
Impact of Harvest on Switchgrass Leaf Microbial Communities

 , and
, and 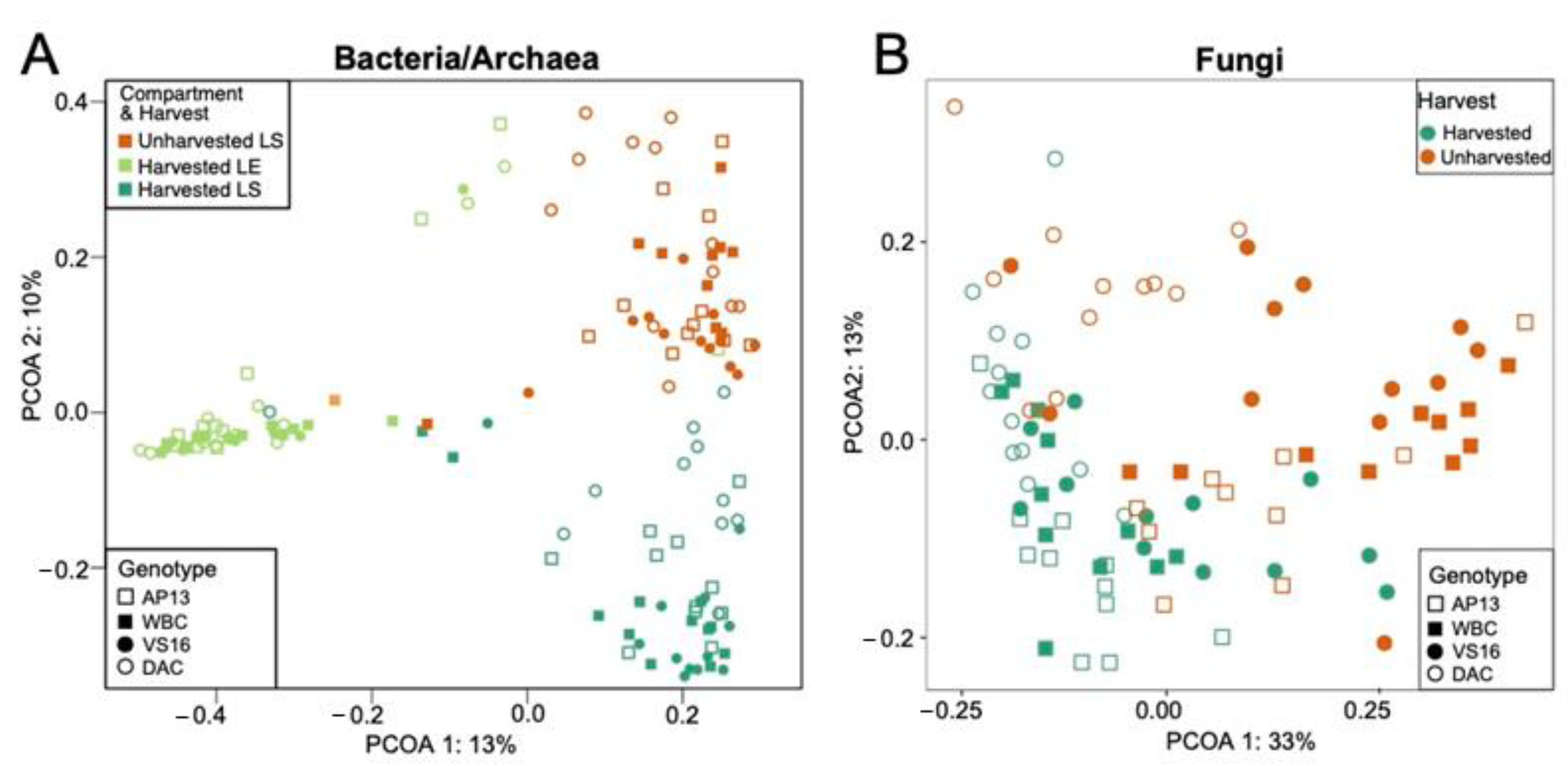{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Switchgrass Plants and Study Location
2.2. Leaf Sampling and Library Preparation
2.3. DNA Extraction, Amplification and Sequencing
2.4. α- and β-Diversity Analyses
3. Results and Discussion
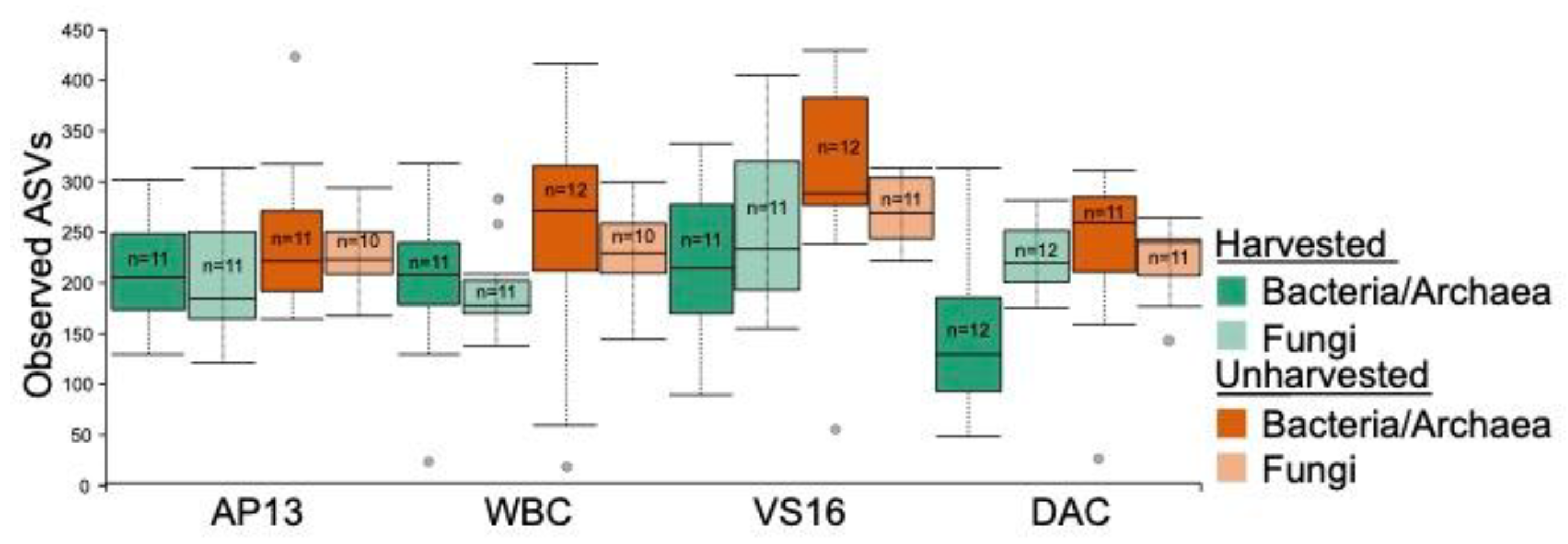
3.1. Sequencing Summary
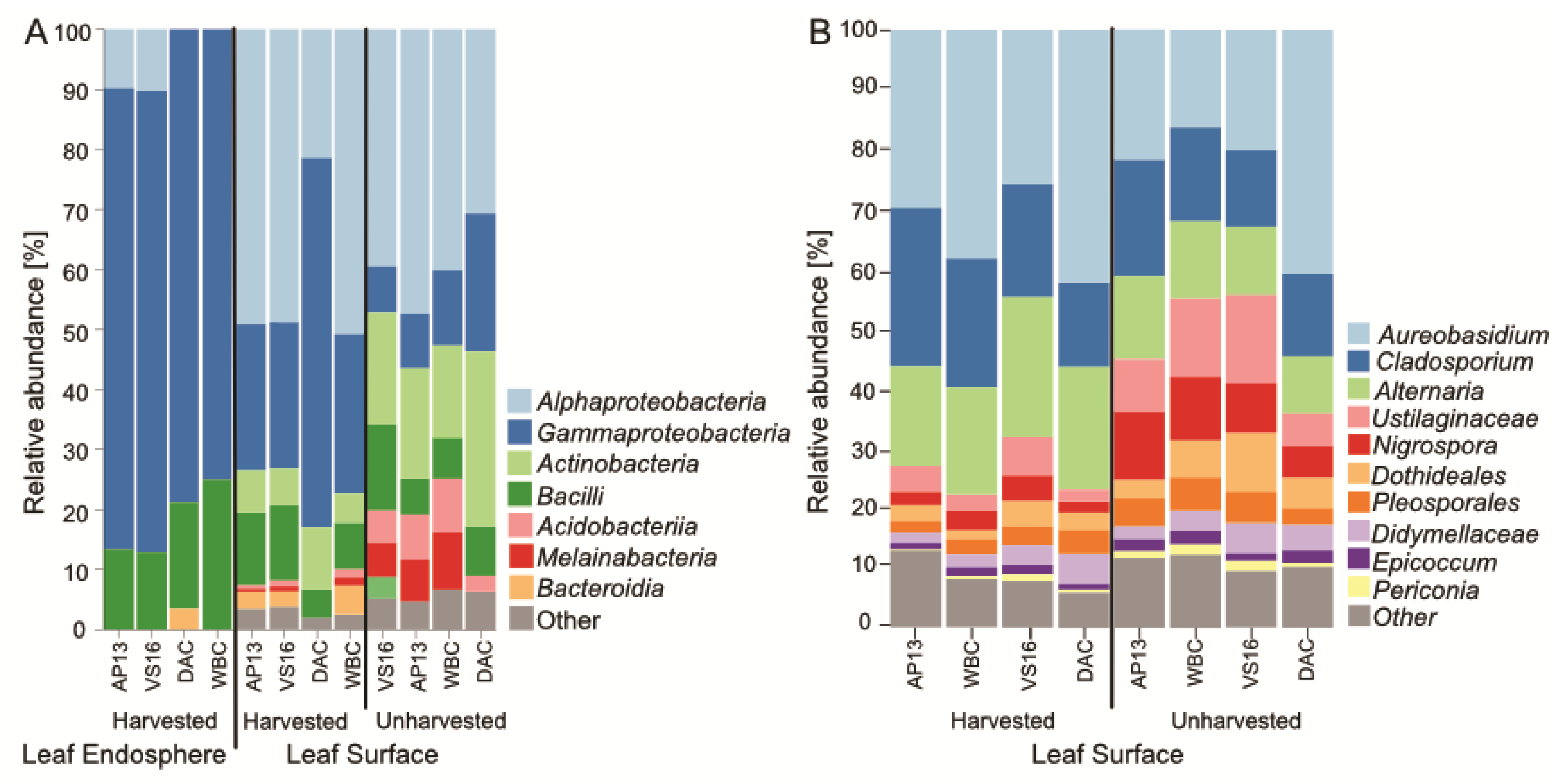
3.2. Microbial Community Assembly Was Impacted by Leaf Compartment, Harvest Treatment and Genotype
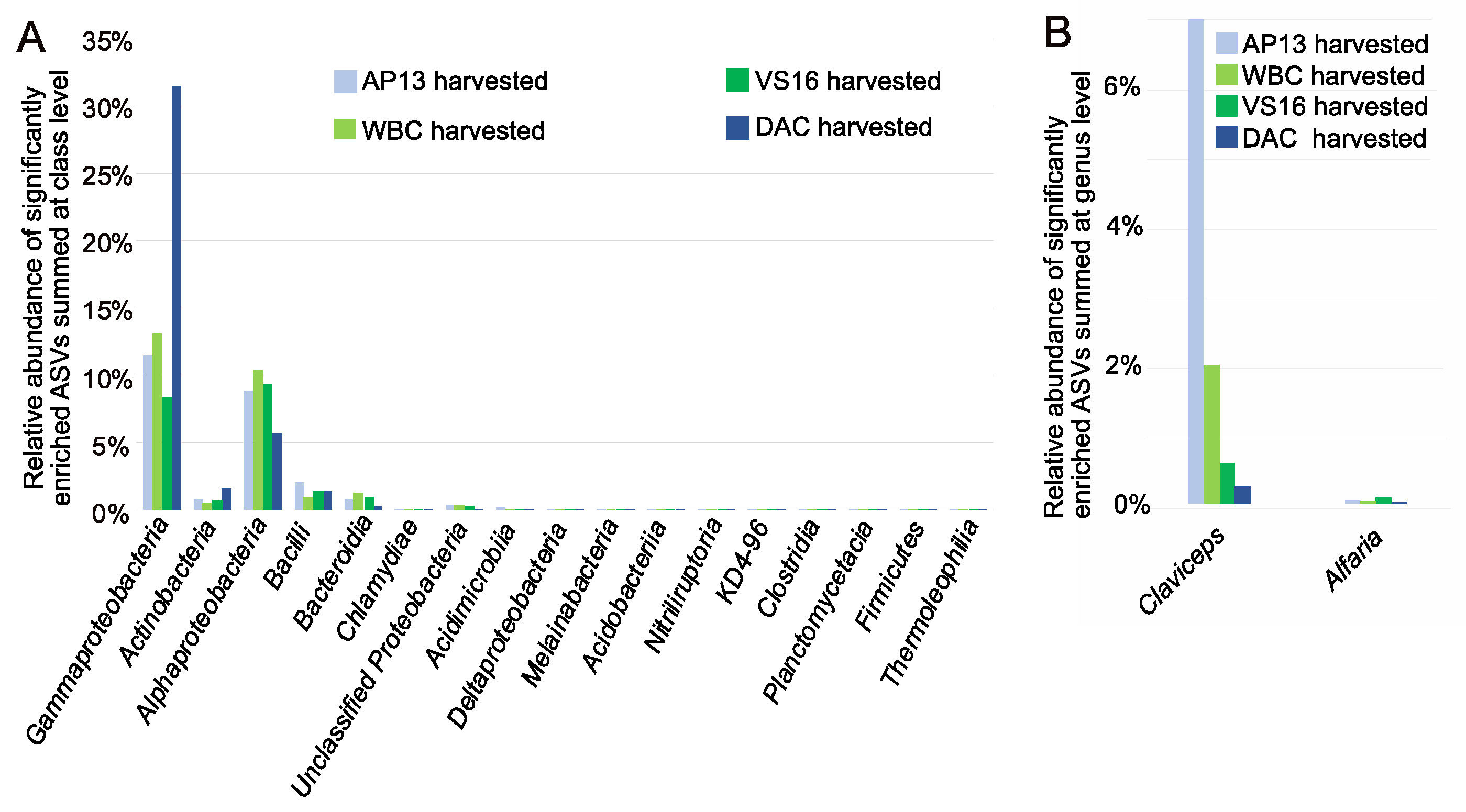
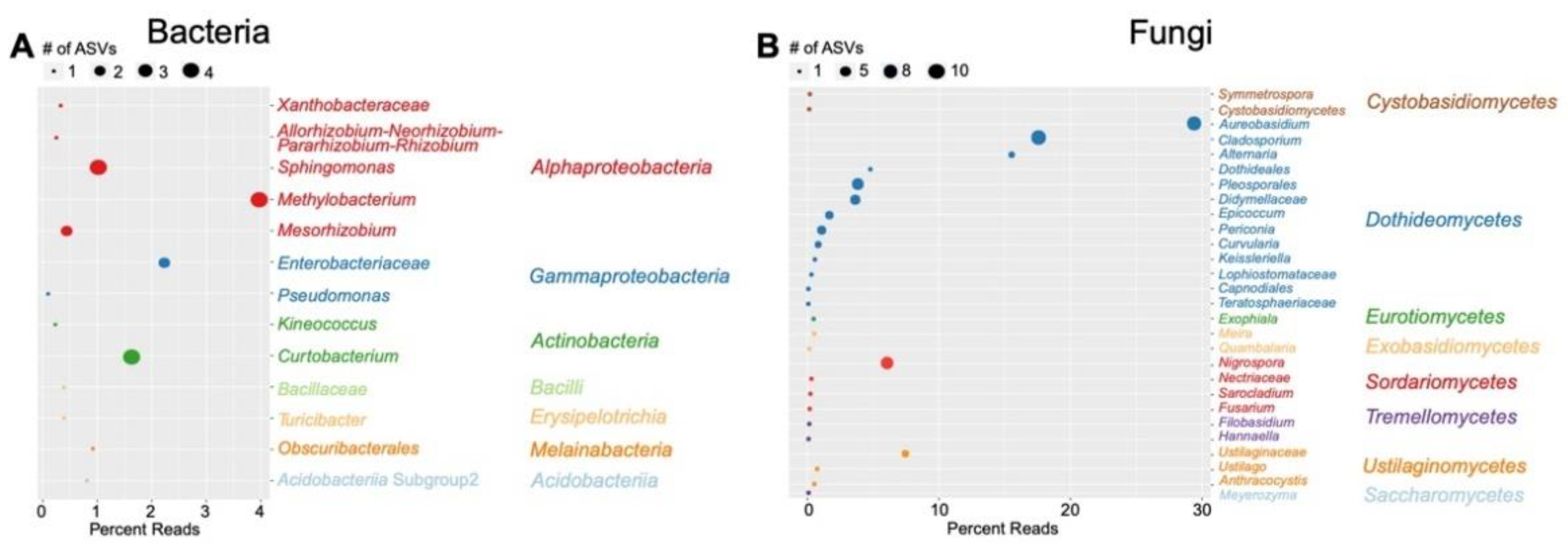
3.3. Differentially Abundant ASVs in Leaves from Harvested Plants Were Related to Pathogens
3.4. The Local Switchgrass Leaf Surface Bacterial Core Was Smaller than the Fungal Core
4. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The Rhizosphere Microbiome and Plant Health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Singer, E.; Bonnette, J.; Kenaley, S.C.; Woyke, T.; Juenger, T.E. Plant Compartment and Genetic Variation Drive Microbiome Composition in Switchgrass Roots. Environ. Microbiol. Rep. 2019, 11, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, K.S.; Snoek, L.B.; Koorem, K.; Geisen, S.; Bloem, L.J.; ten Hooven, F.; Kostenko, O.; Krigas, N.; Manrubia, M.; Caković, D.; et al. Range-Expansion Effects on the Belowground Plant Microbiome. Nat. Ecol. Evol. 2019, 3, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the Soil Type on the Microbiome in the Rhizosphere of Field-Grown Lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Pii, Y.; Borruso, L.; Brusetti, L.; Crecchio, C.; Cesco, S.; Mimmo, T. The Interaction between Iron Nutrition, Plant Species and Soil Type Shapes the Rhizosphere Microbiome. Plant Physiol. Biochem. 2016, 99, 39–48. [Google Scholar] [CrossRef]
- Wallenstein, M.D. Managing and Manipulating the Rhizosphere Microbiome for Plant Health: A Systems Approach. Rhizosphere 2017, 3, 230–232. [Google Scholar] [CrossRef]
- Bano, S.A.; Uzair, B. Arbuscular Mycorrhizal Fungi (AMF) for Improved Plant Health and Production. In Microbial Biotechnology in Crop Protection; Springer: Singapore, 2021; ISBN 978-981-16-0048-7. [Google Scholar]
- Kumar, A.; Dubey, A. Rhizosphere Microbiome: Engineering Bacterial Competitiveness for Enhancing Crop Production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef]
- Tall, S.; Meyling, N.V. Probiotics for Plants? Growth Promotion by the Entomopathogenic Fungus Beauveria Bassiana Depends on Nutrient Availability. Microb. Ecol. 2018, 76, 1002–1008. [Google Scholar] [CrossRef]
- Nieto-Jacobo, M.F.; Steyaert, J.M.; Salazar-Badillo, F.B.; Nguyen, D.V.; Rostás, M.; Braithwaite, M.; De Souza, J.T.; Jimenez-Bremont, J.F.; Ohkura, M.; Stewart, A.; et al. Environmental Growth Conditions of Trichoderma Spp. Affects Indole Acetic Acid Derivatives, Volatile Organic Compounds, and Plant Growth Promotion. Front. Plant Sci. 2017, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Senthil Kumar, C.M.; Jacob, T.K.; Devasahayam, S.; Thomas, S.; Geethu, C. Multifarious Plant Growth Promotion by an Entomopathogenic Fungus Lecanicillium Psalliotae. Microbiol. Res. 2018, 207, 153–160. [Google Scholar] [CrossRef]
- Taghavi, S.; Garafola, C.; Monchy, S.; Newman, L.; Hoffman, A.; Weyens, N.; Barac, T.; Vangronsveld, J.; van der Lelie, D. Genome Survey and Characterization of Endophytic Bacteria Exhibiting a Beneficial Effect on Growth and Development of Poplar Trees. Appl. Environ. Microbiol. 2009, 75, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Fedorov, D.N.; Ekimova, G.A.; Doronina, N.V.; Trotsenko, Y.A. 1-Aminocyclopropane-1-Carboxylate (ACC) Deaminases from Methylobacterium Radiotolerans and Methylobacterium Nodulans with Higher Specificity for ACC. FEMS Microbiol. Lett. 2013, 343, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, M.-Y.; Khan, N.; Tan, L.-L.; Yang, S. Potentials, Utilization, and Bioengineering of Plant Growth-Promoting Methylobacterium for Sustainable Agriculture. Sustainability 2021, 13, 3941. [Google Scholar] [CrossRef]
- Fürnkranz, M.; Wanek, W.; Richter, A.; Abell, G.; Rasche, F.; Sessitsch, A. Nitrogen Fixation by Phyllosphere Bacteria Associated with Higher Plants and Their Colonizing Epiphytes of a Tropical Lowland Rainforest of Costa Rica. ISME J. 2008, 2, 561–570. [Google Scholar] [CrossRef]
- Grady, K.L.; Sorensen, J.W.; Stopnisek, N.; Guittar, J.; Shade, A. Assembly and Seasonality of Core Phyllosphere Microbiota on Perennial Biofuel Crops. Nat. Commun. 2019, 10, 4135. [Google Scholar] [CrossRef] [Green Version]
- Hestrin, R.; Lee, M.R.; Whitaker, B.K.; Pett-Ridge, J. The Switchgrass Microbiome: A Review of Structure, Function, and Taxonomic Distribution. Phytobiomes J. 2021, 5, 14–28. [Google Scholar] [CrossRef]
- Soman, C.; Keymer, D.P.; Kent, A.D. Edaphic Correlates of Feedstock-associated Diazotroph Communities. GCB Bioenergy 2018, 10, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, B.K.; Reynolds, H.L.; Clay, K. Foliar Fungal Endophyte Communities Are Structured by Environment but Not Host Ecotype in Panicum Virgatum (Switchgrass). Ecology 2018, 99, 2703–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodsky, O.L.; Shek, K.L.; Dinwiddie, D.; Bruner, S.G.; Gill, A.S.; Hoch, J.M.; Palmer, M.I.; McGuire, K.L. Microbial Communities in Bioswale Soils and Their Relationships to Soil Properties, Plant Species, and Plant Physiology. Front. Microbiol. 2019, 10, 2368. [Google Scholar] [CrossRef] [PubMed]
- Giauque, H.; Connor, E.W.; Hawkes, C.V. Endophyte Traits Relevant to Stress Tolerance, Resource Use and Habitat of Origin Predict Effects on Host Plants. New Phytol. 2019, 221, 2239–2249. [Google Scholar] [CrossRef]
- Bahulikar, R.A.; Torres-Jerez, I.; Worley, E.; Craven, K.; Udvardi, M.K. Diversity of Nitrogen-Fixing Bacteria Associated with Switchgrass in the Native Tallgrass Prairie of Northern Oklahoma. Appl. Environ. Microbiol. 2014, 80, 5636–5643. [Google Scholar] [CrossRef] [Green Version]
- Roley, S.S.; Xue, C.; Hamilton, S.K.; Tiedje, J.M.; Robertson, G.P. Isotopic Evidence for Episodic Nitrogen Fixation in Switchgrass (Panicum virgatum L.). Soil Biol. Biochem. 2019, 129, 90–98. [Google Scholar] [CrossRef]
- Chen, H.; Yang, Z.K.; Yip, D.; Morris, R.H.; Lebreux, S.J.; Cregger, M.A.; Klingeman, D.M.; Hui, D.; Hettich, R.L.; Wilhelm, S.W.; et al. One-Time Nitrogen Fertilization Shifts Switchgrass Soil Microbiomes within a Context of Larger Spatial and Temporal Variation. PLoS ONE 2019, 14, e0211310. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, A.; Staley, C.; Lamb, J.; Sheaffer, C.; Kaiser, T.; Gutknecht, J.; Sadowsky, M.J.; Rosen, C. Cultivar and Phosphorus Effects on Switchgrass Yield and Rhizosphere Microbial Diversity. Appl. Microbiol. Biotechnol. 2019, 103, 1973–1987. [Google Scholar] [CrossRef]
- Singer, E.; Bonnette, J.; Woyke, T.; Juenger, T.E. Conservation of Endophyte Bacterial Community Structure Across Two Panicum Grass Species. Front. Microbiol. 2019, 10, 2181. [Google Scholar] [CrossRef] [Green Version]
- Bowsher, A.W.; Benucci, G.M.N.; Bonito, G.; Shade, A. Seasonal Dynamics of Core Fungi in the Switchgrass Phyllosphere, and Co-Occurrence with Leaf Bacteria. Phytobiomes J. 2021, 5, 60–68. [Google Scholar] [CrossRef]
- Mohammed, Y.A.; Raun, W.; Kakani, G.; Zhang, H.; Taylor, R.; Desta, K.G.; Jared, C.; Mullock, J.; Bushong, J.; Sutradhar, A.; et al. Nutrient Sources and Harvesting Frequency on Quality Biomass Production of Switchgrass (Panicum virgatum L.) for Biofuel. Biomass Bioenergy 2015, 81, 242–248. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Estaki, M.; Jiang, L.; Bokulich, N.A.; McDonald, D.; González, A.; Kosciolek, T.; Martino, C.; Zhu, Q.; Birmingham, A.; Vázquez-Baeza, Y.; et al. QIIME 2 Enables Comprehensive End-to-End Analysis of Diverse Microbiome Data and Comparative Studies with Publicly Available Data. Curr. Protoc. Bioinform. 2020, 70, e100. [Google Scholar] [CrossRef] [PubMed]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; Version 2.5-7; 2020; Available online: https://rdrr.io/cran/vegan/ (accessed on 21 December 2021).
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.L. An Analysis of Variation Between Upland and Lowland Switchgrass, Panicum virgatum L., in Central Oklahoma. Ecology 1966, 47, 980–992. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorholt, J.A. Microbial Life in the Phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant Compartment and Biogeography Affect Microbiome Composition in Cultivated and Native Agave Species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and Function of the Bacterial Root Microbiota in Wild and Domesticated Barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopold, D.R.; Busby, P.E. Host Genotype and Colonist Arrival Order Jointly Govern Plant Microbiome Composition and Function. Curr. Biol. 2020, 30, 3260–3266.e5. [Google Scholar] [CrossRef]
- Arrigoni, E.; Antonielli, L.; Pindo, M.; Pertot, I.; Perazzolli, M. Tissue Age and Plant Genotype Affect the Microbiota of Apple and Pear Bark. Microbiol. Res. 2018, 211, 57–68. [Google Scholar] [CrossRef]
- Knapp, D.G.; Lázár, A.; Molnár, A.; Vajna, B.; Karácsony, Z.; Váczy, K.Z.; Kovács, G.M. Above-ground Parts of White Grapevine Vitis Vinifera Cv. Furmint Share Core Members of the Fungal Microbiome. Environ. Microbiol. Rep. 2021, 13, 509–520. [Google Scholar] [CrossRef]
- Alderman, S.C.; Halse, R.R.; White, J.F. A Reevaluation of the Host Range and Geographical Distribution of Clavicepser Species in the United States. Plant Dis. 2004, 88, 63–81. [Google Scholar] [CrossRef] [Green Version]
- Lombard, L.; Houbraken, J.; Decock, C.; Samson, R.A.; Meijer, M.; Réblová, M.; Groenewald, J.Z.; Crous, P.W. Generic Hyper-Diversity in Stachybotriaceae. Pers. Int. Mycol. J. 2016, 36, 156–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, O.; Kang, B.; Lee, Y.; Kim, S.; Kwon, J.-H.; Lee, J.; Kim, J. Bacterial Disease Complex Including Bleached Spot, Soft Rot, and Blight on Onion Seedlings Caused by Complex Infections. Plant Dis. 2021, PDIS-03-21-0484-RE. [Google Scholar] [CrossRef] [PubMed]
- Mortuza, M.F.; Tomooka, N.; Habibi, S.; Akatsu, T.; Djedidi, S.; Naito, K.; Ohkama-Ohtsu, N.; Yokoyama, T. Multiphase Characterization of Wild Vigna Associated Root Nodule Bacteria from Japanese Subtropical Islands Unveiled Novel High Temperature Resistant Bradyrhizobium Strains Having High Symbiotic Compatibility with Soybean and Mungbean. Soil Sci. Plant Nutr. 2020, 66, 285–298. [Google Scholar] [CrossRef]
- Lee, M.R.; Hawkes, C.V. Plant and Soil Drivers of Whole-Plant Microbiomes: Variation in Switchgrass Fungi from Coastal to Mountain Sites. Phytobiomes J. 2021, 5, 69–79. [Google Scholar] [CrossRef]
- Kleczewski, N.M.; Bauer, J.T.; Bever, J.D.; Clay, K.; Reynolds, H.L. A Survey of Endophytic Fungi of Switchgrass (Panicum virgatum) in the Midwest, and Their Putative Roles in Plant Growth. Fungal Ecol. 2012, 5, 521–529. [Google Scholar] [CrossRef]
- Yao, H.; Sun, X.; He, C.; Maitra, P.; Li, X.-C.; Guo, L.-D. Phyllosphere Epiphytic and Endophytic Fungal Community and Network Structures Differ in a Tropical Mangrove Ecosystem. Microbiome 2019, 7, 57. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Duan, T.; Sun, X.; Zheng, Y.; Wang, Y.; Hu, M.; Yao, H.; Ji, N.; Lv, P.; Chen, L.; et al. Host Genotype Strongly Influences Phyllosphere Fungal Communities Associated with Mussaenda pubescens Var. alba (Rubiaceae). Fungal Ecol. 2018, 36, 141–151. [Google Scholar] [CrossRef]
- Abdelfattah, A.; Li Destri Nicosia, M.G.; Cacciola, S.O.; Droby, S.; Schena, L. Metabarcoding Analysis of Fungal Diversity in the Phyllosphere and Carposphere of Olive (Olea europaea). PLoS ONE 2015, 10, e0131069. [Google Scholar] [CrossRef] [Green Version]
- Izuno, A.; Kanzaki, M.; Artchawakom, T.; Wachrinrat, C.; Isagi, Y. Vertical Structure of Phyllosphere Fungal Communities in a Tropical Forest in Thailand Uncovered by High-Throughput Sequencing. PLoS ONE 2016, 11, e0166669. [Google Scholar] [CrossRef]
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A Class-Wide Phylogenetic Assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef]
- Prior, R.; Feige, A.; Begerow, D. Antagonistic Activity of the Phyllosphere Fungal Community. Sydowia Int. J. Mycol. 2017, 183–198. [Google Scholar] [CrossRef]
- Trevathan-Tackett, S.; Allnutt, T.; Sherman, C.; Richardson, M.; Crowley, T.; Macreadie, P. Spatial Variation of Bacterial and Fungal Communities of Estuarine Seagrass Leaf Microbiomes. Aquat. Microb. Ecol. 2020, 84, 59–74. [Google Scholar] [CrossRef]
- Jacobsen, C.S.; Hjelmsø, M.H. Agricultural Soils, Pesticides and Microbial Diversity. Curr. Opin. Biotechnol. 2014, 27, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, S.; Srivastava, P.; Devi, R.S.; Bhadouria, R. Influence of Synthetic Fertilizers and Pesticides on Soil Health and Soil Microbiology. In Agrochemicals Detection, Treatment and Remediation; Elsevier: Amsterdam, The Netherlands, 2020; pp. 25–54. ISBN 978-0-08-103017-2. [Google Scholar]
- Santhanam, R.; Luu, V.T.; Weinhold, A.; Goldberg, J.; Oh, Y.; Baldwin, I.T. Native Root-Associated Bacteria Rescue a Plant from a Sudden-Wilt Disease That Emerged during Continuous Cropping. Proc. Natl. Acad. Sci. USA 2015, 112, E5013–E5020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolla, G.; Badri, D.V.; Bakker, M.G.; Manter, D.K.; Vivanco, J.M. Soil Microbiomes Vary in Their Ability to Confer Drought Tolerance to Arabidopsis. Appl. Soil Ecol. 2013, 68, 1–9. [Google Scholar] [CrossRef]
- Singer, E.; Vogel, J.P.; Northen, T.; Mungall, C.J.; Juenger, T.E. Novel and Emerging Capabilities That Can Provide a Holistic Understanding of the Plant Root Microbiome. Phytobiomes J. 2021, 5, 122–132. [Google Scholar] [CrossRef]
- Guretzky, J.A.; Biermacher, J.T.; Cook, B.J.; Kering, M.K.; Mosali, J. Switchgrass for Forage and Bioenergy: Harvest and Nitrogen Rate Effects on Biomass Yields and Nutrient Composition. Plant Soil 2011, 339, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Yeoh, Y.K.; Dennis, P.G.; Paungfoo-Lonhienne, C.; Weber, L.; Brackin, R.; Ragan, M.A.; Schmidt, S.; Hugenholtz, P. Evolutionary Conservation of a Core Root Microbiome across Plant Phyla along a Tropical Soil Chronosequence. Nat. Commun. 2017, 8, 215. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singer, E.; Carpenter, E.M.; Bonnette, J.; Woyke, T.; Juenger, T.E. Impact of Harvest on Switchgrass Leaf Microbial Communities. Genes 2022, 13, 22. https://doi.org/10.3390/genes13010022
Singer E, Carpenter EM, Bonnette J, Woyke T, Juenger TE. Impact of Harvest on Switchgrass Leaf Microbial Communities. Genes. 2022; 13(1):22. https://doi.org/10.3390/genes13010022
Chicago/Turabian StyleSinger, Esther, Elizabeth M. Carpenter, Jason Bonnette, Tanja Woyke, and Thomas E. Juenger. 2022. "Impact of Harvest on Switchgrass Leaf Microbial Communities" Genes 13, no. 1: 22. https://doi.org/10.3390/genes13010022