
DDT Resistance in Anopheles pharoensis from Northern Cameroon Associated with High Cuticular Hydrocarbon Production

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Sites
2.2. Mosquito Collection, Rearing and Conservation
2.3. Insecticide Bioassay
2.4. Genomic DNA Extraction from Individual Mosquitoes
2.5. Total Nucleic Acid (NA) Extraction from Mosquito Pools
2.6. Species Identification
2.7. kdr Detection in Anopheles Mosquitoes
2.8. Cuticular Hydrocarbons (CHCs) Identification and Quantitation by GC-MS and GC-FID
3. Results
3.1. Species Identification
3.2. Bioassay Analysis
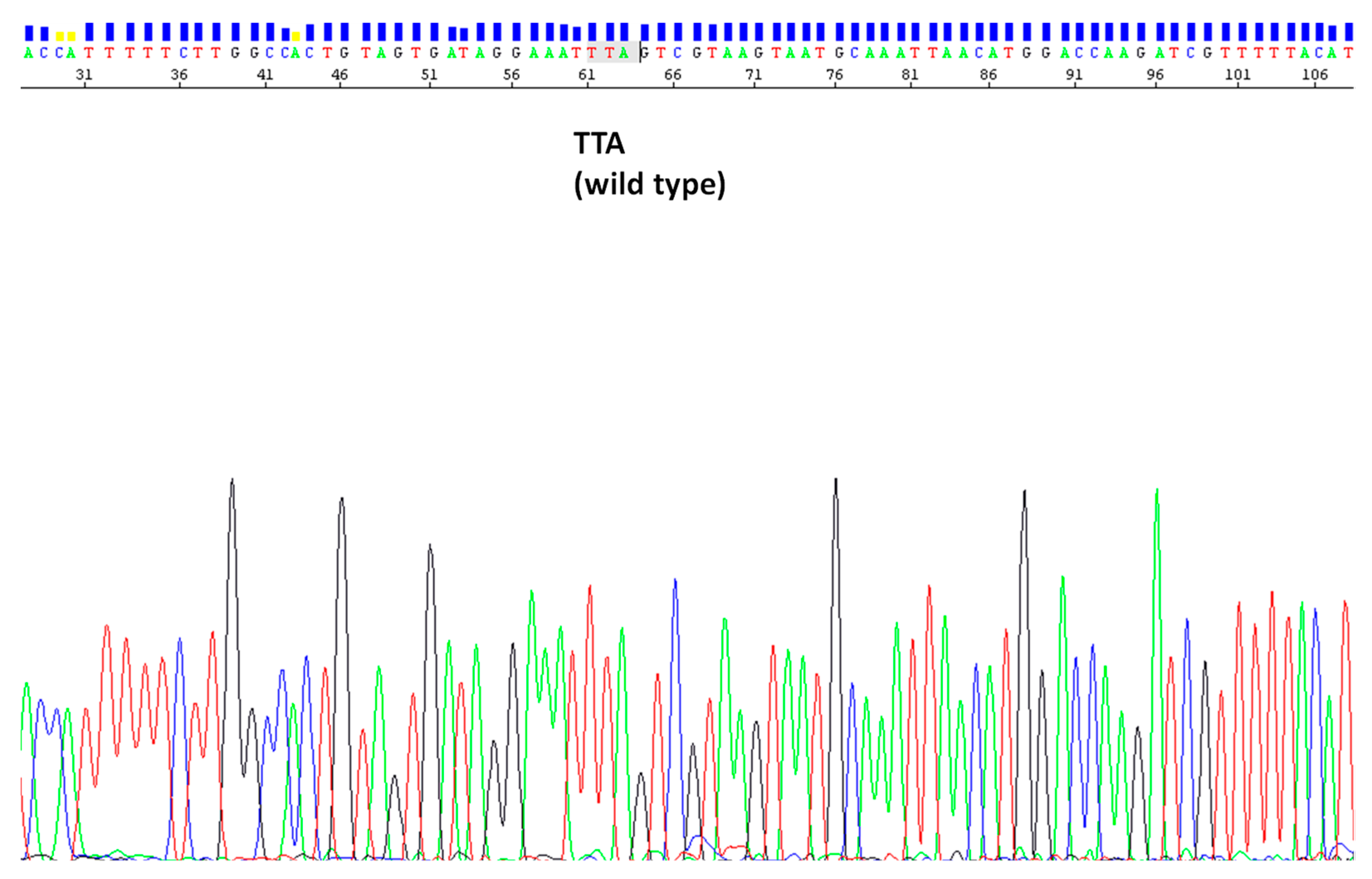
3.3. kdr L1014F/S Analysis in An. pharoensis Samples
3.4. Analysis of Amount of Cuticular Hydrocarbon (CHC) in Mosquito
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2019; World Health Organization: Geneva Swizerland, 2019. [Google Scholar]
- Antonio-Nkondjio, C.; Ndo, C.; Njiokou, F.; Bigoga, J.D.; Awono-Ambene, P.; Etang, J.; Ekobo, A.S.; Wondji, C.S. Review of malaria situation in Cameroon: Technical viewpoint on challenges and prospects for disease elimination. Parasit. Vectors 2019, 12, 501. Available online: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-019-3753-8 (accessed on 29 January 2021). [CrossRef] [PubMed]
- Antonio-Nkondjio, C.; Simard, F.D.R.; Awono-Ambene, P.; Chouaibou, M.; Tchuinkam, T.O.; Fontenille, D. Complexity of the Malaria Vectorial System in Cameroon: Contribution of Secondary Vectors to Malaria Transmission. J. Med. Entomol. 2006, 43, 7. [Google Scholar] [CrossRef]
- Antonio-Nkondjio, C.; Atangana, J.; Ndo, C.; Awono-Ambene, P.; Fondjo, E.; Fontenille, D.; Simard, F. Malaria transmission and rice cultivation in Lagdo, northern Cameroon. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 352–359. Available online: https://www.sciencedirect.com/science/article/pii/S0035920307004415 (accessed on 25 May 2022). [CrossRef] [PubMed]
- Gillies, M.T.; De Meillon, B. The Anophelinae of Africa South of the Sahara (Ethiopian Zoogeographical Region). Anophelinae Afr South Sahara Ethiop Zoogeographical Reg. Johannesburg: South African Institute for Medical Research, P.O. Box 1038, S. Africa. 1968. Available online: https://www.cabdirect.org/cabdirect/abstract/19692900946 (accessed on 25 May 2022).
- Awono-Ambene, P.H.; Etang, J.; Antonio-Nkondjio, C.; Ndo, C.; Eyisap, W.E.; Piameu, M.C.; Mandeng, E.S.; Mbakop, R.L.; Toto, J.C.; Patchoke, S.; et al. The bionomics of the malaria vector Anopheles rufipes Gough, 1910 and its susceptibility to deltamethrin insecticide in North Cameroon. Parasit. Vectors 2018, 11, 253. Available online: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-018-2809-5 (accessed on 24 May 2022). [CrossRef] [PubMed]
- Ibrahim, S.S.; Mukhtar, M.M.; Irving, H.; Labbo, R.; Kusimo, M.O.; Mahamadou, I.; Wondji, C.S. High Plasmodium infection and multiple insecticide resistance in a major malaria vector Anopheles coluzzii from Sahel of Niger Republic. Malar. J. 2019, 18, 181. Available online: https://malariajournal.biomedcentral.com/articles/10.1186/s12936-019-2812-0 (accessed on 3 June 2022). [CrossRef] [PubMed]
- Kerah-Hinzoumbé, C.; Péka, M.; Antonio-Nkondjio, C.; Donan-Gouni, I.; Awono-Ambene, P.; Samè-Ekobo, A.; Simard, F. Malaria vectors and transmission dynamics in Goulmoun, a rural city in south-western Chad. BMC Infect Dis. BioMed Cent. 2009, 9, 1–11. Available online: https://bmcinfectdis.biomedcentral.com/articles/10.1186/1471-2334-9-71 (accessed on 31 July 2022).
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Rubio-Palis, Y.; Chareonviriyaphap, T.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; et al. A global map of dominant malaria vectors. Parasit. Vectors 2012, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Jones, C. The possibility of active long-distance migrations by Anopheles pharoensis Theobald. Bull. World Health Organ. 1962, 27, 299–302. Available online: https://apps.who.int/iris/handle/10665/267602 (accessed on 13 August 2022). [PubMed]
- Balkew, M.; Elhassan, I.; Ibrahim, M.; Gebre-Michael, T.; Engers, H. Very high DDT-resistant population of Anopheles pharoensis Theobald (Diptera: Culicidae) from Gorgora, Northern Ethiopia. Parasite 2007, 13, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Bamou, R.; Kopya, E.; Djamouko-Djonkam, L.; Awono-Ambene, P.; Tchuinkam, T.; Njiokou, F.; Antonio-Nkondjio, C. Assessment of the Anophelinae blood seeking bionomic and pyrethroids resistance of local malaria vectors in the forest region of Southern Cameroon. JEZS 2020, 8, 1054–1062. [Google Scholar]
- Rapport 2016 sur le Paludisme dans le Monde–OMS–ReMed. Available online: http://remed.org/ressources/1084/ (accessed on 27 July 2022).
- Tabue, R.N.; Awono-Ambene, P.; Etang, J.; Atangana, J.; C, A.-N.; Toto, J.C.; Patchoke, S.; Leke, R.G.; Fondjo, E.; Mnzava, A.P.; et al. Role of Anopheles (Cellia) rufipes (Gough, 1910) and other local anophelines in human malaria transmission in the northern savannah of Cameroon: A cross-sectional survey. Parasit. Vectors 2017, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, J.; Wilkerson, R.C.; Besansky, N.J. Evolution of mitochondrial and ribosomal gene sequences in anophelinae (Diptera: Culicidae): Implications for phylogeny reconstruction. Mol. Phylogenet. Evol. 2001, 18, 479–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 associated with insecticide resistance catalyzes cuticular hydrocarbon production in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Nkondjio, C.; Fossog, B.T.; Kopya, E.; Poumachu, Y.; Djantio, B.M.; Ndo, C.; Tchuinkam, T.; Awono-Ambene, P.; Wondji, C.S. Rapid evolution of pyrethroid resistance prevalence in Anopheles gambiae populations from the cities of Douala and Yaoundé (Cameroon). Malar. J. 2015, 14, 155. [Google Scholar] [CrossRef] [PubMed]
- Sonhafouo-Chiana, N.; Nkahe, L.D.; Kopya, E.; Awono-Ambene, P.H.; Wanji, S.; Wondji, C.S.; Antonio-Nkondjio, C. Rapid evolution of insecticide resistance and patterns of pesticides usage in agriculture in the city of Yaoundé, Cameroon. Parasit. Vectors 2022, 15, 186. Available online: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-022-05321-8 (accessed on 27 July 2022). [CrossRef] [PubMed]
- Menze, B.D.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Riveron, J.M.; Wondji, C.S. Bionomics and insecticides resistance profiling of malaria vectors at a selected site for experimental hut trials in central Cameroon. Malar. J. 2018, 17, 317. Available online: https://malariajournal.biomedcentral.com/articles/10.1186/s12936-018-2467-2 (accessed on 23 May 2022). [CrossRef] [PubMed]
- Bamou, R.; Kopya, E.; Nkahe, L.D.; Menze, B.D.; Awono-Ambene, P.; Tchuinkam, T.; Njiokou, F.; Wondji, C.S.; Antonio-Nkondjio, C. Increased prevalence of insecticide resistance in Anopheles coluzzii populations in the city of Yaoundé, Cameroon and influence on pyrethroid-only treated bed net efficacy. Parasite 2021, 28, 8. Available online: https://www.parasite-journal.org/10.1051/parasite/2021003 (accessed on 22 May 2022). [CrossRef] [PubMed]
- Talipouo, A.; Mavridis, K.; Nchoutpouen, E.; Djiappi-Tchamen, B.; Fotakis, E.A.; Kopya, E.; Bamou, R.; Kekeunou, S.; Awono-Ambene, P.; Balabanidou, V.; et al. High insecticide resistance mediated by different mechanisms in Culex quinquefasciatus populations from the city of Yaoundé, Cameroon. Sci. Rep. 2021, 11, 7322. Available online: http://www.nature.com/articles/s41598-021-86850-7 (accessed on 23 May 2022). [CrossRef] [PubMed]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect. Sci. 2018, 27, 68–74. Available online: https://linkinghub.elsevier.com/retrieve/pii/S2214574517301190 (accessed on 6 June 2022). [CrossRef] [PubMed]
- Hemingway, J.; Ranson, H. Insecticide Resistance in Insect Vectors of Human Disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Population | N | Species ID |
|---|---|---|
| Yangah | 10 | 100% An. pharoensis |
| Simatou | 10 | 100% An. pharoensis |
| Maga | 10 | 100% An. pharoensis |
| Sites/Strain | Species | Tested | Dead | Mortality Rate (95% CI) |
|---|---|---|---|---|
| Maga | An. pharoensis | 86 | 54 | 62.79% (47.2–81.9) |
| Simatou | An. pharoensis | 90 | 69 | 76.67% (59.65–97.03) |
| Yangah | An. pharoensis | 25 | 20 | 80% (48.87–123.55) |
| Kisumu | An. gambiae | 100 | 100 | 100% |
| Population | Sample Size (Alleles) | Phenotype | kdr L1014F/S Genotype |
|---|---|---|---|
| Yangah | 20 | DDT resistant | Wild type (TTA or TTG) |
| Simatou | 20 | DDT resistant | Wild type (TTA or TTG) |
| Maga | 74 | DDT resistant | Wild type (TTA or TTG) |
| Kisumu | 30 | susceptible strain | Wild type (TTA or TTG) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kala-Chouakeu, N.A.; Kopya, E.; Balabanidou, V.; Djiappi, B.T.; Papapostolou, K.M.; Tchuinkam, T.; Antonio-Nkondjio, C. DDT Resistance in Anopheles pharoensis from Northern Cameroon Associated with High Cuticular Hydrocarbon Production. Genes 2022, 13, 1723. https://doi.org/10.3390/genes13101723
Kala-Chouakeu NA, Kopya E, Balabanidou V, Djiappi BT, Papapostolou KM, Tchuinkam T, Antonio-Nkondjio C. DDT Resistance in Anopheles pharoensis from Northern Cameroon Associated with High Cuticular Hydrocarbon Production. Genes. 2022; 13(10):1723. https://doi.org/10.3390/genes13101723
Chicago/Turabian StyleKala-Chouakeu, Nelly Armanda, Edmond Kopya, Vasileia Balabanidou, Borel Tchamen Djiappi, Kyriaki Maria Papapostolou, Timoléon Tchuinkam, and Christophe Antonio-Nkondjio. 2022. "DDT Resistance in Anopheles pharoensis from Northern Cameroon Associated with High Cuticular Hydrocarbon Production" Genes 13, no. 10: 1723. https://doi.org/10.3390/genes13101723