfer in plants. Funct. Integr. Genomics 2014, 14, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Soucy, S.M.; Huang, J.; Gogarten, J. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef]
- Arnold, B.J.; Huang, I.T.; Hanage, W. Horizontal gene transfer and adaptive evolution in bacteria. Nat. Rev. Microbiol. 2022, 20, 206–218. [Google Scholar] [CrossRef]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of legumes by members of the beta-subclass of Proteobacteria. Nature 2001, 411, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Martinezromero, E. Recent Developments in Rhizobium Taxonomy. Plant Soil 1994, 161, 11–20. [Google Scholar] [CrossRef]
- Martínez, E.; Palacios, R.; Sánchez, F. Nitrogen-fixing nodules induced by Agrobacterium tumefaciens harboring Rhizobium phaseoli plasmids. J. Bacteriol. 1987, 169, 2828–2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geddes, B.A.; Kearsley, J.; Morton, R.; Finan, T.M. The Genomes of Rhizobia, in Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2020; pp. 213–249. [Google Scholar]
- Kelly, S.; Sullivan, J.; Ronson, C.; Tian, R.; Bräu, L.; Munk, C.; Goodwin, L.; Han, C.; Woyke, T.; Reddy, T.; et al. Genome sequence of the Lotu ssp. microsymbiont Mesorhizobium loti strain R7A. Stand. Genomic Sci. 2014, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.M.; Grossman, A.D. Integrative and Conjugative Elements (ICEs): What They Do and How They Work. Annu. Rev. Genet. 2015, 49, 577–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remigi, P.; Zhu, J.; Young, J.P.W.; Masson-Boivin, C. Symbiosis within Symbiosis: Evolving Nitrogen-Fixing Legume Symbionts. Trends Microbiol. 2016, 24, 63–75. [Google Scholar] [CrossRef]
- Sullivan, J.T.; Ronson, C.W. Evolution of rhizobia by acquisition of a 500-kb symbiosis island that integrates into a phe-tRNA gene. Proc. Natl. Acad. Sci. USA 1998, 95, 5145–5149. [Google Scholar] [CrossRef]
- Sullivan, J.T.; Patrick, H.N.; Lowther, W.L.; Scott, D.B.; Ronson, C.W. Nodulating strains of Rhizobium loti arise through chromosomal symbiotic gene transfer in the environment. Proc. Natl. Acad. Sci. USA 1995, 92, 8985–8989. [Google Scholar] [CrossRef] [Green Version]
- Marchetti, M.; Capela, D.; Glew, M.; Cruveiller, S.; Chane-Woon-Ming, B.; Gris, C.; Masson-Boivin, C. Experimental evolution of a plant pathogen into a legume symbiont. PLoS Biol. 2010, 8, e1000280. [Google Scholar] [CrossRef]
- Graham, P.H.; Vance, C. Legumes: Importance and constraints to greater use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef] [Green Version]
- Smil, V. Nitrogen in crop production: An account of global flows. Glob. Biogeochem. Cycles 1999, 13, 647–662. [Google Scholar] [CrossRef] [Green Version]
- Jackson, L.E.; Burger, M.; Cavagnaro, T.R. Roots, nitrogen transformations, and ecosystem services. Annu. Rev. Plant Biol. 2008, 59, 341–363. [Google Scholar] [CrossRef] [Green Version]
- Dreyfus, B.L.; Elmerich, C.; Dommergues, Y.R. Free-living Rhizobium strain able to grow on n(2) as the sole nitrogen source. Appl. Environ. Microbiol. 1983, 45, 711–713. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.T.; Trzebiatowski, J.R.; Cruickshank, R.W.; Gouzy, J.; Brown, S.D.; Elliot, R.M.; Ronson, C.W. Comparative sequence analysis of the symbiosis island of Mesorhizobium loti strain R7A. J. Bacteriol. 2002, 184, 3086–3095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, A.N.T.; Woods, L.C.; Gorrell, R.; Ramanan, S.; Kwok, T.; McDonald, M.J. Recombination resolves the cost of horizontal gene transfer in experimental populations of Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2022, 119, e2119010119. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.; Serbus, L.R. Gene Transfer Agents in Symbiotic Microbes. Results Probl. Cell Differ. 2020, 69, 25–76. [Google Scholar] [PubMed]
- Audette, G.F.; Manchak, J.; Beatty, P.; Klimke, W.A.; Frost, L.S. Entry exclusion in F-like plasmids requires intact TraG in the donor that recognizes its cognate TraS in the recipient. Microbiology 2007, 153 Pt 2, 442–451. [Google Scholar] [CrossRef]
- Geddes, B.A.; Kearsley, J.V.; Huang, J.; Zamani, M.; Muhammed, Z.; Sather, L.; Finan, T.M. Minimal gene set from Sinorhizobium (Ensifer) meliloti pSymA required for efficient symbiosis with Medicago. Proc. Natl. Acad. Sci. USA 2021, 118, e2018015118. [Google Scholar] [CrossRef] [PubMed]
- Unay, J.; Perret, X. A Minimal Genetic Passkey to Unlock Many Legume Doors to Root Nodulation by Rhizobia. Genes 2020, 11, 521. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Wang, H.; Wu, P.; Li, T.; Tang, Y.; Naseer, N.; Zhu, J. Plant nodulation inducers enhance horizontal gene transfer of Azorhizobium caulinodans symbiosis island. Proc. Natl. Acad. Sci. USA 2016, 113, 13875–13880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskett, T.L.; Paramasivan, P.; Mendes, M.D.; Green, P.; Geddes, B.A.; Knights, H.E.; Jorrin, B.; Ryu, M.; Brett, P.; Voigt, C.A.; et al. Engineered plant control of associative nitrogen fixation. Proc. Natl. Acad. Sci. USA 2022, 119, e2117465119. [Google Scholar] [CrossRef]
- Wathugala, N.D.; Hemananda, K.M.; Yip, C.B.; Hynes, M.F. Defining the requirements for the conjugative transfer of Rhizobium leguminosarum plasmid pRleVF39b. Microbiology 2020, 166, 318–331. [Google Scholar] [CrossRef]
- López-Fuentes, E.; Torres-Tejerizo, G.; Cervantes, L.; Brom, S. Genes encoding conserved hypothetical proteins localized in the conjugative transfer region of plasmid pRet42a from Rhizobium etli CFN42 participate in modulating transfer and affect conjugation from different donors. Front. Microbiol. 2014, 5, 793. [Google Scholar]
- Ding, H.; Yip, C.B.; Hynes, M.F. Genetic characterization of a novel rhizobial plasmid conjugation system in Rhizobium leguminosarum bv. viciae strain VF39SM. J. Bacteriol. 2013, 195, 328–339. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lv, S.; Liu, T.; Wei, J.; Qu, S.; Lu, Y.; Liu, H. CRISPR/Cas9 genome editing shows the important role of AZC_2928 gene in nitrogen-fixing bacteria of plants. Funct. Integr. Genomics 2020, 20, 657–668. [Google Scholar] [CrossRef]
- Pistorio, M.; Torres Tejerizo, G.A.; Del Papa, M.F.; de los Angeles Giusti, M.; Lozano, M.; Lagares, A. rptA, a novel gene from Ensifer (Sinorhizobium) meliloti involved in conjugal transfer. FEMS Microbiol. Lett. 2013, 345, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.; Guo, D.; Deng, S.; Lu, X.; Zhu, J.; Rao, B.; Zhu, J. Ohr and OhrR Are Critical for Organic Peroxide Resistance and Symbiosis in Azorhizobium caulinodans ORS571. Genes 2020, 11, 335. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids. Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Metcalf, W.W.; Jiang, W.; Daniels, L.L.; Kim, S.K.; Haldimann, A.; Wanner, B.L. Conditionally replicative and conjugative plasmids carrying lacZ alpha for cloning, mutagenesis, and allele replacement in bacteria. Plasmid 1996, 35, 1–13. [Google Scholar] [CrossRef]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Chart, H.; Smith, H.; La Ragione, R.; Woodward, M. An investigation into the pathogenic properties of Escherichia coli strains BLR, BL21, DH5alpha and EQ1. J. Appl. Microbiol. 2000, 89, 1048–1058. [Google Scholar] [CrossRef]
- Ferrières, L.; Hémery, G.; Nham, T.; Gueérout, A.-M.; Mazel, D.; Beloin, C.; Ghigo, J.-M. Silent Mischief: Bacteriophage Mu Insertions Contaminate Products of Escherichia coli Random Mutagenesis Performed Using Suicidal Transposon Delivery Plasmids Mobilized by Broad-Host-Range RP4 Conjugative Machinery. J. Bacteriol. 2010, 192, 6418–6427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goethals, K.; Van Montagu, M.; Holsters, M. Conserved motifs in a divergent nod box of Azorhizobium caulinodans ORS571 reveal a common structure in promoters regulated by LysR-type proteins. Proc. Natl. Acad. Sci. USA 1992, 89, 1646–1650. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Nickels, L.M.; Wang, H.; Ling, J.; Zhong, Z.; Zhu, J. OxyR-regulated catalase activity is critical for oxidative stress resistance, nodulation and nitrogen fixation in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2016, 363, fnw130. [Google Scholar] [CrossRef] [Green Version]
- Miller, J. Experiments in Molecular Genetics 1972 Cold Spring Harbor; Cold Spring Harbor Laboratory: Laurel Hollow, NY, USA, 1972; pp. 431–433. [Google Scholar]
- Kalogeraki, V.S.; Winans, S.C. Suicide plasmids containing promoterless reporter genes can simultaneously disrupt and create fusions to target genes of diverse bacteria. Gene 1997, 188, 69–75. [Google Scholar] [CrossRef]
- Jiang, G.; Yang, J.; Li, X.; Cao, Y.; Liu, X.; Ling, J.; Wang, H.; Zhong, Z.; Zhu, J. Alkyl hydroperoxide reductase is important for oxidative stress resistance and symbiosis in Azorhizobium caulinodans. FEMS Microbiol. Lett. 2019, 366, fnz014. [Google Scholar] [CrossRef]
- Guo, M.; Feng, H.; Zhang, J.; Wang, W.; Wang, Y.; Li, Y.; Gao, C.; Chen, H.; Feng, Y.; He, Z.-G. Dissecting transcription regulatory pathways through a new bacterial one-hybrid reporter system. Genome Res. 2009, 19, 1301–1308. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Chen, H.; Qian, G.; Liu, F. LetR is a TetR family transcription factor from Lysobacter controlling antifungal antibiotic biosynthesis. Appl. Microbiol. Biotechnol. 2017, 101, 3273–3282. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Shen, D.; Yang, N.; Chou, S.H.; Gomelsky, M.; Qian, G. Coordinated control of the type IV pili and c-di-GMP-dependent antifungal antibiotic production in Lysobacter by the response regulator PilR. Mol. Plant Pathol. 2021, 22, 602–617. [Google Scholar] [CrossRef]
- Allemann, M.N.; Allen, E.E. Genetic Regulation of the Bacterial Omega-3 Polyunsaturated Fatty Acid Biosynthesis Pathway. J. Bacteriol. 2020, 202, e00050-20. [Google Scholar] [CrossRef] [PubMed]
- Guan, G.; Dai, P.-H.; Osborne, T.F.; Kim, J.B.; Shechter, I. Multiple Sequence Elements are Involved in the Transcriptional Regulation of the Human Squalene Synthase Gene. J. Biol. Chem. 1997, 272, 10295–10302. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Hao, G.; Li, Z.; Zhou, Y.; Garcia-Sillas, R.; Li, J.; Wang, H.; Kan, B.; Zhu, J. CitAB Two-Component System-Regulated Citrate Utilization Contributes to Vibrio cholerae Competitiveness with the Gut Microbiota. Infect. Immun. 2019, 87, e00746-18. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Zhang, X.; Zhang, Y.; Zhu, M.; Yang, P.; Yuan, J.; Zhou, Y.X.T.; Wang, W.; Chen, S.; Liang, H.; et al. RpoN-Dependent Direct Regulation of Quorum Sensing and the Type VI Secretion System in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2018, 200, e00205-18. [Google Scholar] [CrossRef] [Green Version]
- Jahn, C.E.; Charkowski, A.O.; Willis, D.K. Evaluation of isolation methods and RNA integrity for bacterial RNA quantitation. J. Microbiol. Methods 2008, 75, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ゔゔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.L.; Wood, T.K.; Peti, W. Rebecca Page Structure of the Escherichia coli antitoxin MqsA (YgiT/b3021) bound to its gene promoter reveals extensive domain rearrangements and the specificity of transcriptional regulation. J. Biol. Chem. 2011, 286, 2285–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, A.; Tamman, H.; Ainelo, A.; Konijnenberg, A.; Hadži, S.; Sobott, F.; Garcia-Pino, A.; Hõrak, R.; Loris, R. A dual role in regulation and toxicity for the disordered N-terminus of the toxin GraT. Nat. Commun. 2019, 10, 972. [Google Scholar] [CrossRef]
- Wang, H.C.; Ko, T.; Wu, M.; Ku, S.; Wu, H.; Wang, A. Neisseria conserved protein DMP19 is a DNA mimic protein that prevents DNA binding to a hypothetical nitrogen-response transcription factor. Nucleic Acids Res. 2012, 40, 5718–5730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, J.P.; Major, A.S.; Komarovsky, V.M.; Sullivan, J.T.; Dy, R.L.; Hynes, M.F.; Salmond, G.P.C.; Ronson, C.W. A widely conserved molecular switch controls quorum sensing and symbiosis island transfer in Mesorhizobium loti through expression of a novel antiactivator. Mol. Microbiol. 2013, 87, 1–13. [Google Scholar] [CrossRef]
- Ramsay, J.P.; Tester, L.G.L.; Major, A.S.; Sullivan, J.T.; Edgar, C.D.; Kleffmann, T.; Patterson-House, J.R.; Hall, D.A.; Tate, W.P.; Hynes, M.F.; et al. Ribosomal frameshifting and dual-target antiactivation restrict quorum-sensing-activated transfer of a mobile genetic element. Proc. Natl. Acad. Sci. USA 2015, 112, 4104–4109. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Minamisawa, K.; Uchiumi, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Iriguchi, M.; Kawashima, K.; et al. Complete genomic sequence of nitrogen-fixing symbiotic bacterium Bradyrhizobium japonicum USDA110. DNA Res. 2002, 9, 189–197. [Google Scholar] [CrossRef]
- Shimada, T.; Hirao, K.; Kori, A.; Yamamoto, K.; Ishihama, A. RutR is the uracil/thymine-sensing master regulator of a set of genes for synthesis and degradation of pyrimidines. Mol. Microbiol. 2007, 66, 744–757. [Google Scholar] [CrossRef]
- Loh, K.D.; Hirao, K.; Kori, A.; Yamamoto, K.; Ishihama, A. A previously undescribed pathway for pyrimidine catabolism. Proc. Natl. Acad. Sci. USA 2006, 103, 5114–5119. [Google Scholar] [CrossRef] [Green Version]
- Gogarten, J.; Townsend, J. Horizontal gene transfer, genome innovation and evolution. Nat. Rev. Microbiol. 2005, 3, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, J.; Chen, L.; Wang, J.; Cheng, Q.; Dixon, R.; Wang, Y. Using synthetic biology to distinguish and overcome regulatory and functional barriers related to nitrogen fixation. PLoS ONE 2013, 8, e68677. [Google Scholar] [CrossRef] [Green Version]
- McCloskey, D.; Xu, S.; Sandberg, T.E.; Brunk, E.; Hefner, Y.; Szubin, R.; Feist, A.M. Palsson Evolution of gene knockout strains of E. coli reveal regulatory architectures governed by metabolism. Nat. Commun. 2018, 9, 3796. [Google Scholar] [CrossRef] [Green Version]
- Ryu, M.H.; Zhang, J.; Khokhani, T.T.D.; Geddes, B.A.; Mus, F.; Garcia-Costas, A.; Peters, J.W.; Poole, P.S.; Ané, J. Control of nitrogen fixation in bacteria that associate with cereals. Nat. Microbiol. 2020, 5, 314–330. [Google Scholar] [CrossRef]
- Doin de Moura, G.G.; Remigi, P.; Masson-Boivin, C. Delphine Capela Experimental Evolution of Legume Symbionts: What Have We Learnt? Genes 2020, 11, 339. [Google Scholar] [CrossRef] [Green Version]
- Inomura, K.; Bragg, J.; Follows, M.J. A quantitative analysis of the direct and indirect costs of nitrogen fixation: A model based on Azotobacter vinelandii. ISME J. 2017, 11, 166–175. [Google Scholar] [CrossRef]
- Taylor, B.N.; Menge, D.N.L. Light, nitrogen supply, and neighboring plants dictate costs and benefits of nitrogen fixation for seedlings of a tropical nitrogen-fixing tree. New Phytol. 2021, 231, 1758–1769. [Google Scholar] [CrossRef]
- Bañuelos-Vazquez, L.A.; Tejerizo, G.T.; la Luz, L.C.; Girard, L.; Romero, D.; Brom, S. Conjugative transfer between Rhizobium etli endosymbionts inside the root nodule. Environ. Microbiol. 2019, 21, 3430–3441. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, K.; Liu, Y.; Zou, D.; Wang, D.; Xie, Z. Effects of Calcium and Signal Sensing Systems on Azorhizobium caulinodans Biofilm Formation and Host Colonization. Front. Microbiol. 2020, 11, 563367. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Li, Y.; Bai, X.; Wu, H.; Bian, L.; Hu, X. LuxR-Type Regulator AclR1 of Azorhizobium caulinodans Regulates Cyclic di-GMP and Numerous Phenotypes in Free-Living and Symbiotic States. Mol. Plant. Microbe Interact. 2020, 33, 528–538. [Google Scholar] [CrossRef]
- Mølbak, L.; Molin, S.; Kroer, N. Root growth and exudate production define the frequency of horizontal plasmid transfer in the Rhizosphere. FEMS Microbiol. Ecol. 2007, 59, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Kroer, N.; Barkay, T.; Sørensen, S.; Weber, D. Effect of root exudates and bacterial metabolic activity on conjugal gene transfer in the rhizosphere of a marsh plant. FEMS Microbiol. Ecol. 1998, 25, 375–384. [Google Scholar] [CrossRef]
- Rashid, M.M.; Ikawa, Y.; Tsuge, S. GamR, the LysR-Type Galactose Metabolism Regulator, Regulates hrp Gene Expression via Transcriptional Activation of Two Key hrp Regulators, HrpG and HrpX, in Xanthomonas oryzae pv. oryzae. Appl. Environ. Microbiol. 2016, 82, 3947–3958. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Chen, Q.; Wu, C.; Li, Y.; Wang, S.; Chen, X.; Qiu, B.; Li, Y.; Mao, D.; Lin, H.; et al. A Novel Module Promotes Horizontal Gene Transfer in Azorhizobium caulinodans ORS571. Genes 2022, 13, 1895. https://doi.org/10.3390/genes13101895
Li M, Chen Q, Wu C, Li Y, Wang S, Chen X, Qiu B, Li Y, Mao D, Lin H, et al. A Novel Module Promotes Horizontal Gene Transfer in Azorhizobium caulinodans ORS571. Genes. 2022; 13(10):1895. https://doi.org/10.3390/genes13101895
Chicago/Turabian StyleLi, Mingxu, Qianqian Chen, Chuanhui Wu, Yiyang Li, Sanle Wang, Xuelian Chen, Bowen Qiu, Yuxin Li, Dongmei Mao, Hong Lin, and et al. 2022. "A Novel Module Promotes Horizontal Gene Transfer in Azorhizobium caulinodans ORS571" Genes 13, no. 10: 1895. https://doi.org/10.3390/genes13101895