
Genomic Predictors of Brisk Walking Are Associated with Elite Sprinter Status

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Participants
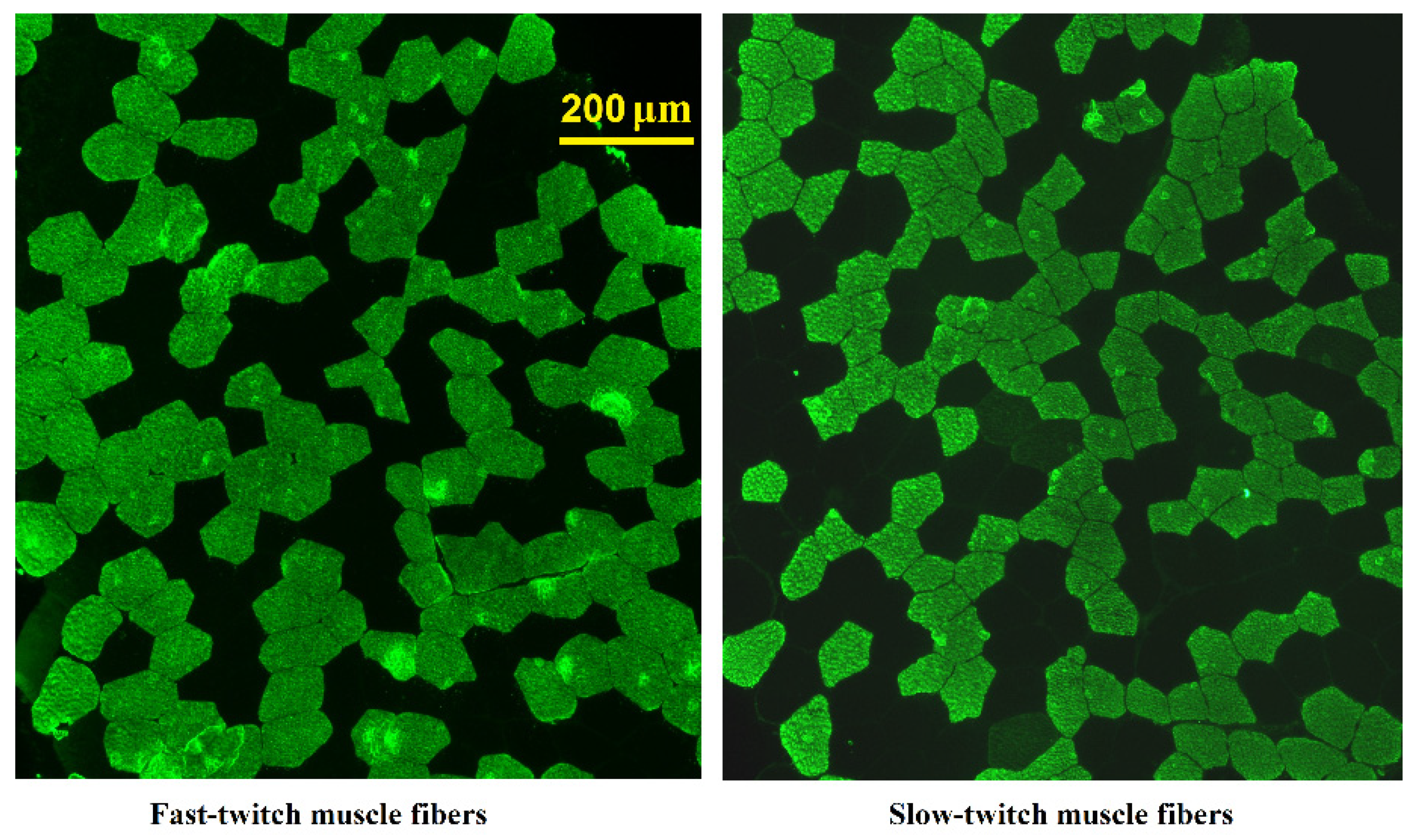
2.3. Evaluation of Muscle Fiber Composition by Immunohistochemistry
2.4. Genotyping
2.5. Statistical Analyses
3. Results
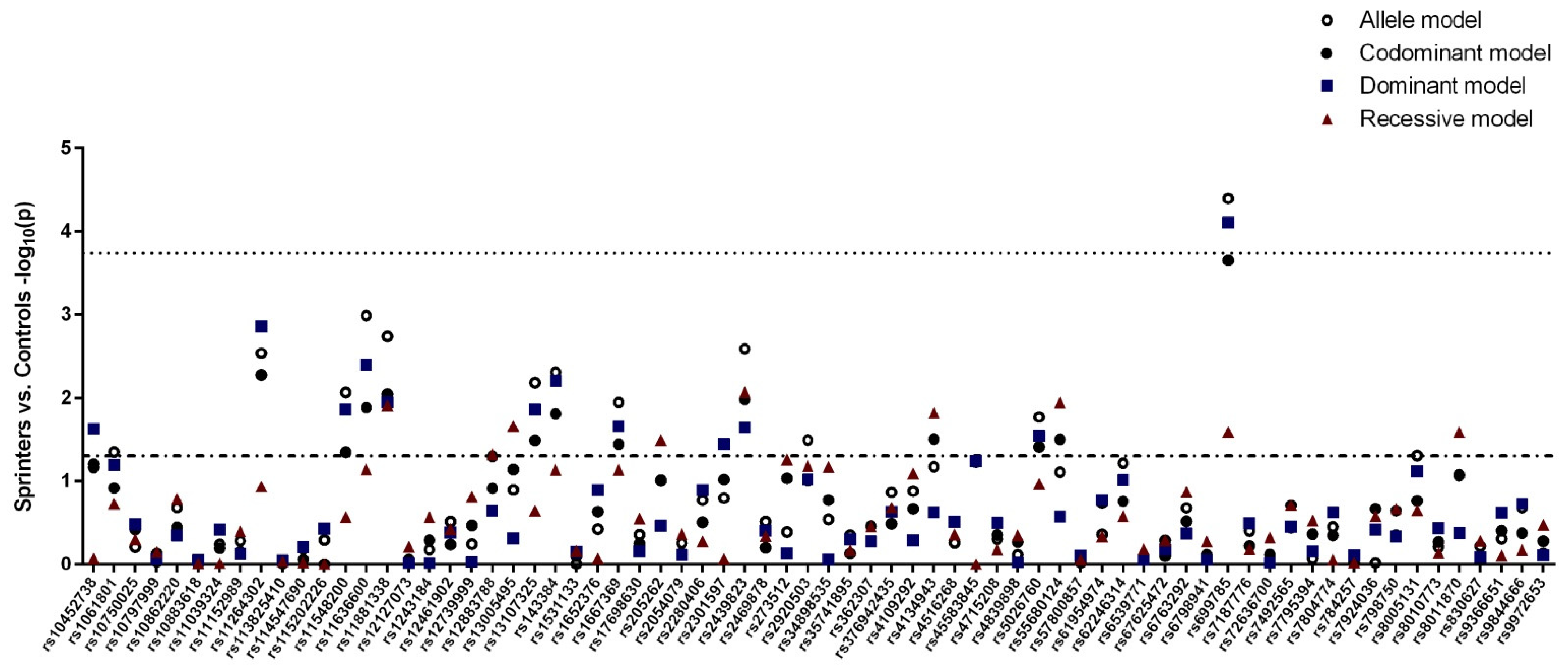
3.1. Case–Control Study
3.2. Polygenic Analysis
3.3. Muscle Biopsy Study
3.4. Bioinformatical Analysis of 15 SNPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yates, T.; Zaccardi, F.; Dhalwani, N.N.; Davies, M.J.; Bakrania, K.; Celis-Morales, C.A.; Gill, J.; Franks, P.W.; Khunti, K. Association of walking pace and handgrip strength with all-cause, cardiovascular, and cancer mortality: A UK biobank observational study. Eur. Heart J. 2017, 38, 3232–3240. [Google Scholar] [CrossRef]
- Celis-Morales, C.A.; Gray, S.; Petermann, F.; Iliodromiti, S.; Welsh, P.; Lyall, D.M.; Anderson, J.; Pellicori, P.; Mackay, D.F.; Pell, J.P.; et al. Walking pace is associated with lower risk of all-cause and cause-specific mortality. Med. Sci. Sports Exerc. 2019, 51, 472–480. [Google Scholar] [CrossRef]
- Bohannon, R.W. Comfortable and maximum walking speed of adults aged 20–79 years: Reference values and determinants. Age Ageing 1997, 26, 15–19. [Google Scholar] [CrossRef]
- Timmins, I.R.; Zaccardi, F.; Nelson, C.P.; Franks, P.W.; Yates, T.; Dudbridge, F. Genome-wide association study of self-reported walking pace suggests beneficial effects of brisk walking on health and survival. Commun. Biol. 2020, 3, 634. [Google Scholar] [CrossRef]
- Fragala, M.S.; Alley, D.E.; Shardell, M.D.; Harris, T.B.; McLean, R.R.; Kiel, D.P.; Cawthon, P.M.; Dam, T.T.; Ferrucci, L.; Guralnik, J.M.; et al. Comparison of handgrip and leg extension strength in predicting slow gait speed in older adults. J. Am. Geriatr. Soc. 2016, 64, 144–150. [Google Scholar] [CrossRef]
- Ossowski, Z.M.; Wiech, M.; Ellwart, M. Association between gait speed and muscle mass and strength in postmenopausal women. Balt. J. Health Phys. Act. 2019, 11, 106–114. [Google Scholar] [CrossRef]
- Schlicht, J.; Camaione, D.N.; Owen, S.V. Effect of intense strength training on standing balance, walking speed, and sit-to-stand performance in older adults. J. Gerontol. A Biol. Sci. Med. Sci. 2001, 56, 281–286. [Google Scholar] [CrossRef]
- Morin, J.B.; Bourdin, M.; Edouard, P.; Peyrot, N.; Samozino, P.; Lacour, J.R. Mechanical determinants of 100-m sprint running performance. Eur. J. Appl. Physiol. 2012, 112, 3921–3930. [Google Scholar] [CrossRef]
- Haugen, T.; McGhie, D.; Ettema, G. Sprint running: From fundamental mechanics to practice-a review. Eur. J. Appl. Physiol. 2019, 119, 1273–1287. [Google Scholar] [CrossRef]
- Ginevičienė, V.; Utkus, A.; Pranckevičienė, E.; Semenova, E.A.; Hall, E.; Ahmetov, I.I. Perspectives in sports genomics. Biomedicines 2022, 10, 298. [Google Scholar] [CrossRef]
- Ahmetov, I.; Kulemin, N.; Popov, D.; Naumov, V.; Akimov, E.; Bravy, Y.; Egorova, E.; Galeeva, A.; Generozov, E.; Kostryukova, E.; et al. Genome-wide association study identifies three novel genetic markers associated with elite endurance performance. Biol. Sport 2015, 32, 3–9. [Google Scholar] [CrossRef]
- Al-Khelaifi, F.; Yousri, N.A.; Diboun, I.; Semenova, E.A.; Kostryukova, E.S.; Kulemin, N.A.; Borisov, O.V.; Andryushchenko, L.B.; Larin, A.K.; Generozov, E.V.; et al. Genome-wide association study reveals a novel association between MYBPC3 gene polymorphism, endurance athlete status, aerobic capacity and steroid metabolism. Front. Genet. 2020, 11, 595. [Google Scholar] [CrossRef]
- Pickering, C.; Suraci, B.; Semenova, E.A.; Boulygina, E.A.; Kostryukova, E.S.; Kulemin, N.A.; Borisov, O.V.; Khabibova, S.A.; Larin, A.K.; Pavlenko, A.V.; et al. A genome-wide association study of sprint performance in elite youth football players. J. Strength Cond. Res. 2019, 33, 2344–2351. [Google Scholar] [CrossRef]
- Rankinen, T.; Fuku, N.; Wolfarth, B.; Wang, G.; Sarzynski, M.A.; Alexeev, D.G.; Ahmetov, I.I.; Boulay, M.R.; Cieszczyk, P.; Eynon, N.; et al. No evidence of a common DNA variant profile specific to world class endurance athletes. PLoS ONE 2016, 11, e0147330. [Google Scholar] [CrossRef]
- Guilherme, J.; Semenova, E.A.; Zempo, H.; Martins, G.L.; Lancha Junior, A.H.; Miyamoto-Mikami, E.; Kumagai, H.; Tobina, T.; Shiose, K.; Kakigi, R.; et al. Are genome-wide association study identified single-nucleotide polymorphisms associated with sprint athletic status? A replication study with 3 different cohorts. Int. J. Sports Physiol. Perform. 2021, 16, 489–495. [Google Scholar] [CrossRef]
- Malczewska-Lenczowska, J.; Orysiak, J.; Majorczyk, E.; Sitkowski, D.; Starczewski, M.; Zmijewski, P. HIF-1α and NFIA-AS2 Polymorphisms as Potential Determinants of Total Hemoglobin Mass in Endurance Athletes. J. Strength Cond. Res. 2022, 36, 1596–1604. [Google Scholar] [CrossRef]
- Kikuchi, N.; Moreland, E.; Homma, H.; Semenova, E.A.; Saito, M.; Larin, A.K.; Kobatake, N.; Yusupov, R.A.; Okamoto, T.; Nakazato, K.; et al. Genes and weightlifting performance. Genes 2022, 13, 25. [Google Scholar] [CrossRef]
- Williams, A.G.; Folland, J.P. Similarity of polygenic profiles limits the potential for elite human physical performance. J. Physiol. 2008, 586, 113–121. [Google Scholar] [CrossRef]
- Li, S.; Zhang, C.; Takemori, H.; Zhou, Y.; Xiong, Z.Q. TORC1 regulates activity-dependent CREB-target gene transcription and dendritic growth of developing cortical neurons. J. Neurosci. 2009, 29, 2334–2343. [Google Scholar] [CrossRef]
- Syed, A.; Lukacsovich, T.; Pomeroy, M.; Bardwell, A.J.; Decker, G.T.; Waymire, K.G.; Purcell, J.; Huang, W.; Gui, J.; Padilla, E.M.; et al. Miles to go (mtgo) encodes FNDC3 proteins that interact with the chaperonin subunit CCT3 and are required for NMJ branching and growth in Drosophila. Dev. Biol. 2019, 445, 37–53. [Google Scholar] [CrossRef]
- Berdeaux, R.; Hutchins, C. Anabolic and Pro-metabolic Functions of CREB-CRTC in Skeletal Muscle: Advantages and Obstacles for Type 2 Diabetes and Cancer Cachexia. Front. Endocrinol. 2019, 10, 535. [Google Scholar]
- Kim, H.R.; Rahman, F.U.; Kim, K.S.; Kim, E.K.; Cho, S.M.; Lee, K.; Moon, O.S.; Seo, Y.W.; Yoon, W.K.; Won, Y.S.; et al. Critical roles of E2F3 in growth and musculo-skeletal phenotype in mice. Int. J. Med. Sci. 2019, 16, 1557–1563. [Google Scholar] [PubMed]
- Vissing, K.; Schjerling, P. Simplified data access on human skeletal muscle transcriptome responses to differentiated exercise. Sci. Data 2014, 1, 140041. [Google Scholar]
- Rundqvist, H.C.; Montelius, A.; Osterlund, T.; Norman, B.; Esbjornsson, M.; Jansson, E. Acute sprint exercise transcriptome in human skeletal muscle. PLoS ONE 2019, 14, e0223024. [Google Scholar] [CrossRef]
- Tikkanen, E.; Gustafsson, S.; Amar, D.; Shcherbina, A.; Waggott, D.; Ashley, E.A.; Ingelsson, E. Biological insights into muscular strength: Genetic findings in the UK Biobank. Sci. Rep. 2018, 8, 6451. [Google Scholar] [CrossRef]
- Moreland, E.; Borisov, O.V.; Semenova, E.A.; Larin, A.K.; Andryushchenko, O.N.; Andryushchenko, L.B.; Generozov, E.V.; Williams, A.G.; Ahmetov, I.I. Polygenic profile of elite strength athletes. J. Strength Cond. Res. 2022, 36, 2509–2514. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhang, M.; Tian, A.; Chen, L.; Sun, Z.; Wang, L.; Chen, P. Exploring the genetic correlation between obesity-related traits and regional brain volumes: Evidence from UK biobank cohort. Neuroimage Clin. 2021, 33, 102870. [Google Scholar] [CrossRef]
- Ahmad, S.; Poveda, A.; Shungin, D.; Barroso, I.; Hallmans, G.; Renström, F.; Franks, P.W. Established bmi-associated genetic variants and their prospective associations with BMI and other cardiometabolic traits: The glacier study. Int. J. Obes. 2016, 40, 1346–1352. [Google Scholar]
- Riveros-McKay, F.; Mistry, V.; Bounds, R.; Hendricks, A.; Keogh, J.M.; Thomas, H.; Henning, E.; Corbin, L.J.; Understanding Society Scientific Group; O’Rahilly, S.; et al. Genetic architecture of human thinness compared to severe obesity. PLoS Genet. 2019, 15, e1007603. [Google Scholar]
- UKB Neale v2. UK Biobank Gwas, Round 2 Results . [Released 1st august 2018]. 2018. Available online: http://www.nealelab.is/uk-biobank/ (accessed on 6 September 2022).
- Peng, L.; Jin, S.; Lu, J.; Ouyang, C.; Guo, J.; Xie, Z.; Shen, H.; Wang, P. Association between growth differentiation factor 5 rs143383 genetic polymorphism and the risk of knee osteoarthritis among Caucasian but not Asian: A meta-analysis. Arthritis Res. Ther. 2020, 22, 215. [Google Scholar] [CrossRef]
- Hatazawa, Y.; Ono, Y.; Hirose, Y.; Kanai, S.; Fujii, N.L.; Machida, S.; Nishino, I.; Shimizu, T.; Okano, M.; Kamei, Y.; et al. Reduced dnmt3a increases gdf5 expression with suppressed satellite cell differentiation and impaired skeletal muscle regeneration. Faseb J. 2018, 32, 1452–1467. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Chang, Q.; Huang, T.; Huang, C. Prospective cohort study of the risk factors for stress fractures in chinese male infantry recruits. J. Int. Med. Res. 2016, 44, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Stastny, P.; Lehnert, M.; De Ste Croix, M.; Petr, M.; Svoboda, Z.; Maixnerova, E.; Varekova, R.; Botek, M.; Petrek, M.; Kocourkova, L.; et al. Effect of COL5A1, GDF5, and PPARA genes on a movement screen and neuromuscular performance in adolescent team sport athletes. J. Strength Cond. Res. 2019, 33, 2057–2065. [Google Scholar] [CrossRef]
- Guilherme, J.; Lancha, A.H., Jr. Total genotype score and athletic status: An exploratory cross-sectional study of a Brazilian athlete cohort. Ann. Hum. Genet. 2020, 84, 141–150. [Google Scholar] [CrossRef]
- Wang, G.; Tanaka, M.; Eynon, N.; North, K.N.; Williams, A.G.; Collins, M.; Moran, C.N.; Britton, S.L.; Fuku, N.; Ashley, E.A.; et al. The Future of Genomic Research in Athletic Performance and Adaptation to Training. Med. Sport Sci. 2016, 61, 55–67. [Google Scholar]
- Guilherme, J.P.L.F.; Semenova, E.A.; Borisov, O.V.; Larin, A.K.; Moreland, E.; Generozov, E.V.; Ahmetov, I.I. Genomic predictors of testosterone levels are associated with muscle fiber size and strength. Eur. J. Appl. Physiol. 2022, 122, 415–423. [Google Scholar] [CrossRef]
- Maciejewska-Skrendo, A.; Sawczuk, M.; Cięszczyk, P.; Ahmetov, I.I. Genes and power athlete status. In Sports, Exercise, and Nutritional Genomics; Academic Press: Cambridge, MA, USA, 2019; pp. 41–72. [Google Scholar]



{kind=link}
{kind=link}
| Consequence | HSWA (%) | p-Value (Allele Model) | ||||||
|---|---|---|---|---|---|---|---|---|
| Chr. | Gene | SNP | Type | HSWA | Sprinters | Controls | Diff. | |
| 1 | ADAM15 | rs11264302 | Intronic | G | 55.8 | 42.9 | 0.129 | 0.003 |
| 1 | CCT3 | rs11548200 | Exonic | T | 95.6 | 89.7 | 0.058 | 0.009 |
| 1 | IGSF3 | rs699785 | Intronic | A | 30.7 | 15.5 | 0.152 | 0.00004 |
| 2 | FHL2 | rs55680124 | ncRNA | C | 88.3 | 82.9 | 0.053 | 0.078 |
| 3 | PPARG | rs2920503 | Intergenic | T | 33.9 | 25.4 | 0.085 | 0.032 |
| 4 | SLC39A8 | rs13107325 | Exonic | C | 97.1 | 91.7 | 0.053 | 0.007 |
| 6 | C6orf106 | rs205262 | Intronic | A | 79.2 | 73.0 | 0.062 | 0.096 |
| 6 | E2F3 | rs4134943 | Intronic | T | 19.0 | 13.1 | 0.059 | 0.067 |
| 7 | AUTS2 | rs10452738 | Intronic | A | 75.5 | 68.3 | 0.072 | 0.063 |
| 10 | CRTAC1 | rs2439823 | Intronic | A | 54.0 | 40.9 | 0.131 | 0.003 |
| 14 | AKAP6 | rs12883788 | Intergenic | C | 56.9 | 48.4 | 0.085 | 0.050 |
| 14 | NRXN3 | rs8011870 | ncRNA | G | 67.2 | 59.9 | 0.073 | 0.085 |
| 19 | CRTC1 | rs11881338 | Intronic | A | 57.7 | 44.0 | 0.137 | 0.002 |
| 19 | ZNF568 | rs1667369 | Intergenic | A | 75.5 | 65.5 | 0.100 | 0.011 |
| 20 | GDF5 | rs143384 | UTR5 | G | 51.1 | 38.9 | 0.122 | 0.005 |
| Alleles | Genotype Distribution (%) | p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | SNP | 1/2 | Group | 1/1 | 1/2 | 2/2 | Cod. | Dom. | Rec. |
| ADAM15 | rs11264302 | A/G | Controls | 35.7 | 42.9 | 21.4 | 0.005 | 0.001 | 0.116 |
| Sprinters | 18.2 | 51.8 | 29.9 | — | — | — | |||
| CCT3 | rs11548200 | T/C | Controls | 81.7 | 15.9 | 2.4 | 0.045 | 0.014 | 0.274 |
| Sprinters | 92.0 | 7.3 | 0.7 | — | — | — | |||
| IGSF3 | rs699785 | G/A | Controls | 71.4 | 26.2 | 2.4 | 0.0002 | 0.0001 | 0.026 |
| Sprinters | 47.4 | 43.8 | 8.8 | — | — | — | |||
| FHL2 | rs55680124 | C/T | Controls | 70.6 | 24.6 | 4.8 | 0.032 | 0.269 | 0.010 |
| Sprinters | 76.6 | 23.4 | 0.0 | — | — | — | |||
| PPARG | rs2920503 | C/T | Controls | 55.6 | 38.1 | 6.3 | 0.098 | 0.095 | 0.065 |
| Sprinters | 45.3 | 41.6 | 13.1 | — | — | — | |||
| SLC39A8 | rs13107325 | C/T | Controls | 84.9 | 13.5 | 1.6 | 0.033 | 0.014 | 0.139 |
| Sprinters | 94.2 | 5.8 | 0.0 | — | — | — | |||
| C6orf106 | rs205262 | A/G | Controls | 56.3 | 33.3 | 10.3 | 0.098 | 0.348 | 0.032 |
| Sprinters | 62.0 | 34.3 | 3.6 | — | — | — | |||
| E2F3 | rs4134943 | C/T | Controls | 73.8 | 26.2 | 0.0 | 0.032 | 0.238 | 0.010 |
| Sprinters | 67.2 | 27.7 | 5.1 | — | — | — | |||
| AUTS2 | rs10452738 | A/G | Controls | 44.4 | 47.6 | 7.9 | 0.068 | 0.024 | 0.846 |
| Sprinters | 58.4 | 34.3 | 7.3 | — | — | — | |||
| CRTAC1 | rs2439823 | G/A | Controls | 34.9 | 48.4 | 16.7 | 0.010 | 0.009 | 0.023 |
| Sprinters | 20.4 | 51.1 | 28.5 | — | — | — | |||
| AKAP6 | rs12883788 | C/T | Controls | 25.4 | 46.0 | 28.6 | 0.121 | 0.230 | 0.048 |
| Sprinters | 32.1 | 49.6 | 18.2 | — | — | — | |||
| NRXN3 | rs8011870 | G/A | Controls | 38.9 | 42.1 | 19.0 | 0.084 | 0.420 | 0.026 |
| Sprinters | 43.8 | 46.7 | 9.5 | — | — | — | |||
| CRTC1 | rs11881338 | T/A | Controls | 31.7 | 48.4 | 19.8 | 0.009 | 0.011 | 0.012 |
| Sprinters | 18.2 | 48.2 | 33.6 | — | — | — | |||
| ZNF568 | rs1667369 | A/C | Controls | 42.1 | 46.8 | 11.1 | 0.036 | 0.022 | 0.073 |
| Sprinters | 56.7 | 38.7 | 5.1 | — | — | — | |||
| GDF5 | rs143384 | A/G | Controls | 38.1 | 46.0 | 15.9 | 0.015 | 0.006 | 0.073 |
| Sprinters | 22.6 | 52.6 | 24.8 | — | — | — | |||
| TGS (70 SNPs) | TGS (15 SNPs) | |||
|---|---|---|---|---|
| Sprinters | Controls | Sprinters | Controls | |
| Number of participants | 137 | 126 | 137 | 126 |
| Minimum score (number of favorable alleles) | 45.7 (64) | 42.9 (60) | 40.0 (12) | 23.3 (7) |
| Maximum score (number of favorable alleles) | 62.9 (88) | 60.0 (84) | 86.7 (26) | 73.3 (22) |
| Mode (number of favorable alleles) | 55.0 (77) | 50.7 (71) | 63.3 (19) | 56.7 (17) |
| Mean (SD) | 53.7 (3.4) | 51.3 (3.8) | 62.5 (7.4) | 53.3 (9.1) |
| Difference between means; p-value | 2.4 ± 0.4; p < 0.0001 | 9.2 ± 1.0; p < 0.0001 | ||
| Skewness (SE) | 0.005 (0.207) | −0.095 (0.216) | −0.024 (0.207) | −0.289 (0.216) |
| Kurtosis (SE) | −0.391 (0.411) | −0.582 (0.428) | 0.811 (0.411) | 0.004 (0.428) |
| Area under the ROC curve (95% CI); p-value | 0.673 (0.608–0.738); p < 0.0001 | 0.781 (0.725–0.837); p < 0.0001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guilherme, J.P.L.F.; Semenova, E.A.; Larin, A.K.; Yusupov, R.A.; Generozov, E.V.; Ahmetov, I.I. Genomic Predictors of Brisk Walking Are Associated with Elite Sprinter Status. Genes 2022, 13, 1710. https://doi.org/10.3390/genes13101710
Guilherme JPLF, Semenova EA, Larin AK, Yusupov RA, Generozov EV, Ahmetov II. Genomic Predictors of Brisk Walking Are Associated with Elite Sprinter Status. Genes. 2022; 13(10):1710. https://doi.org/10.3390/genes13101710
Chicago/Turabian StyleGuilherme, João Paulo L. F., Ekaterina A. Semenova, Andrey K. Larin, Rinat A. Yusupov, Edward V. Generozov, and Ildus I. Ahmetov. 2022. "Genomic Predictors of Brisk Walking Are Associated with Elite Sprinter Status" Genes 13, no. 10: 1710. https://doi.org/10.3390/genes13101710