
The Genetic Factors of the Airway Epithelium Associated with the Pathology of Asthma
 ,
,
Abstract
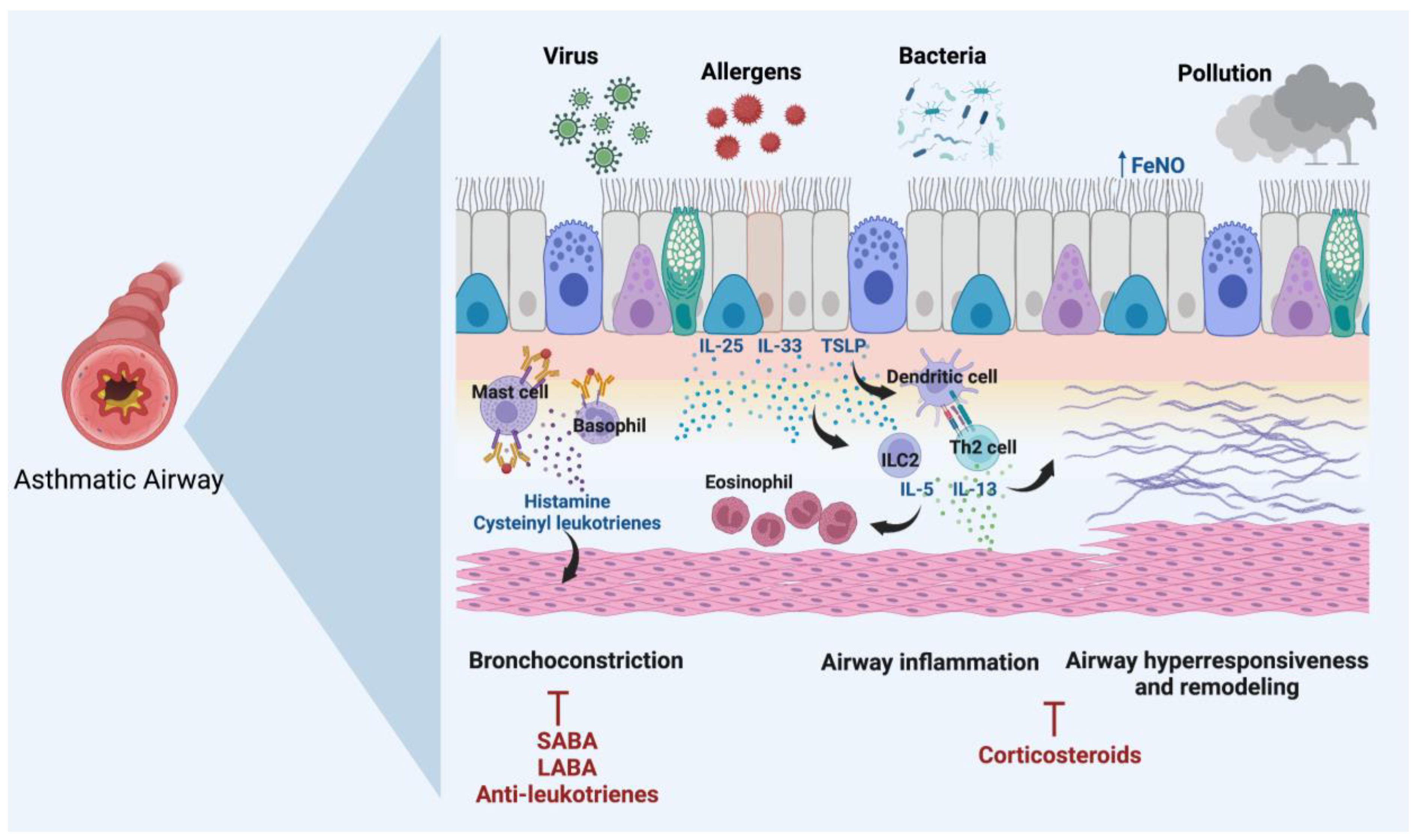
:1. Introduction
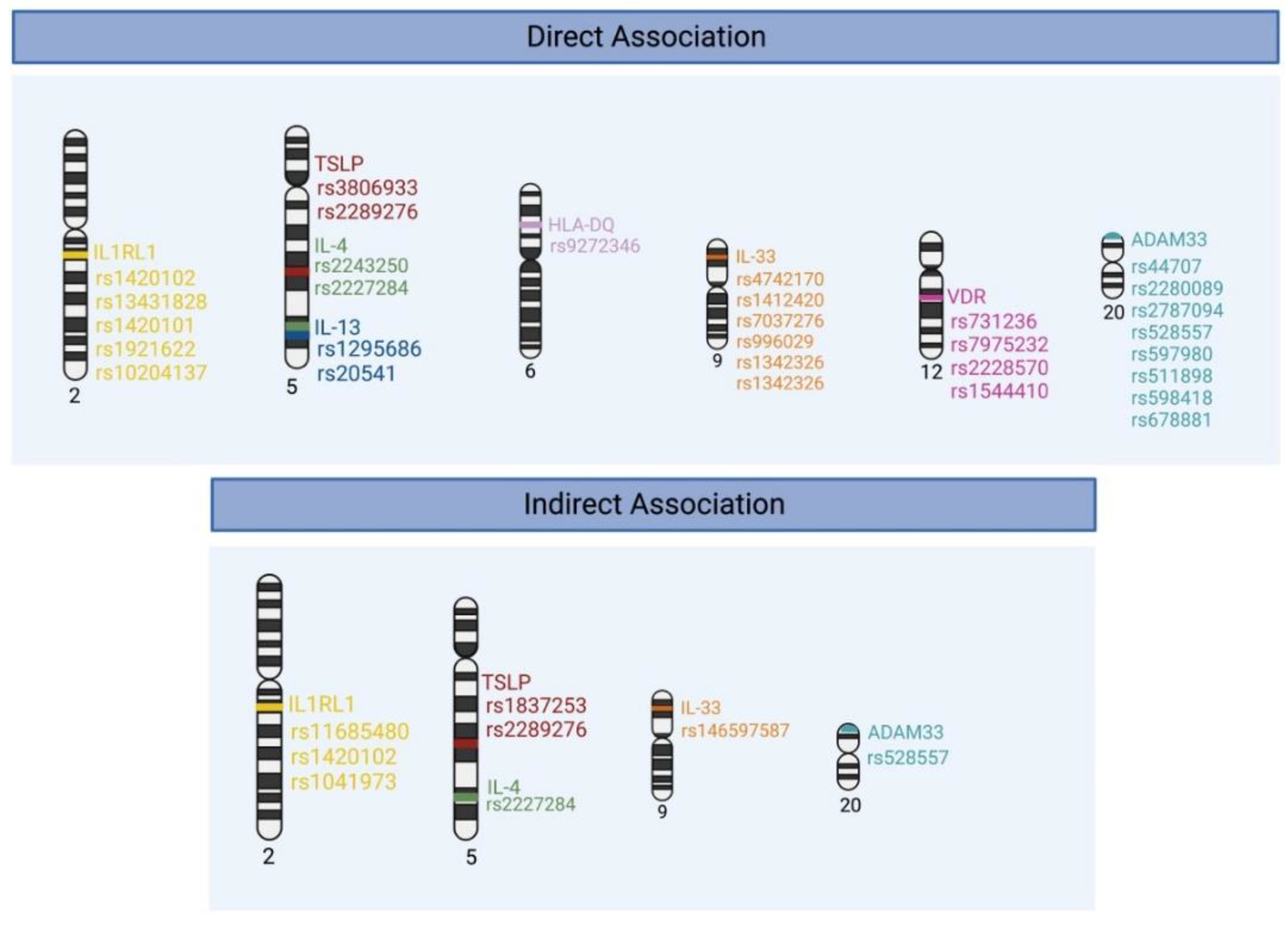
2. Genetic Polymorphisms Associated with Asthma Susceptibility and Severity
2.1. Thymic Stromal Lymphopoietin (TSLP)
2.2. Interleukin-33 (IL-33)
2.3. Interleukin 1 Receptor-like 1 (IL1RL1)
2.4. Interleukin-13 (IL-13)
2.5. Interleukin-4 (IL-4)
2.6. The Human Leukocyte Antigen (HLA)
2.7. A Disintegrin and Metalloproteinase 33 (ADAM33)
2.8. Vitamin D Receptor (VDR)
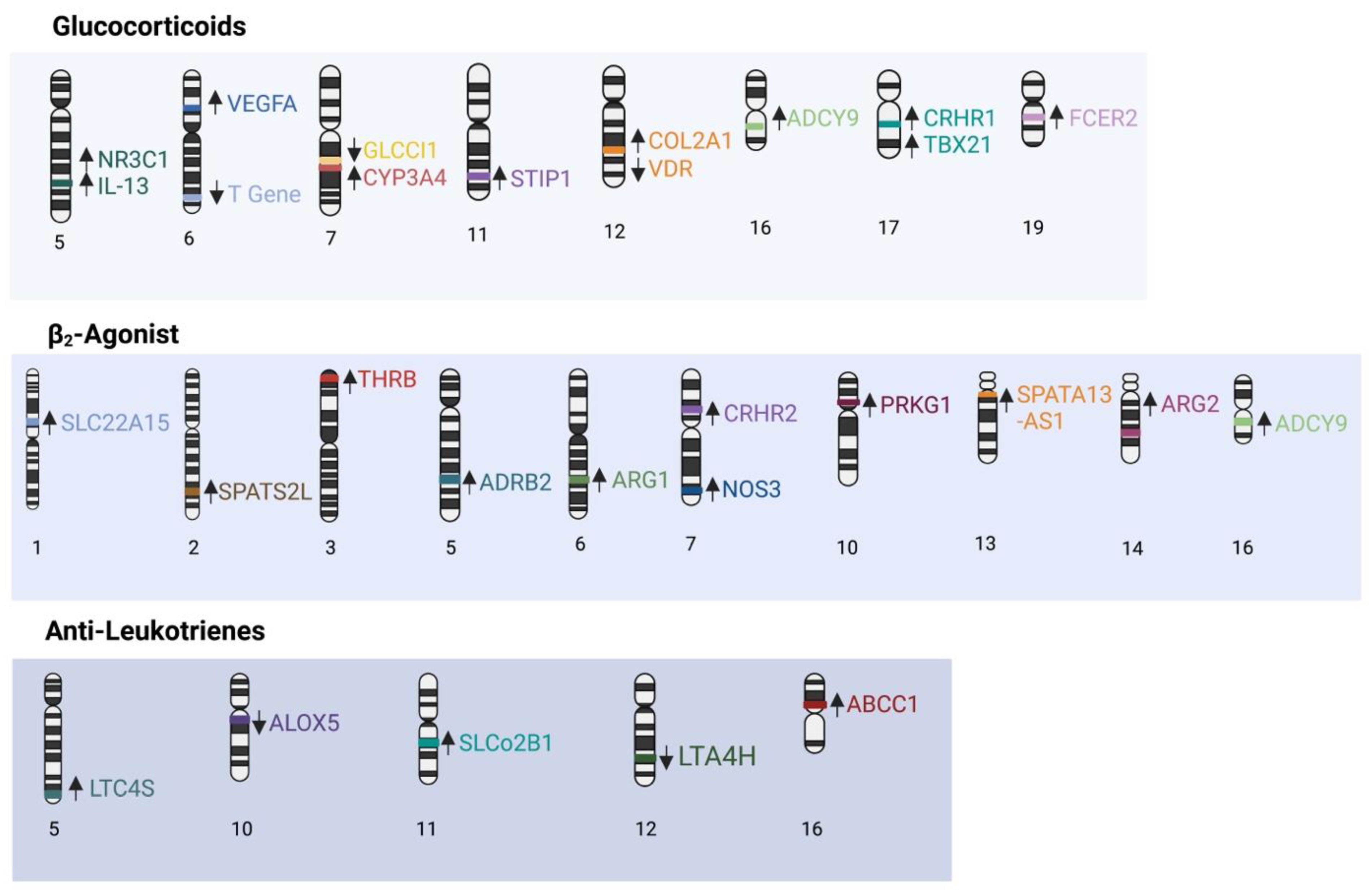
3. Genetic Polymorphisms Associated with Asthma Treatment Response
3.1. Corticosteroids
3.2. β-2 Adrenergic Receptor Pathway
3.3. Leukotrienes
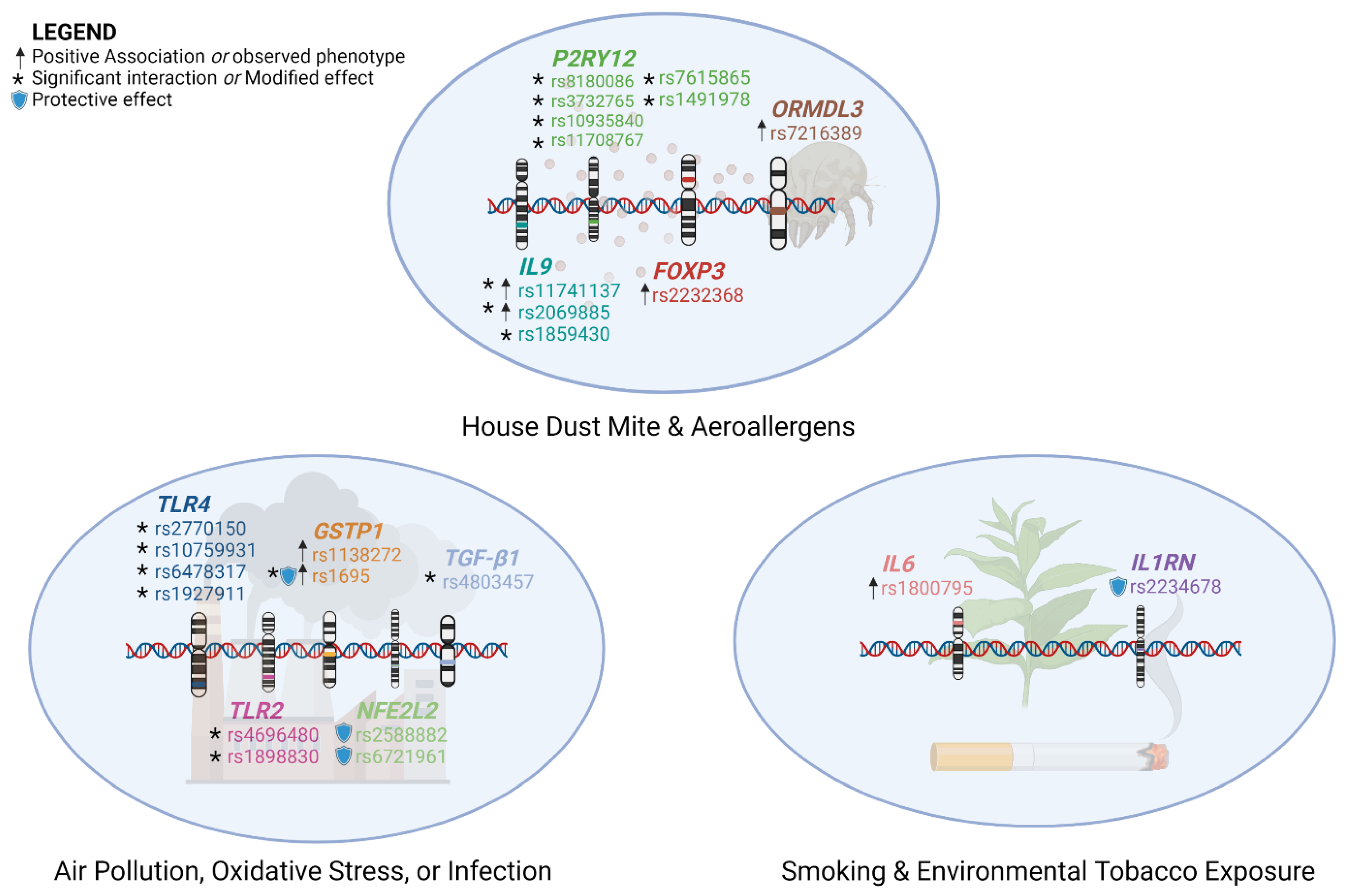
4. Interactions between Genetic Polymorphisms and Environmental Factors
4.1. Air Pollution
4.2. Aeroallergens
4.3. Oxidative Stress
4.4. Smoking
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Quirt, J.; Hildebrand, K.J.; Mazza, J.; Noya, F.; Kim, H. Asthma. Allergy Asthma Clin. Immunol. 2018, 14, 50. [Google Scholar] [CrossRef]
- Boulet, L.-P. Influence of comorbid conditions on asthma. Eur. Respir. J. 2009, 33, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Dharmage, S.C.; Perret, J.L.; Custovic, A. Epidemiology of Asthma in Children and Adults. Front. Pediatr. 2019, 7, 246. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, S.F.; Ulrik, C.S.; Kyvik, K.O.; Ferreira, M.A.R.; Backer, V. Multivariate genetic analysis of atopy phenotypes in a selected sample of twins. Clin. Exp. Allergy 2006, 36, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Asthma mechanisms. Medicine 2016, 44, 265–270. [Google Scholar] [CrossRef]
- Dunican, E.M.; Fahy, J.V. The Role of Type 2 Inflammation in the Pathogenesis of Asthma Exacerbations. Ann. Am. Thorac. Soc. 2015, 12 (Suppl. 2), S144–S149. [Google Scholar] [CrossRef]
- Shen, L.X.; Basilion, J.P.; Stanton, V.P., Jr. Single-nucleotide polymorphisms can cause different structural folds of mRNA. Proc. Natl. Acad. Sci. USA 1999, 96, 7871–7876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, V.; Patel, N.; Turcotte, M.; Bossé, Y.; Paré, G.; Meyre, D. Benefits and limitations of genome-wide association studies. Nat. Rev. Genet. 2019, 20, 467–484. [Google Scholar] [CrossRef] [PubMed]
- Demehri, S.; Morimoto, M.; Holtzman, M.J.; Kopan, R. Skin-Derived TSLP Triggers Progression from Epidermal-Barrier Defects to Asthma. PLOS Biol. 2009, 7, e1000067. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; O’Connor, B.; Ratoff, J.; Meng, Q.; Mallett, K.; Cousins, D.; Robinson, D.; Zhang, G.; Zhao, J.; Lee, T.H.; et al. Thymic Stromal Lymphopoietin Expression Is Increased in Asthmatic Airways and Correlates with Expression of Th2-Attracting Chemokines and Disease Severity. J. Immunol. 2005, 174, 8183–8190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauvreau, G.M.; O’Byrne, P.M.; Boulet, L.-P.; Wang, Y.; Cockcroft, D.; Bigler, J.; FitzGerald, J.M.; Boedigheimer, M.; Davis, B.E.; Dias, C.; et al. Effects of an Anti-TSLP Antibody on Allergen-Induced Asthmatic Responses. N. Engl. J. Med. 2014, 370, 2102–2110. [Google Scholar] [CrossRef] [PubMed]
- Menzies-Gow, A.; Corren, J.; Bourdin, A.; Chupp, G.; Israel, E.; Wechsler, M.E.; Brightling, C.E.; Griffiths, J.M.; Hellqvist, Å.; Bowen, K.; et al. Tezepelumab in Adults and Adolescents with Severe, Uncontrolled Asthma. N. Engl. J. Med. 2021, 384, 1800–1809. [Google Scholar] [CrossRef] [PubMed]
- Harada, M.; Hirota, T.; Jodo, A.I.; Hitomi, Y.; Sakashita, M.; Tsunoda, T.; Miyagawa, T.; Doi, S.; Kameda, M.; Fujita, K.; et al. Thymic Stromal Lymphopoietin Gene Promoter Polymorphisms Are Associated with Susceptibility to Bronchial Asthma. Am. J. Respir. Cell Mol. Biol. 2011, 44, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Karaaslan, C.; Yavuz, T.S.; Cosgun, E.; Kalayci, O.; Sackesen, C. The Genetic Variants of Thymic Stromal Lymphopoietin Protein in Children with Asthma and Allergic Rhinitis. Int. Arch. Allergy Immunol. 2014, 163, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Hunninghake, G.M.; Soto-Quirós, M.E.; Avila, L.; Kim, H.P.; Lasky-Su, J.; Rafaels, N.; Ruczinski, I.; Beaty, T.H.; Mathias, R.A.; Barnes, K.C.; et al. TSLP polymorphisms are associated with asthma in a sex-specific fashion. Allergy 2010, 65, 1566–1575. [Google Scholar] [CrossRef] [Green Version]
- Moorehead, A.; Hanna, R.; Heroux, D.; Neighbour, H.; Sandford, A.; Gauvreau, G.M.; Sommer, D.D.; Denburg, J.A.; Akhabir, L. A thymic stromal lymphopoietin polymorphism may provide protection from asthma by altering gene expression. Clin. Exp. Allergy 2020, 50, 471–478. [Google Scholar] [CrossRef]
- Ranjbar, M.; Matloubi, M.; Assarehzadegan, M.-A.; Fallahpour, M.; Sadeghi, F.; Soleyman-Jahi, S.; Janani, L. Association between Two Single Nucleotide Polymorphisms of Thymic Stromal Lymphopoietin (TSLP) Gene and Asthma in Iranian Population. Iran. J. Allergy Asthma Immunol. 2020, 19, 362–372. [Google Scholar] [CrossRef]
- Chan, B.C.L.; Lam, C.W.K.; Tam, L.-S.; Wong, C.K. IL33: Roles in Allergic Inflammation and Therapeutic Perspectives. Front. Immunol. 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Liew, F.Y.; Girard, J.-P.; Turnquist, H.R. Interleukin-33 in health and disease. Nat. Rev. Immunol. 2016, 16, 676–689. [Google Scholar] [CrossRef]
- Préfontaine, D.; Lajoie-Kadoch, S.; Foley, S.; Audusseau, S.; Olivenstein, R.; Halayko, A.J.; Lemière, C.; Martin, J.G.; Hamid, Q. Increased Expression of IL-33 in Severe Asthma: Evidence of Expression by Airway Smooth Muscle Cells. J. Immunol. 2009, 183, 5094–5103. [Google Scholar] [CrossRef]
- Moffatt, M.F.; Gut, I.G.; Demenais, F.; Strachan, D.P.; Bouzigon, E.; Heath, S.; von Mutius, E.; Farrall, M.; Lathrop, M.; Cookson, W.O.C.M. A Large-Scale, Consortium-Based Genomewide Association Study of Asthma. N. Engl. J. Med. 2010, 363, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.; Zheng, X.; Huang, J.; Hu, X. Association of IL33, IL1RL1, IL1RAP Polymorphisms and Asthma in Chinese Han Children. Front. Cell Dev. Biol. 2021, 9, 759542. [Google Scholar] [CrossRef] [PubMed]
- Lajunen, T.K.; Jaakkola, J.J.K.; Jaakkola, M.S. Interleukin 33(IL33) polymorphisms associate with incident adult-onset asthma. Eur. Respir. J. 2015, 46, OA1457. [Google Scholar] [CrossRef]
- Matloubi, M.; Ranjbar, M.; Assarehzadegan, M.-A.; Fallahpour, M.; Sadeghi, F.; Soleyman-Jahi, S.; Janani, L. The Impact of Interleukin (IL)-33 Gene Polymorphisms and Environmental Factors on Risk of Asthma in the Iranian Population. Lung 2020, 198, 105–112. [Google Scholar] [CrossRef]
- Smith, D.; Helgason, H.; Sulem, P.; Bjornsdottir, U.S.; Lim, A.C.; Sveinbjornsson, G.; Hasegawa, H.; Brown, M.; Ketchem, R.R.; Gavala, M.; et al. A rare IL33 loss-of-function mutation reduces blood eosinophil counts and protects from asthma. PLoS Genet. 2017, 13, e1006659. [Google Scholar] [CrossRef]
- Queiroz, G.A.; Costa, R.S.; Alcantara-Neves, N.M.; Nunes de Oliveira Costa, G.; Barreto, M.L.; Carneiro, V.L.; Figueiredo, C.A. IL33 and IL1RL1 variants are associated with asthma and atopy in a Brazilian population. Int. J. Immunogenet. 2017, 44, 51–61. [Google Scholar] [CrossRef]
- Griesenauer, B.; Paczesny, S. The ST2/IL-33 Axis in Immune Cells during Inflammatory Diseases. Front. Immunol. 2017, 8, 475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Lu, J.; Yin, X.; Sun, Y.; Li, S. Association of IL-33, IL1RL1 gene polymorphisms with serum IL-33 levels and risk of asthma in adults and asthmatic bronchitis in children (Chinese). Biotechnol. Biotechnol. Equip. 2018, 32, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Mendez, J.A.; Nazario, S.; Laureano, A.; Baez, A.; Rivera, B. IL-33 and IL1RL1 Single Nucleotide Polymorphisms and Their Association with Asthma Among Puerto Ricans. J. Allergy Clin. Immunol. 2014, 133, AB67. [Google Scholar] [CrossRef]
- Savenije, O.E.M.; Kerkhof, M.; Reijmerink, N.E.; Brunekreef, B.; de Jongste, J.C.; Smit, H.A.; Wijga, A.H.; Postma, D.S.; Koppelman, G.H. Interleukin-1 receptor–like 1 polymorphisms are associated with serum IL1RL1-a, eosinophils, and asthma in childhood. J. Allergy Clin. Immunol. 2011, 127, 750–756.e5. [Google Scholar] [CrossRef]
- Dijk, F.N.; Xu, C.; Melén, E.; Carsin, A.-E.; Kumar, A.; Nolte, I.M.; Gruzieva, O.; Pershagen, G.; Grotenboer, N.S.; Savenije, O.E.M.; et al. Genetic regulation of IL1RL1 methylation and IL1RL1-a protein levels in asthma. Eur. Respir. J. 2018, 51, 1701377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijk, F.N.; Vijverberg, S.J.; Hernandez-Pacheco, N.; Repnik, K.; Karimi, L.; Mitratza, M.; Farzan, N.; Nawijn, M.C.; Burchard, E.G.; Engelkes, M.; et al. IL1RL1 gene variations are associated with asthma exacerbations in children and adolescents using inhaled corticosteroids. Allergy 2020, 75, 984–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A. IL-13 Effector Functions. Annu. Rev. Immunol. 2003, 21, 425–456. [Google Scholar] [CrossRef]
- Granada, M.; Wilk, J.B.; Tuzova, M.; Strachan, D.P.; Weidinger, S.; Albrecht, E.; Gieger, C.; Heinrich, J.; Himes, B.E.; Hunninghake, G.M.; et al. A genome-wide association study of plasma total IgE concentrations in the Framingham Heart Study. J. Allergy Clin. Immunol. 2012, 129, 840–845.e21. [Google Scholar] [CrossRef] [Green Version]
- Halwani, R.; Vazquez-Tello, A.; Kenana, R.; Al-Otaibi, M.; Alhasan, K.A.; Shakoor, Z.; Al-Muhsen, S. Association of IL-13 rs20541 and rs1295686 variants with symptomatic asthma in a Saudi Arabian population. J. Asthma 2018, 55, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.M.; Howson, J.M.; Walker, N.; Spickett, G.P.; Jones, R.W.; Ring, S.M.; McArdle, W.L.; Lowe, C.E.; Bailey, R.; Payne, F. Association of IL13 with total IgE: Evidence against an inverse association of atopy and diabetes. J. Allergy Clin. Immunol. 2006, 117, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Ramphul, K.; Lv, J.; Hua, L.; Liu, Q.; Fang, D.; Ji, R.; Bao, Y. Single nucleotide polymorphisms predisposing to asthma in children of Mauritian Indian and Chinese Han ethnicity. Braz. J. Med. Biol. Res. 2014, 47, 394–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Li, J.; Ding, Z.; Li, J.; Li, B.; Yu, Z.; Tan, W. Association between IL-13 +1923C/T polymorphism and asthma risk: A meta-analysis based on 26 case-control studies. Biosci. Rep. 2017, 37, BSR20160505. [Google Scholar] [CrossRef] [Green Version]
- Utsumi, Y.; Miyamoto, T.; Sekimura, K.; Sasaki, N.; Suzuki, N.; Nakamura, Y.; Kobayashi, H.; Yamaucho, K. The effect of single-nucleotide polymorphism in IL-13 on airway hyperresponsiveness in asthmatics. Eur. Respir. J. 2012, 40, P1786. [Google Scholar]
- Vladich, F.D.; Brazille, S.M.; Stern, D.; Peck, M.L.; Ghittoni, R.; Vercelli, D. IL-13 R130Q, a common variant associated with allergy and asthma, enhances effector mechanisms essential for human allergic inflammation. J. Clin. Investig. 2005, 115, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Imraish, A.; Abu-Thiab, T.; Zihlif, M. IL-13 and FOXO3 genes polymorphisms regulate IgE levels in asthmatic patients. Biomed. Rep. 2021, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.-Z.; Deng, J.-M.; Xu, H.; Nong, Z.-X.; Xiao, C.-Q.; Liu, Z.-M.; Qin, S.-M.; Jiang, H.-X.; Liu, G.-N.; Chen, Y.-Q. Effect of Inhaled Interleukin-4 on Airway Hyperreactivity in Asthmatics. Am. J. Respir. Crit. Care Med. 1998, 157, 1818–1821. [Google Scholar] [CrossRef] [PubMed]
- Steinke, J.W.; Borish, L. Th2 cytokines and asthma—Interleukin-4: Its role in the pathogenesis of asthma, and targeting it for asthma treatment with interleukin-4 receptor antagonists. Respir. Res. 2001, 2, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Basehore, M.J.; Howard, T.D.; Lange, L.A.; Moore, W.C.; Hawkins, G.A.; Marshik, P.L.; Harkins, M.S.; Meyers, D.A.; Bleecker, E.R. A comprehensive evaluation of IL4 variants in ethnically diverse populations: Association of total serum IgE levels and asthma in white subjects. J. Allergy Clin. Immunol. 2004, 114, 80–87. [Google Scholar] [CrossRef]
- Suzuki, I.; Hizawa, N.; Yamaguchi, E.; Kawakami, Y. Association between a C+33T polymorphism in the IL-4 promoter region and total serum IgE levels. Clin. Exp. Allergy 2000, 30, 1746–1749. [Google Scholar] [CrossRef]
- Rosenwasser, L.J.; Borish, L. Promoter polymorphisms predisposing to the development of asthma and atopy. Clin. Exp. Allergy 1998, 28 (Suppl. 5), 13–15, discussion 26–18. [Google Scholar] [CrossRef]
- Wang, R.-S.; Jin, H.-X.; Shang, S.-Q.; Liu, X.-Y.; Chen, S.-J.; Jin, Z.-B. Associations of IL-2 and IL-4 Expression and Polymorphisms with the Risks of Mycoplasma pneumoniae Infection and Asthma in Children. Arch. Bronconeumol. 2015, 51, 571–578. [Google Scholar] [CrossRef]
- Micheal, S.; Minhas, K.; Ishaque, M.; Ahmed, F.; Ahmed, A. IL-4 gene polymorphisms and their association with atopic asthma and allergic rhinitis in Pakistani patients. J. Investig. Allergy Clin. Immunol. 2013, 23, 107–111. [Google Scholar]
- Terasaki, P.I. History of HLA: Ten Recollections; UCLA Tissue Typing Laboratory Press: Los Angeles, CA, USA, 1990. [Google Scholar]
- Svejgaard, A.; Platz, P.; Ryder, L.P.; Nielsen, L.S.; Thomsen, M. HL-A and Disease Associations—A Survey. Transplant Rev. 1975, 22, 3–43. [Google Scholar] [CrossRef]
- Lasky-Su, J.; Himes, B.E.; Raby, B.A.; Klanderman, B.J.; Sylvia, J.S.; Lange, C.; Melen, E.; Martinez, F.D.; Israel, E.; Gauderman, J.; et al. HLA-DQ strikes again: Genome-wide association study further confirms HLA-DQ in the diagnosis of asthma among adults. Clin. Exp. Allergy 2012, 42, 1724–1733. [Google Scholar] [CrossRef]
- Gao, J.; Lin, Y.; Qiu, C.; Liu, Y.; Ma, Y.; Gao, J.; Liu, Y. Relationship between HLA-DQA1, -DQB1 genes polymorphism and susceptilibity to bronchial asthma among Northern Hans. Zhonghua Yi Xue Za Zhi 2002, 82, 379–383. [Google Scholar] [PubMed]
- Guo, X.; Ni, P.; Li, L. Association between asthma and the polymorphism of HLA-DQ genes. Zhonghua Jie He He Hu Xi Za Zhi = Zhonghua Jiehe He Huxi Zazhi = Chin. J. Tuberc. Respir. Dis. 2001, 24, 139–141. [Google Scholar]
- Movahedi, M.; Moin, M.; Gharagozlou, M.; Aghamohammadi, A.; Dianat, S.; Moradi, B.; Nicknam, M.H.; Nikbin, B.; Amirzargar, A. Association of HLA class II alleles with childhood asthma and Total IgE levels. Iran. J. Allergy Asthma Immunol. 2008, 7, 215–220. [Google Scholar]
- Munthe-Kaas, M.C.; Carlsen, K.L.; Carlsen, K.H.; Egeland, T.; Håland, G.; Devulapalli, C.S.; Akselsen, H.; Undlien, D. HLA Dr-Dq haplotypes and the TNFA-308 polymorphism: Associations with asthma and allergy. Allergy 2007, 62, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Awasthi, S.; Gao, P. ADAM Metallopeptidase Domain 33 (ADAM33): A Promising Target for Asthma. Mediat. Inflamm. 2014, 2014, 572025. [Google Scholar] [CrossRef] [Green Version]
- Powell, R.M.; Hamilton, L.M.; Holgate, S.T.; Davies, D.E.; Holloway, J.W. ADAM33: A novel therapeutic target for asthma. Expert Opin. Ther. Targets 2003, 7, 485–494. [Google Scholar] [CrossRef]
- Van Eerdewegh, P.; Little, R.D.; Dupuis, J.; Del Mastro, R.G.; Falls, K.; Simon, J.; Torrey, D.; Pandit, S.; McKenny, J.; Braunschweiger, K.; et al. Association of the ADAM33 gene with asthma and bronchial hyperresponsiveness. Nature 2002, 418, 426–430. [Google Scholar] [CrossRef]
- Farjadian, S.; Moghtaderi, M.; Hoseini-Pouya, B.-A.; Ebrahimpour, A.; Nasiri, M. ADAM33 gene polymorphisms in Southwestern Iranian patients with asthma. Iran J. Basic Med. Sci. 2018, 21, 813–817. [Google Scholar] [CrossRef]
- Schedel, M.; Depner, M.; Schoen, C.; Weiland, S.K.; Vogelberg, C.; Niggemann, B.; Lau, S.; Illig, T.; Klopp, N.; Wahn, U.; et al. The role of polymorphisms in ADAM33, a disintegrin and metalloprotease 33, in childhood asthma and lung function in two German populations. Respir. Res. 2006, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Lin, R.; Wang, C.-C.; Rei, J.; Sun, Y.; Yang, Y.-L.; Lin, Y.-Y. ADAM33 gene polymorphisms identified to be associated with asthma in a Chinese Li population. Biomed. Rep. 2017, 6, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Thongngarm, T.; Jameekornrak, A.; Limwongse, C.; Sangasapaviliya, A.; Jirapongsananuruk, O.; Assawamakin, A.; Chaiyaratana, N.; Luangwedchakarn, V.; Thongnoppakhun, W. Association between ADAM33 polymorphisms and asthma in a Thai population. Asian Pac. J. Allergy Immunol. 2008, 26, 205–211. [Google Scholar] [PubMed]
- Awasthi, S.; Tripathi, P.; Ganesh, S.; Husain, N. Association of ADAM33 gene polymorphisms with asthma in Indian children. J. Hum. Genet. 2011, 56, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, Y.; Karunas, A.; Gimalova, G.; Guryeva, L.; Ramazanova, N.; Levashova, S.; Muhtarova, L.; Murzina, R.; Etkina, E.; Zagidullin, S.; et al. Association of ADAM33 gene polymorphisms with asthma in Volga-Ural region of Russia. Eur. Respir. J. 2011, 38, p436. [Google Scholar]
- Ning, X.; Zhang, Y.; Wu, H.; Bai, L.; Gong, C.; Wang, Z. Genetic association of ADAM33 polymorphisms with childhood asthma in Chinese Han population: A case-control study. Medicine 2019, 98, e17327. [Google Scholar] [CrossRef] [PubMed]
- Hansdottir, S.; Monick, M.M.; Hinde, S.L.; Lovan, N.; Look, D.C.; Hunninghake, G.W. Respiratory Epithelial Cells Convert Inactive Vitamin D to Its Active Form: Potential Effects on Host Defense. J. Immunol. 2008, 181, 7090–7099. [Google Scholar] [CrossRef]
- Martineau, A.R.; Cates, C.J.; Urashima, M.; Jensen, M.; Griffiths, A.P.; Nurmatov, U.; Sheikh, A.; Griffiths, C.J. Vitamin D for the management of asthma. Cochrane Database Syst. Rev. 2016, 2019, CD011511. [Google Scholar] [CrossRef]
- Mirzakhani, H.; Al-Garawi, A.A.; Carey, V.J.; Qiu, W.; Litonjua, A.A.; Weiss, S.T. Expression network analysis reveals cord blood vitamin D-associated genes affecting risk of early life wheeze. Thorax 2019, 74, 200–202. [Google Scholar] [CrossRef]
- Kilic, M.; Ecin, S.; Taskin, E.; Sen, A.; Kara, M. The Vitamin D Receptor Gene Polymorphisms in Asthmatic Children: A Case-Control Study. Pediatr. Allergy Immunol. Pulmonol. 2019, 32, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.E.; Hassan, M.H.; Toghan, R.; Rashwan, N.I. Analysis of 25-hydroxy cholecalciferol, immunoglobulin E, and vitamin D receptor single nucleotide polymorphisms (Apa 1, Taq 1, and Bsm 1), among sample of Egyptian children with bronchial asthma: A case-control study. Pediatr. Pulmonol. 2020, 55, 1349–1358. [Google Scholar] [CrossRef]
- Hutchinson, K.; Kerley, C.P.; Faul, J.; Greally, P.; Coghlan, D.; Louw, M.; Elnazir, B.; Rochev, Y. Vitamin D receptor variants and uncontrolled asthma. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 108–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maalmi, H.; Sassi, F.H.; Berraies, A.; Ammar, J.; Hamzaoui, K.; Hamzaoui, A. Association of vitamin D receptor gene polymorphisms with susceptibility to asthma in Tunisian children: A case control study. Hum. Immunol. 2013, 74, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Pillai, D.K.; Iqbal, S.F.; Benton, A.S.; Lerner, J.; Wiles, A.; Foerster, M.; Ozedirne, T.; Holbrook, H.P.; Payne, P.W., Jr.; Gordish-Dressman, H.; et al. Associations between genetic variants in vitamin D metabolism and asthma characteristics in young African Americans: A pilot study. J. Investig. Med. 2011, 59, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Kabesch, M.; Schedel, M.; Carr, D.; Woitsch, B.; Fritzsch, C.; Weiland, S.K.; von Mutius, E. IL-4/IL-13 pathway genetics strongly influence serum IgE levels and childhood asthma. J. Allergy Clin. Immunol. 2006, 117, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Berenguer, A.G.; Fernandes, A.T.; Oliveira, S.; Rodrigues, M.; Ornelas, P.; Romeira, D.; Serrão, T.; Rosa, A.; Câmara, R. Genetic polymorphisms and asthma: Findings from a case–control study in the Madeira island population. Biol. Res. 2014, 47, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulou, A.; Kouis, P.; Middleton, N.; Kolokotroni, O.; Karpathios, T.; Nicolaidou, P.; Yiallouros, P.K. Association of vitamin D receptor gene polymorphisms and vitamin D levels with asthma and atopy in Cypriot adolescents: A case–control study. Multidiscip. Respir. Med. 2015, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Ma, T. Associations of Genetic Polymorphisms Relevant to Metabolic Pathway of Vitamin D3 with Development and Prognosis of Childhood Bronchial Asthma. DNA Cell Biol. 2017, 36, 682–692. [Google Scholar] [CrossRef]
- Bossé, Y. Asthmatic airway hyperresponsiveness: The ants in the tree. Trends Mol. Med. 2012, 18, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Ducharme, F.M.; Lasserson, T.J.; Cates, C.J. Long-acting beta2-agonists versus anti-leukotrienes as add-on therapy to inhaled corticosteroids for chronic asthma. Cochrane Database Syst. Rev. 2006, 4, Cd003137. [Google Scholar] [CrossRef]
- Drazen, J.M.; Silverman, E.K.; Lee, T.H. Heterogeneity of therapeutic responses in asthma. Br. Med. Bull. 2000, 56, 1054–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantisira, K.G.; Lake, S.; Silverman, E.S.; Palmer, L.; Lazarus, R.; Silverman, E.K.; Liggett, S.B.; Gelfand, E.W.; Rosenwasser, L.J.; Richter, B.; et al. Corticosteroid pharmacogenetics: Association of sequence variants in CRHR1 with improved lung function in asthmatics treated with inhaled corticosteroids. Hum. Mol. Genet. 2004, 13, 1353–1359. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Hu, X.; Zheng, X.; Kuang, J.; Liu, C.; Wang, X.; Tang, Y. Effects of STIP1 and GLCCI1 polymorphisms on the risk of childhood asthma and inhaled corticosteroid response in Chinese asthmatic children. BMC Pulm. Med. 2020, 20, 303. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, G.A.; Lazarus, R.; Smith, R.S.; Tantisira, K.G.; Meyers, D.A.; Peters, S.P.; Weiss, S.T.; Bleecker, E.R. The glucocorticoid receptor heterocomplex gene STIP1 is associated with improved lung function in asthmatic subjects treated with inhaled corticosteroids. J. Allergy Clin. Immunol. 2009, 123, 1376–1383.e7. [Google Scholar] [CrossRef] [Green Version]
- Keskin, O.; Uluca, Ü.; Birben, E.; Coşkun, Y.; Ozkars, M.Y.; Keskin, M.; Kucukosmanoglu, E.; Kalayci, O. Genetic associations of the response to inhaled corticosteroids in children during an asthma exacerbation. Pediatr. Allergy Immunol. 2016, 27, 507–513. [Google Scholar] [CrossRef] [PubMed]
- Stockmann, C.; Fassl, B.; Gaedigk, R.; Nkoy, F.; Uchida, D.A.; Monson, S.; Reilly, C.A.; Leeder, J.S.; Yost, G.S.; Ward, R.M. Fluticasone Propionate Pharmacogenetics: CYP3A4*22 Polymorphism and Pediatric Asthma Control. J. Pediatr. 2013, 162, 1222–1227.e2. [Google Scholar] [CrossRef] [Green Version]
- Tantisira, K.G.; Hwang, E.S.; Raby, B.A.; Silverman, E.S.; Lake, S.L.; Richter, B.G.; Peng, S.L.; Drazen, J.M.; Glimcher, L.H.; Weiss, S.T. TBX21: A functional variant predicts improvement in asthma with the use of inhaled corticosteroids. Proc. Natl. Acad. Sci. USA 2004, 101, 18099–18104. [Google Scholar] [CrossRef]
- Ye, Y.M.; Lee, H.Y.; Kim, S.-H.; Jee, Y.-K.; Lee, S.K.; Park, H.S. Pharmacogenetic study of the effects of NK2R G231E G>A and TBX21 H33Q C>G polymorphisms on asthma control with inhaled corticosteroid treatment. J. Clin. Pharm. Ther. 2009, 34, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Duong-Quy, S.; Le-Thi-Minh, H.; Nguyen-Thi-Bich, H.; Pham-Thu, H.; Thom, V.; Pham-Thi-Hong, N.; Duong-Thi-Ly, H.; Nguyen-Huy, B.; Ngo-Minh, X.; Nguyen-Thi-Dieu, T.; et al. Correlations between exhaled nitric oxide, rs28364072 polymorphism of FCER2 gene, asthma control, and inhaled corticosteroid responsiveness in children with asthma. J. Breath Res. 2020, 15, 016012. [Google Scholar] [CrossRef]
- Wan, Z.; Tang, Y.; Song, Q.; Zhang, J.; Xie, W.; He, Y.; Huang, R.; Zheng, X.; Liu, C.; Liu, J. Gene polymorphisms in VEGFA and COL2A1 are associated with response to inhaled corticosteroids in children with asthma. Pharmacogenomics 2019, 20, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Tantisira, K.G.; Lasky-Su, J.; Harada, M.; Murphy, A.; Litonjua, A.A.; Himes, B.E.; Lange, C.; Lazarus, R.; Sylvia, J.; Klanderman, B.; et al. Genomewide Association between GLCCI1 and Response to Glucocorticoid Therapy in Asthma. N. Engl. J. Med. 2011, 365, 1173–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantisira, K.G.; Damask, A.; Szefler, S.J.; Schuemann, B.; Markezich, A.; Su, J.; Klanderman, B.; Sylvia, J.; Wu, R.; Martinez, F.; et al. Genome-wide Association Identifies the T Gene as a Novel Asthma Pharmacogenetic Locus. Am. J. Respir. Crit. Care Med. 2012, 185, 1286–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bollag, R.J.; Siegfried, Z.; Cebra-Thomas, J.A.; Garvey, N.; Davison, E.M.; Silver, L.M. An ancient family of embryonically expressed mouse genes sharing a conserved protein motif with the T locus. Nat. Genet. 1994, 7, 383–389. [Google Scholar] [CrossRef]
- Liu, L.; Yue, D.; Hu, L.; Wang, F.; Huang, Y.; Liao, Y. Relationship between interleukin-13 rs20541 single nucleotide polymorphisms and therapeutic efficacy in children with asthma. J. Int. Med. Res. 2020, 48, 300060520929179. [Google Scholar] [CrossRef] [PubMed]
- Bivona, G.; Agnello, L.; Ciaccio, M. The immunological implication of the new vitamin D metabolism. Cent. Eur. J. Immunol. 2018, 43, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Sjoukes, A.; Richards, D.; Banya, W.; Hawrylowicz, C.; Bush, A.; Saglani, S. Relationship between serum vitamin D, disease severity, and airway remodeling in children with asthma. Am. J. Respir. Crit. Care Med. 2011, 184, 1342–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, S.; Hübner, M.; Jung, M.; Blettner, M.; Buhl, R. Severe and uncontrolled adult asthma is associated with vitamin D insufficiency and deficiency. Respir. Res. 2013, 14, 25. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, N.A.; Abdel-Rehim, A.S. (Eds.) Influence of vitamin D receptor gene FokI and ApaI polymorphisms on glucocorticoid response in patients with asthma. In International Forum of Allergy & Rhinology; Wiley Online Library: Hoboken, NJ, USA, 2020. [Google Scholar]
- Tantisira, K.G.; Small, K.M.; Litonjua, A.A.; Weiss, S.T.; Liggett, S.B. Molecular properties and pharmacogenetics of a polymorphism of adenylyl cyclase type 9 in asthma: Interaction between β-agonist and corticosteroid pathways. Hum. Mol. Genet. 2005, 14, 1671–1677. [Google Scholar] [CrossRef] [Green Version]
- Kotani, Y.; Nishimura, Y.; Maeda, H.; Yokoyama, M. β2-Adrenergic Receptor Polymorphisms Affect Airway Responsiveness to Salbutamol in Asthmatics. J. Asthma 1999, 36, 583–590. [Google Scholar] [CrossRef]
- Martinez, F.D.; Graves, P.E.; Baldini, M.; Solomon, S.; Erickson, R. Association between genetic polymorphisms of the beta2-adrenoceptor and response to albuterol in children with and without a history of wheezing. J. Clin. Investig. 1997, 100, 3184–3188. [Google Scholar] [CrossRef]
- Lima, J.J.; Thomason, D.B.; Mohamed, M.H.N.; Eberle, L.V.; Self, T.H.; Johnson, J.A. Impact of genetic polymorphisms of the β2-adrenergic receptor on albuterol bronchodilator pharmacodynamics. Clin. Pharmacol. Ther. 1999, 65, 519–525. [Google Scholar] [CrossRef]
- Lima, J.J.; Mohamed, M.H.; Self, T.H.; Eberle, L.V.; Johnson, J.A. Importance of β2Adrenergic Receptor Genotype, Gender and Race on Albuterol-evoked Bronchodilation in Asthmatics. Pulm. Pharmacol. Ther. 2000, 13, 127–134. [Google Scholar] [CrossRef]
- Israel, E.; Drazen, J.M.; Liggett, S.B.; Boushey, H.A.; Cherniack, R.M.; Chinchilli, V.M.; Cooper, D.M.; Fahy, J.V.; Fish, J.E.; Ford, J.G.; et al. The Effect of Polymorphisms of the β2-Adrenergic Receptor on the Response to Regular Use of Albuterol in Asthma. Am. J. Respir. Crit. Care Med. 2000, 162, 75–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israel, E.; Chinchilli, V.M.; Ford, J.G.; Boushey, H.A.; Cherniack, R.; Craig, T.J.; Deykin, A.; Fagan, J.K.; Fahy, J.V.; Fish, J.; et al. Use of regularly scheduled albuterol treatment in asthma: Genotype-stratified, randomised, placebo-controlled cross-over trial. Lancet 2004, 364, 1505–1512. [Google Scholar] [CrossRef]
- Bleecker, E.R.; Postma, D.S.; Lawrance, R.M.; Meyers, D.A.; Ambrose, H.J.; Goldman, M. Effect of ADRB2 polymorphisms on response to longacting β2-agonist therapy: A pharmacogenetic analysis of two randomised studies. Lancet 2007, 370, 2118–2125. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Kunselman, S.J.; Chinchilli, V.M.; Bleecker, E.; Boushey, H.A.; Calhoun, W.J.; Ameredes, B.T.; Castro, M.; Craig, T.J.; Denlinger, L. Effect of β2-adrenergic receptor polymorphism on response to longacting β2 agonist in asthma (LARGE trial): A genotype-stratified, randomised, placebo-controlled, crossover trial. Lancet 2009, 374, 1754–1764. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Ye, Y.M.; Lee, H.Y.; Sin, H.J.; Park, H.S.; Ms, H.Y.L. Combined pharmacogenetic effect of ADCY9 and ADRB2 gene polymorphisms on the bronchodilator response to inhaled combination therapy. J. Clin. Pharm. Ther. 2011, 36, 399–405. [Google Scholar] [CrossRef]
- Poon, A.H.; Tantisira, K.G.; Litonjua, A.A.; Lazarus, R.; Xu, J.; Lasky-Su, J.; Lima, J.J.; Irvin, C.G.; Hanrahan, J.P.; Lange, C. Association of corticotropin releasing hormone receptor 2 (CRHR2) genetic variants with acute bronchodilator response in asthma. Pharm. Genom. 2008, 18, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, K.A.; Torgerson, D.G.; Gignoux, C.R.; Galanter, J.M.; Roth, L.A.; Huntsman, S.; Eng, C.; Oh, S.S.; Yee, S.W.; Lin, L.; et al. A genome-wide association study of bronchodilator response in Latinos implicates rare variants. J. Allergy Clin. Immunol. 2014, 133, 370–378.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litonjua, A.A.; Lasky-Su, J.; Schneiter, K.; Tantisira, K.G.; Lazarus, R.; Klanderman, B.; Lima, J.J.; Irvin, C.G.; Peters, S.P.; Hanrahan, J.P. ARG1 is a novel bronchodilator response gene: Screening and replication in four asthma cohorts. Am. J. Respir. Crit. Care Med. 2008, 178, 688–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonk, J.M.; Postma, D.S.; Maarsingh, H.; Bruinenberg, M.; Koppelman, G.H.; Meurs, H. Arginase 1 and arginase 2 variations associate with asthma, asthma severity and β2 agonist and steroid response. Pharmacogen. Genom. 2010, 20, 179–186. [Google Scholar] [CrossRef]
- Iordanidou, M.; Paraskakis, E.; Tavridou, A.; Paschou, P.; Chatzimichael, A.; Manolopoulos, V.G. G894T polymorphism of eNOS gene is a predictor of response to combination of inhaled corticosteroids with long-lasting β2-agonists in asthmatic children. Pharmacogenomics 2012, 13, 1363–1372. [Google Scholar] [CrossRef]
- Duan, Q.L.; Du, R.; Lasky-Su, J.; Klanderman, B.J.; Partch, A.B.; Peters, S.P.; Irvin, C.G.; Hanrahan, J.P.; Lima, J.J.; Blake, K.V.; et al. A polymorphism in the thyroid hormone receptor gene is associated with bronchodilator response in asthmatics. Pharmacogen. J. 2012, 13, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Himes, B.E.; Jiang, X.; Hu, R.; Wu, A.C.; Lasky-Su, J.A.; Klanderman, B.J.; Ziniti, J.; Senter-Sylvia, J.; Lima, J.J.; Irvin, C.G.; et al. Genome-Wide Association Analysis in Asthma Subjects Identifies SPATS2L as a Novel Bronchodilator Response Gene. PLoS Genet. 2012, 8, e1002824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padhukasahasram, B.K.; Yang, J.J.; Levin, A.M.; Yang, M.; Burchard, E.G.; Kumar, R.; Kwok, P.-Y.; Seibold, M.A.; Lanfear, D.E.; Williams, L.K. Gene-based association identifies SPATA13-AS1 as a pharmacogenomic predictor of inhaled short-acting beta-agonist response in multiple population groups. Pharmacogen. J. 2014, 14, 365–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishe, J.N.; Labilloy, G.; Higley, R.; Casey, D.; Ginn, A.; Baskovich, B.; Blake, K.V. Single Nucleotide Polymorphisms (SNPs) in PRKG1 & SPATA13-AS1 are associated with bronchodilator response: A pilot study during acute asthma exacerbations in African American children. Pharmacogen. Genom. 2021, 31, 146–154. [Google Scholar] [CrossRef]
- Spear, M.L.; Hu, D.; Pino-Yanes, M.; Huntsman, S.; Eng, C.; Levin, A.M.; Ortega, V.E.; White, M.J.; McGarry, M.E.; Thakur, N.; et al. A genome-wide association and admixture mapping study of bronchodilator drug response in African Americans with asthma. Pharmacogen. J. 2019, 19, 249–259. [Google Scholar] [CrossRef]
- Drazen, J.M.; Yandava, C.N.; Dubé, L.; Szczerback, N.; Hippensteel, R.; Pillari, A.; Israel, E.; Schork, N.; Silverman, E.S.; Katz, D.A.; et al. Pharmacogenetic association between ALOX5 promoter genotype and the response to anti-asthma treatment. Nat. Genet. 1999, 22, 168–170. [Google Scholar] [CrossRef]
- Telleria, J.J.; Blanco-Quiros, A.; Varillas, D.; Armentia, A.; Fernandez-Carvajal, I.; Alonso, M.J.; Diez, I. ALOX5 promoter genotype and response to montelukast in moderate persistent asthma. Respir. Med. 2008, 102, 857–861. [Google Scholar] [CrossRef]
- Sampson, A.P.; Siddiqui, S.; Buchanan, D.; Howarth, P.H.; Holgate, S.T.; Holloway, J.W.; Sayers, I. Variant LTC4 synthase allele modifies cysteinyl leukotriene synthesis in eosinophils and predicts clinical response to zafirlukast. Thorax 2000, 55, S28–S31. [Google Scholar] [CrossRef] [Green Version]
- Tantisira, K.G.; Lima, J.; Sylvia, J.; Klanderman, B.; Weiss, S.T. 5-lipoxygenase pharmacogenetics in asthma: Overlap with CystLTR1 loci. Pharmacogen. Genom. 2009, 19, 244. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.J.; Zhang, S.; Grant, A.; Shao, L.; Tantisira, K.G.; Allayee, H.; Wang, J.; Sylvester, J.; Holbrook, J.; Wise, R.; et al. Influence of Leukotriene Pathway Polymorphisms on Response to Montelukast in Asthma. Am. J. Respir. Crit. Care Med. 2006, 173, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Mougey, E.B.; Feng, H.; Castro, M.; Irvin, C.G.; Lima, J.J. Absorption of montelukast is transporter mediated: A common variant of OATP2B1 is associated with reduced plasma concentrations and poor response. Pharmacogen. Genom. 2009, 19, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.-A.; Lee, H.-M.; Joo, H.-J.; Park, I.-B.; Park, J.-Y. Effects of polymorphisms of the SLCO2B1 transporter gene on the pharmacokinetics of montelukast in humans. J. Clin. Pharmacol. 2013, 53, 1186–1193. [Google Scholar] [CrossRef]
- Tapaninen, T.; Karonen, T.; Backman, J.T.; Neuvonen, P.J.; Niemi, M. SLCO2B1 c.935G>A single nucleotide polymorphism has no effect on the pharmacokinetics of montelukast and aliskiren. Pharmacogen. Genom. 2013, 23, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Tantisira, K.G.; Silverman, E.S.; Mariani, T.J.; Xu, J.; Richter, B.G.; Klanderman, B.J.; Litonjua, A.; Lazarus, R.; Rosenwasser, L.J.; Fuhlbrigge, A.L.; et al. FCER2: A pharmacogenetic basis for severe exacerbations in children with asthma. J. Allergy Clin. Immunol. 2007, 120, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Koster, E.S.; Maitland-van der Zee, A.-H.; Tavendale, R.; Mukhopadhyay, S.; Vijverberg, S.J.H.; Raaijmakers, J.A.M.; Palmer, C.N.A. FCER2 T2206C variant associated with chronic symptoms and exacerbations in steroid-treated asthmatic children. Allergy 2011, 66, 1546–1552. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Lin, Y.-C.; Lee, Y.-C.; Wang, J.-Y.; Hsiue, T.-R.; Guo, Y.L. Glutathione S-transferase P1 gene polymorphism and air pollution as interactive risk factors for childhood asthma. Clin. Exp. Allergy 2004, 34, 1707–1713. [Google Scholar] [CrossRef]
- Ercan, H.; Birben, E.; Dizdar, E.A.; Keskin, O.; Karaaslan, C.; Soyer, O.U.; Dut, R.; Sackesen, C.; Besler, T.; Kalayci, O. Oxidative stress and genetic and epidemiologic determinants of oxidant injury in childhood asthma. J. Allergy Clin. Immunol. 2006, 118, 1097–1104. [Google Scholar] [CrossRef]
- Aynacioglu, A.S.; Nacak, M.; Filiz, A.; Ekinci, E.; Roots, I. Protective role of glutathione S-transferase P1 (GSTP1) Val105Val genotype in patients with bronchial asthma. Br. J. Clin. Pharmacol. 2003, 57, 213–217. [Google Scholar] [CrossRef] [Green Version]
- MacIntyre, E.A.; Brauer, M.; Melén, E.; Bauer, C.P.; Bauer, M.; Berdel, D.; Bergström, A.; Brunekreef, B.; Chan-Yeung, M.; Klümper, C.; et al. GSTP1 and TNF Gene Variants and Associations between Air Pollution and Incident Childhood Asthma: The Traffic, Asthma and Genetics (TAG) Study. Environ. Health Perspect. 2014, 122, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Mukhammadiyeva, G.F.; Bakirov, A.B.; Karimov, D.O.; Ziatdinova, M.M.; Valova, Y.V.; Borisova, A.I.; Distanova, A.A. Analysis of the GSTP1 rs 1695 polymorphism association with the development of asthma and phenotypic manifestations. J. Asthma 2021, 59, 1065–1069. [Google Scholar] [CrossRef]
- Becker, S.; Dailey, L.; Soukup, J.M.; Silbajoris, R.; Devlin, R.B. TLR-2 is involved in airway epithelial cell response to air pollution particles. Toxicol. Appl. Pharmacol. 2005, 203, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Kerkhof, M.; Postma, D.S.; Brunekreef, B.; Reijmerink, N.E.; Wijga, A.H.; De Jongste, J.C.; Gehring, U.; Koppelman, G.H. Toll-like receptor 2 and 4 genes influence susceptibility to adverse effects of traffic-related air pollution on childhood asthma. Thorax 2010, 65, 690–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, C.Y.; Pang, L.; Holland, E.; Knox, A.J. TGF-beta1 stimulates IL-8 release, COX-2 expression, and PGE(2) release in human airway smooth muscle cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, L201–L207. [Google Scholar] [CrossRef] [PubMed]
- Kokturk, N.; Tatlicioglu, T.; Memis, L.; Akyurek, N.; Akyol, G. Expression of Transforming Growth Factor β1 in Bronchial Biopsies in Asthma and COPD. J. Asthma 2003, 40, 887–893. [Google Scholar] [CrossRef]
- Salam, M.T.; Gauderman, W.J.; McConnell, R.; Lin, P.-C.; Gilliland, F.D. Transforming Growth Factor-β1 C-509T Polymorphism, Oxidant Stress, and Early-Onset Childhood Asthma. Am. J. Respir. Crit. Care Med. 2007, 176, 1192–1199. [Google Scholar] [CrossRef]
- Akdis, M.B.; Verhagen, J.; Taylor, A.; Karamloo, F.; Karagiannidis, C.; Crameri, R.; Thunberg, S.; Deniz, G.N.; Valenta, R.; Fiebig, H.; et al. Immune Responses in Healthy and Allergic Individuals Are Characterized by a Fine Balance between Allergen-specific T Regulatory 1 and T Helper 2 Cells. J. Exp. Med. 2004, 199, 1567–1575. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues Marques, C.; Sampaio Dotto Fiuza, B.; Magalhães da Silva, T.; Cana Brasil Carneiro, T.; Santos Costa, R.; Francisca de Assis Silva, M.; Lauane Luz Viana, W.; Leão Carneiro, V.; Maria Alcantara-Neves, N.; Lima Barreto, M.; et al. Impact of FOXP3 gene polymorphisms and gene-environment interactions in asthma and atopy in a Brazilian population. Gene 2022, 838, 146706. [Google Scholar] [CrossRef]
- Shimbara, A.; Christodoulopoulos, P.; Soussi-Gounni, A.; Olivenstein, R.; Nakamura, Y.; Levitt, R.C.; Nicolaides, N.C.; Holroyd, K.J.; Tsicopoulos, A.; Lafitte, J.-J.; et al. IL-9 and its receptor in allergic and nonallergic lung disease: Increased expression in asthma. J. Allergy Clin. Immunol. 2000, 105, 108–115. [Google Scholar] [CrossRef]
- Sordillo, J.E.; Kelly, R.; Bunyavanich, S.; McGeachie, M.; Qiu, W.; Croteau-Chonka, D.C.; Soto-Quiros, M.; Avila, L.; Celedón, J.C.; Brehm, J.M.; et al. Genome-wide expression profiles identify potential targets for gene-environment interactions in asthma severity. J. Allergy Clin. Immunol. 2015, 136, 885–892.e2. [Google Scholar] [CrossRef] [Green Version]
- Schedel, M.; Michel, S.; Gaertner, V.D.; Toncheva, A.A.; Depner, M.; Binia, A.; Schieck, M.; Rieger, M.T.; Klopp, N.; Von Berg, A.; et al. Polymorphisms related to ORMDL3 are associated with asthma susceptibility, alterations in transcriptional regulation of ORMDL3, and changes in TH2 cytokine levels. J. Allergy Clin. Immunol. 2015, 136, 893–903.e14. [Google Scholar] [CrossRef]
- Bräuner, E.V.; Loft, S.; Raaschou-Nielsen, O.; Vogel, U.; Andersen, P.S.; Sørensen, M. Effects of a 17q21 chromosome gene variant, tobacco smoke and furred pets on infant wheeze. Genes Immun. 2012, 13, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paruchuri, S.; Tashimo, H.; Feng, C.; Maekawa, A.; Xing, W.; Jiang, Y.; Kanaoka, Y.; Conley, P.; Boyce, J.A. Leukotriene E4–induced pulmonary inflammation is mediated by the P2Y12 receptor. J. Exp. Med. 2009, 206, 2543–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunyavanich, S.; Boyce, J.A.; Raby, B.A.; Weiss, S.T. Gene-by-environment effect of house dust mite on purinergic receptor P2Y12 (P2RY12) and lung function in children with asthma. Clin. Exp. Allergy 2012, 42, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brigelius-Flohé, R.; Kipp, A.P. Chapter Four—Selenium in the Redox Regulation of the Nrf2 and the Wnt Pathway. In Methods in Enzymology; Cadenas, E., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 527, pp. 65–86. [Google Scholar]
- Linaker, C.H.; Coggon, D.; Holgate, S.T.; Clough, J.; Josephs, L.; Chauhan, A.J.; Inskip, H. Personal exposure to nitrogen dioxide and risk of airflow obstruction in asthmatic children with upper respiratory infection. Thorax 2000, 55, 930–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ungvári, I.; Hadadi, É.; Virág, V.; Nagy, A.; Kiss, A.; Kalmár, A.; Zsigmond, G.; Semsei, F.; Falus, A.; Szalai, C. Relationship between air pollution, NFE2L2 gene polymorphisms and childhood asthma in a Hungarian population. J. Community Genet. 2012, 3, 25–33. [Google Scholar] [CrossRef]
- Du, J.-W.; Xu, Z.-L.; Xu, Q.-X. Interaction of Interleukin 7 Receptor (IL7R) and IL6 Gene Polymorphisms with Smoking Associated with Susceptibility to Asthma in Chinese Han Adults. Immunol. Investig. 2022, 51, 1364–1371. [Google Scholar] [CrossRef]
- Shao, Y.; Zhang, Y.; Liu, M.; Fernandez-Beros, M.-E.; Qian, M.; Reibman, J. Gene-Environment Interaction between the IL1RN Variants and Childhood Environmental Tobacco Smoke Exposure in Asthma Risk. Int. J. Environ. Res. Public Health 2020, 17, 2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramadas, R.A.; Sadeghnejad, A.; Karmaus, W.; Arshad, S.H.; Matthews, S.; Huebner, M.; Kim, D.-Y.; Ewart, S.L. Interleukin-1R antagonist gene and pre-natal smoke exposure are associated with childhood asthma. Eur. Respir. J. 2007, 29, 502–508. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Study Design | Race | SNP | Allele | Association | Ref |
|---|---|---|---|---|---|---|
| TSLP | Candidate gene study | Caucasian | rs1837253 | C/T | Inversely correlated with risk of asthma | [16] |
| Candidate gene study | Middle eastern | rs2289276 | C/T | Inversely correlated with the risk of asthma | [17] | |
| rs2289278 | C/G | No association | ||||
| Candidate gene study | Asian | rs3806933 | C/T | Associated with asthma susceptibility in adult and childhood asthma | [13] | |
| rs2289276 | C/T | |||||
| rs2289278 | C/G | Associated with lung function [FEV1/FVC] | ||||
| Candidate gene study | Caucasian | rs3806933 | C/T | Associated with asthma in boys | [14] | |
| rs2289276 | C/T | Associated with higher eosinophil counts in asthmatic girls Associated with lower FEV1 level in asthmatic | ||||
| rs10073816 | G/A | No association | ||||
| rs11466749 | A/G | The presence of allergic rhinitis in asthmatic children strengthened the association of the rs11466749 genotype with asthma | ||||
| Candidate gene study | Mixed | rs1837253 | C/T | Associated with a reduced risk of asthma in males | [15] | |
| rs2289276 | C/T | Associated with a reduced risk of asthma in females | ||||
| IL-33 | Candidate gene study | Asian | rs4742170 | C/T | Associated with risk of higher FeNO at baseline | [22] |
| rs2381416 | A/C | No association | ||||
| rs928413 | A/G | No association | ||||
| rs992969 | A/G | No association | ||||
| GWAS | Caucasian | rs146597587 | G/C | Associated with lower eosinophil count and reduced risk of asthma | [25] | |
| Candidate gene study | Caucasian | rs1412420 | C/A | Associated with adult-onset asthma | [23] | |
| rs7037276 | G/C | |||||
| rs996029 | A/T | |||||
| Candidate gene study | Middle eastern | rs1342326 | A/C | Associated with higher risk of asthma | [24] | |
| rs3939286 | G/A | |||||
| IL1RL1 | Candidate gene study | Caucasian | rs11685480 | G/A | Associated with lower blood eosinophil count | [30] |
| rs1420102 | C/A | |||||
| rs1041973 | C/A | Associated with decreased risk of developing asthma | ||||
| Candidate gene study | Puerto Rican | rs1921622 | A/G | No association | [29] | |
| Candidate gene study | Asian | rs1420102 | C/A | Associated with higher risk of asthma | [28] | |
| rs13431828 | C/T | No association | ||||
| Candidate gene study | Mixed | rs13431828 | T/C | Associated with increased risk of exacerbation | [32] | |
| rs1041973 | A/C | No association | ||||
| rs1946131 | G/A | |||||
| rs1420101 | G/A | Associated with lower serum sST2 levels | ||||
| rs1921622 | G/A | |||||
| rs10204137 | G/A | |||||
| IL-13 | Candidate gene study | Mixed | rs1295686 | C/T | Associated with increased risk of asthma | [37] |
| Candidate gene study | Middle eastern | rs20541 | G/A | Associated with increased risk of asthma | [41] | |
| Candidate gene study | Middle eastern | rs1295686 | C/T | Associated with increased risk of asthma | [35] | |
| rs20541 | G/A | |||||
| rs1800925 | G/A | No association | ||||
| rs762534 | C/A | |||||
| Candidate gene study | Asian | rs20541 | G/A | Associated with AHR | [39] | |
| IL-4 | Candidate gene study | Caucasian | rs2243250 | C/T | Associated with higher serum IgE levels | [74] |
| Candidate gene study | Asian | rs2227284 | T/G | Associated with reduced risk of asthma | [47] | |
| rs2243250 | T/C | No association | ||||
| rs2070874 | T/C | |||||
| rs2243290 | A/C | |||||
| Middle eastern | rs2243250 | T/C | Associated with asthma and allergic rhinitis | [48] | ||
| rs2227284 | T/G | |||||
| rs2070874 | T/C | No association | ||||
| HLA | GWAS | Mixed | rs9272346 | A/G | Associated with increased risk asthma | [51] |
| ADAM33 | Candidate gene study | Middle eastern | rs2280091 | A/G | No association | [59] |
| rs3918396 | G/A | |||||
| rs2280089 | G/A | |||||
| rs511898 | C/A | |||||
| Candidate gene study | Asian | rs44707 | G/C | Associated with severe asthma | [61] | |
| rs2280089 | G/A | Associated with increased risk asthma | ||||
| rs2787094 | G/C | Associated with less severe asthma | ||||
| rs612709 | G/A | No association | ||||
| rs511898 | C/A | |||||
| rs2280091 | A/G | |||||
| rs528557 | G/C | |||||
| rs3918396 | G/A | |||||
| Candidate gene study | South Asian | rs528557 | G/C | Associated with increased risk asthma | [63] | |
| rs597980 | C/T | |||||
| rs511898 | C/A | |||||
| rs44707 | G/C | |||||
| rs2787094 | G/C | |||||
| Candidate gene study | Asian | rs528557 | G/C | Associated with low severity asthma | [62] | |
| rs598418 | C/T | Associated with high severity asthma | ||||
| rs44707 | G/C | No association | ||||
| rs2853209 | A/T | |||||
| rs597980 | C/T | |||||
| rs11905233 | G/A | |||||
| rs2787094 rs3746631 | G/C | |||||
| Candidate gene study | Caucasian | rs2787094 | G/C | Associated with mild asthma | [75] | |
| Candidate gene study | Caucasian | rs511898 | G/A | No association | [60] | |
| rs3918395 | G/T | |||||
| rs3918396 | G/A | |||||
| rs528557 | G/C | |||||
| rs44707 | G/C | |||||
| rs597980 | C/T | |||||
| rs574174 | G/A | |||||
| rs2280091 | A/G | |||||
| rs2280090 | C/T | |||||
| rs2787094 | G/C | |||||
| Candidate gene study | Caucasian | rs44707 | G/C | Associated with increased risk of asthma | [64] | |
| rs2787095 | G/T | No association | ||||
| rs2485700 | G/A | |||||
| rs2280091 | A/G | |||||
| Candidate gene study | Asian | rs678881 | G/C | Associated with increased risk of asthma | [65] | |
| rs2280089 | G/A | No association | ||||
| rs2853209 | A/T | |||||
| VDR | Candidate gene study | Caucasian | rs731236 | C/T | Associated with increased risk of asthma | [69] |
| rs7975232 | A/C | Associated with reduced mRNA gene expression in asthmatic group. | ||||
| Candidate gene study | North African | rs731236 | C/T | Associated with increased risk of asthma | [72] | |
| rs7975232 | A/C | |||||
| rs2228570 | T/C | |||||
| rs1544410 | G/A | |||||
| Candidate gene study | North African | rs7975232 | A/C | No association | [70] | |
| rs731236 | C/T | Associated with increased risk of asthma | ||||
| rs1544410 | G/A | |||||
| Candidate gene study | Caucasian | rs731236 | C/T | Associated with increased risk of asthma | [71] | |
| rs7975232 | A/C | |||||
| Candidate gene study | Caucasian | rs731236 | C/T | Associated with increased risk of asthma | [76] | |
| rs7975232 | A/C | No association | ||||
| rs1544410 | G/A | |||||
| Candidate gene study | Asian | rs1544410 | G/A | Associated with increased risk of asthma | [77] | |
| rs7975232 | A/C | |||||
| rs2228570 | T/C |
| Gene | Study Design | Race | SNP | Allele | Drug Class | Response Phenotype | Ref |
|---|---|---|---|---|---|---|---|
| CRHR1 | Candidate gene study | Mixed | rs242941 | C/A | ICS | Variants improves FEV1 | [81] |
| rs1876828 | C/T | ||||||
| STIP1 | Candidate gene study | Asian/Caucasian | rs2236647 | C/T | ICS | Variants improves FEV1 | [82,83] |
| rs6591838 | A/G | ||||||
| rs1011219 | G/A | ||||||
| rs4980524 | A/C | ||||||
| NR3C1 | Candidate gene study | Caucasian | rs41423247 | G/C | ICS | Variants improve FEV1 | [84] |
| TBX21 | Candidate gene study | Mixed | rs2240017 | C/G | ICS | Variant improved airway hyperresponsiveness | [86,87] |
| GLCCI1 | GWAS | Mixed | rs37973 | G/T | ICS | rs37973 Wild-type allele had improved FEV1 and response to ICS compared to mutant allele | [90] |
| rs37972 | A/T | ||||||
| T gene | GWAS | Mixed | rs3127412 | A/C | ICS | Wild-type allele had improved FEV1 and response to ICS compared to mutant allele | [91] |
| rs6456042 | C/A | ||||||
| rs3099266 | A/T | ||||||
| ADCY9 | Candidate gene study | Mixed | rs2230739 | T/C | ICS B2 agonist | Variants improved bronchodilator response | [98] |
| CYP3A4 | Candidate gene study | Mixed | CYP3A4*22 allele | ICS | T-allele variants improved asthma control | [85] | |
| FCER2 | Candidate gene study | Mixed | rs28364072 | A/G | ICS | Variants improve FEV1 | [88,126,127] |
| VEGFA | Candidate gene study | Mixed | rs3025039 | C/T | ICS | Variants improve FEV1 | [89] |
| COL2A1 | Candidate gene study | Mixed | rs3809324 | G/T | ICS | Variants improve FEV1 | [89] |
| IL-13 | Candidate gene study | Mixed | rs20541 | C/A | ICS | Variants improved FEV1 | [93] |
| Vitamin D receptor | Candidate gene study | Middle eastern | FokI rs228570 Apal Rs7975232 | T/C A/C | ICS | FokI variants are associated with GC-resistance | [97] |
| ADRB2 | Candidate Gene Study | Mixed | rs1042716 | G/T | SABA | Short-term use: Arg 16 has greater bronchodilation response than Gly 16 Long-term use: no allele effect | [99,100,101,102] |
| rs11959427 | A/C | ||||||
| ADCY9 | Candidate gene study | Mixed | rs2230739 | T/C | SABA | Acute FEV1 bronchodilation | [98] |
| CRHR2 | Candidate Gene Study | Mixed /South American | rs7793837 | A/T | SABA | Acute FEV1 bronchodilation | [108,109] |
| ARGI | Candidate Gene Study | Caucasian | rs2781659 | A/G | SABA | Acute FEV1 bronchodilation | [110,111] |
| rs2781667 | C/T | ||||||
| ARG2 | Candidate Gene Study | Caucasian | rs7140310 | T/G | SABA | Acute FEV1 bronchodilation | [111] |
| rs10483801 | C/A | ||||||
| THRB | Candidate Gene Study | Caucasian | rs892940 | G/A | SABA | Acute FEV1 bronchodilation | [113] |
| SPATS2L | GWAS | Caucasian | rs295137 | C/T | SABA | Acute FEV1 bronchodilation | [114] |
| SPATA13-AS1 | GWAS | Mixed/African American | rs912142 | A/G | SABA | Acute FEV1 bronchodilation | [115,116] |
| SLC22A15 | Admixture mapping | South American | rs1281748 | A/G | SABA | Acute FEV1 bronchodilation | [109] |
| rs1281743 | G/A | ||||||
| PRKG1 | Candidate Gene Study | Mixed | rs7081864 rs7903366 | Admixture mapping | SABA | Acute FEV1 bronchodilation | [116,117] |
| ADRB2 | Candidate Gene Study | Mixed | rs1042716 | G/T | LABA | No allele effect | [105,106] |
| rs1800888 | C/T | ||||||
| ADCY9 | Candidate Gene Study | Asian | rs2230739 | T/C | LABA | Long-term increased FEV1 response | [107] |
| NOS3 | Candidate Gene Study | Mixed | rs1799983 | A/T | LABA | Acute FEV1 bronchodilation in Asp 298 Glu | [112] |
| ALOX5 | Candidate Gene Study | Mixed | rs892690 | C/T | Antileukotriene | Wild type allele increased BDR | [118,119] |
| rs2115819 | G/A | ||||||
| rs10507391 | C/A | ||||||
| rs4986832 | G/A | ||||||
| rs4987105 | C/T | ||||||
| LTC4 | Candidate Gene Study | Mixed | rs272431 | G/T | Antileukotriene | Mutant allele increased BDR | [120,121] |
| rs730012 | A/C | ||||||
| MRP1 | Candidate Gene Study | Mixed | rs119774 | C/T | Antileukotriene | Mutant allele increased BDR | [121] |
| rs215066 | G/A | ||||||
| LTA4 | Candidate Gene Study | Mixed | rs2660845 | A/G | Antileukotriene | Wild type allele decreased asthma exacerbations | [122] |
| SLCo2B1 | Candidate Gene Study | Mixed | rs12422149 | G/A | Antileukotriene | Increased symptom control | [123] |
| Gene | Study Design | Race | SNP | Allele | Association | Ref |
|---|---|---|---|---|---|---|
| FOXP3 | Candidate gene | Brazilian | rs2232368 | C/T | Associated with asthma susceptibility and onset of atopy in females. | [139] |
| TGF-B1 | Candidate gene | Mixed | rs4803457 | C/T | -509TT genotype carriers have higher susceptibility to early-onset childhood asthma | [137] |
| GSTP1 | Candidate gene | Canadian, Swedish, German, Netherlands (six birth cohorts) | rs1138272 | C/T | Minor allele carriers with NO2 exposure have higher risk to current asthma, ever asthma, and injury from air pollution compared to major allele carriers | [131] |
| rs1695 | A/G* | Minor allele carriers for both rs1138272 and rs1695 exposed to traffic and NO2 may be at an increased risk of asthma, current asthma, and ever asthma. *A/A genotype has protective properties against complications from household allergens, G allele makes asthmatics more prone to complications | [131,132] | |||
| NFE2L2 | Candidate gene | Hungarian | rs2588882 | T/G | Protective effect during infection-induced exacerbation in asthmatics [IIA] | [148] |
| rs6721961 | G/T | |||||
| TLR2 | Candidate gene | Netherlands | rs4696480 | T/A | Modifies effect of exposure of air pollution on asthma from birth to 8 years of age | [134] |
| rs1898830 | A/G | |||||
| TLR4 | Candidate gene | Netherlands | rs2770150 | A/G | Modify the effect of exposure to air pollution on asthma | [134] |
| rs10759931 | G/A | |||||
| rs6478317 | A/G | |||||
| rs1927911 | C/A | |||||
| ORMDL3 | Candidate gene | Danish, delivered in Copenhagen | rs7216389 | T/C | Significant association with risk of recurrent wheeze in homozygous variant allele carriers | [143] |
| IL9 | GWAS | Mixed | rs11741137 | C/T | Significant interactions of the rs11741137, rs2069885, and rs1859430 SNPs with dust mite allergen exposure. Increase in the likelihood of exacerbation for dominant genotype carriers [rs11741137, rs2069885] and with increased dust mite exposure | [141] |
| rs2069885 | G/A | |||||
| rs1859430 | G/A | |||||
| P2RY12 | Candidate gene | Mixed | rs8180086 | G/A | Modified airway hyperresponsiveness values with HDM exposure | [145] |
| rs3732765 | G/A | |||||
| rs10935840 | A/G | |||||
| rs11708767 | G/A | |||||
| rs7615865 | A/T | Modified bronchodilator reversibility with HDM exposure | ||||
| rs1491978 | G/C | |||||
| IL6 | Chinese | rs1800795 | G/C | Gene-smoking environmental interaction increases risk of asthma | [149] | |
| IL1RN | Candidate gene | Mixed [150] or Caucasian [151] | rs2234678 | A/G | Protective properties in the development of asthma in those without childhood environmental tobacco exposure; those with childhood exposure, were at higher risk of early onset asthma. Maternal smoking during pregnancy in infants with the GG genotype had increased susceptibility to repeated asthma and persistent asthma in children | [150,151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranjbar, M.; Whetstone, C.E.; Omer, H.; Power, L.; Cusack, R.P.; Gauvreau, G.M. The Genetic Factors of the Airway Epithelium Associated with the Pathology of Asthma. Genes 2022, 13, 1870. https://doi.org/10.3390/genes13101870
Ranjbar M, Whetstone CE, Omer H, Power L, Cusack RP, Gauvreau GM. The Genetic Factors of the Airway Epithelium Associated with the Pathology of Asthma. Genes. 2022; 13(10):1870. https://doi.org/10.3390/genes13101870
Chicago/Turabian StyleRanjbar, Maral, Christiane E. Whetstone, Hafsa Omer, Lucy Power, Ruth P. Cusack, and Gail M. Gauvreau. 2022. "The Genetic Factors of the Airway Epithelium Associated with the Pathology of Asthma" Genes 13, no. 10: 1870. https://doi.org/10.3390/genes13101870