
Genetic Distribution of Five Spinocerebellar Ataxia Microsatellite Loci in Mexican Native American Populations and Its Impact on Contemporary Mestizo Populations

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
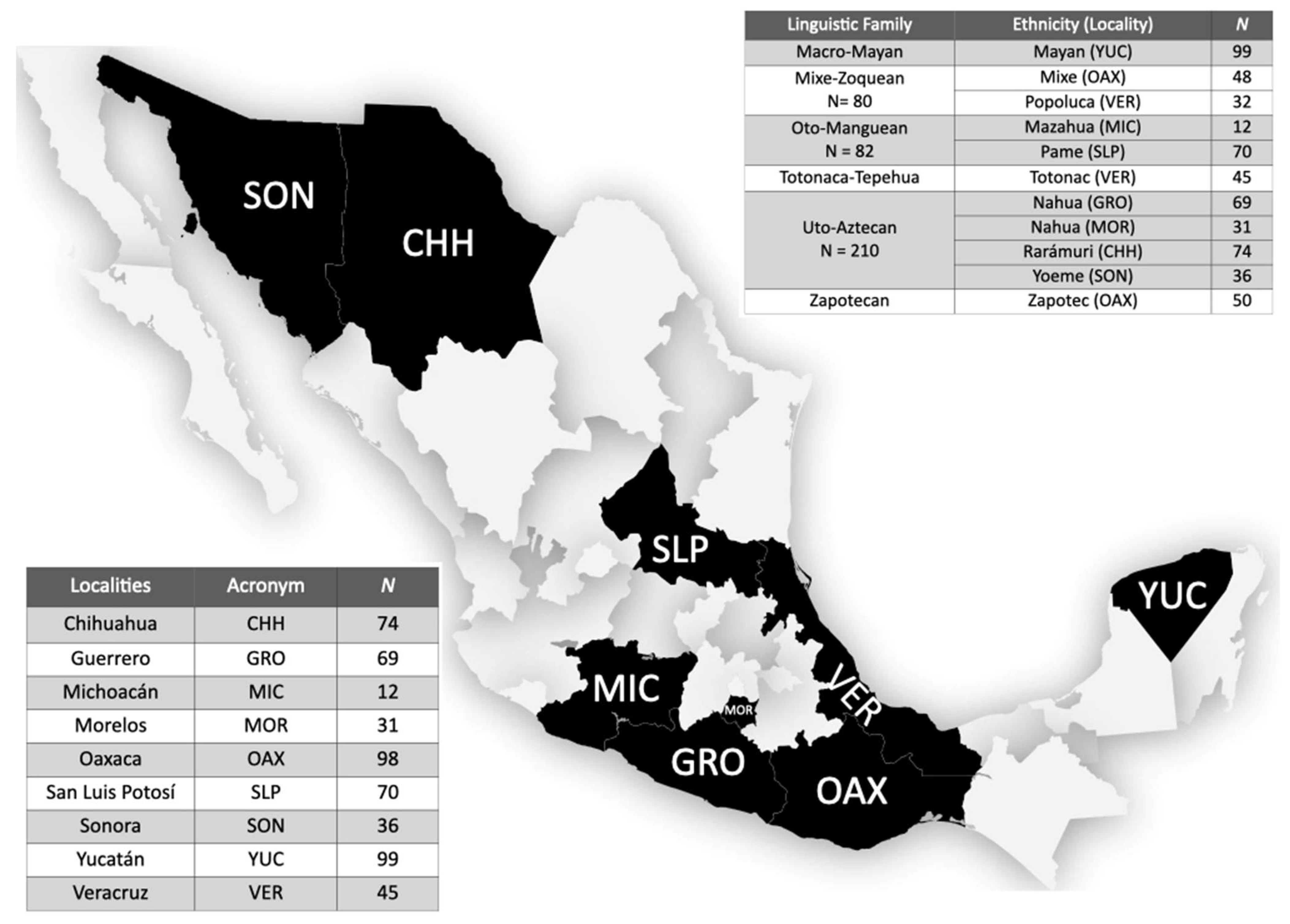
2.1. Subjects
2.2. Molecular Genetic Analysis
2.3. Statistical Analyses
2.4. Comparative Data
3. Results
3.1. Allele Frequencies and Genetic Diversity of Five SCA Loci in Mexican Native American Populations
3.2. Absence of SCA Pathological Alleles in Mexican Native Americans Populations
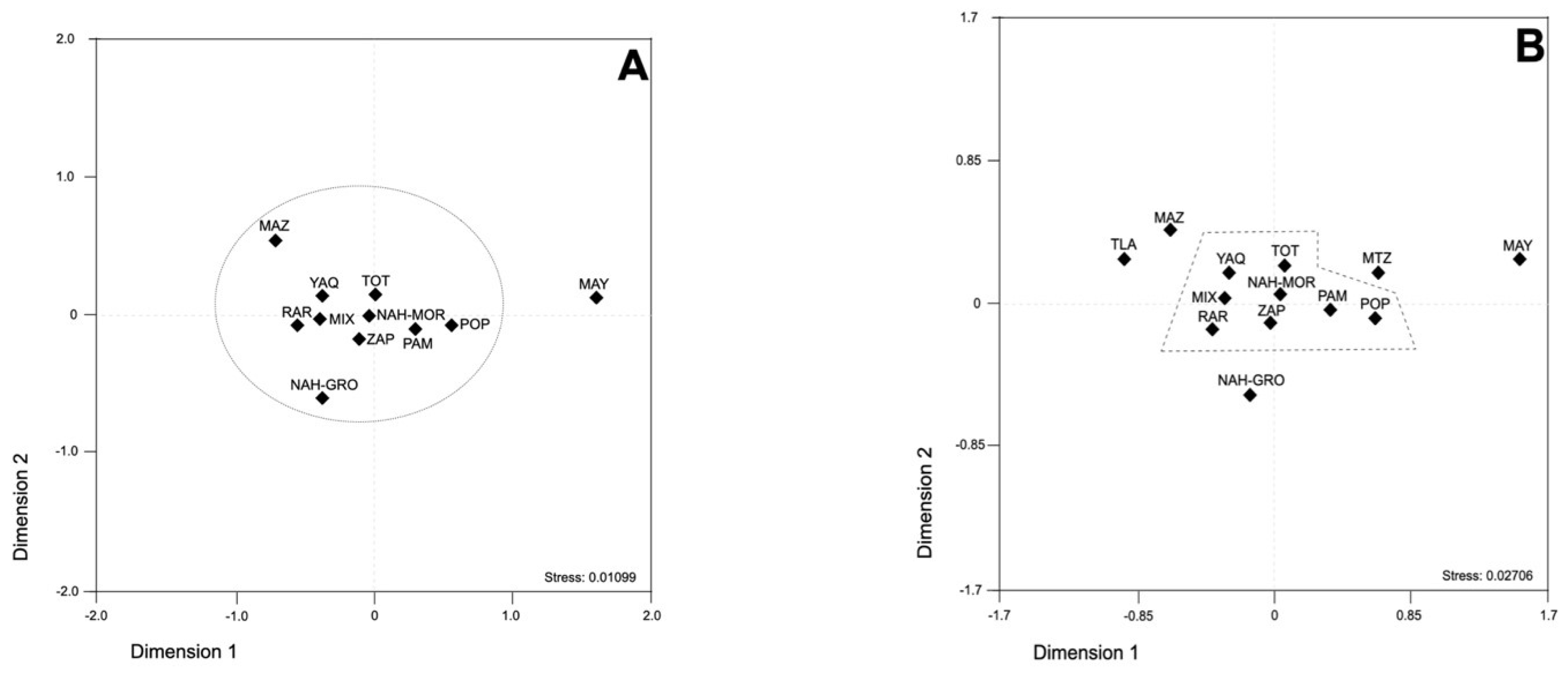
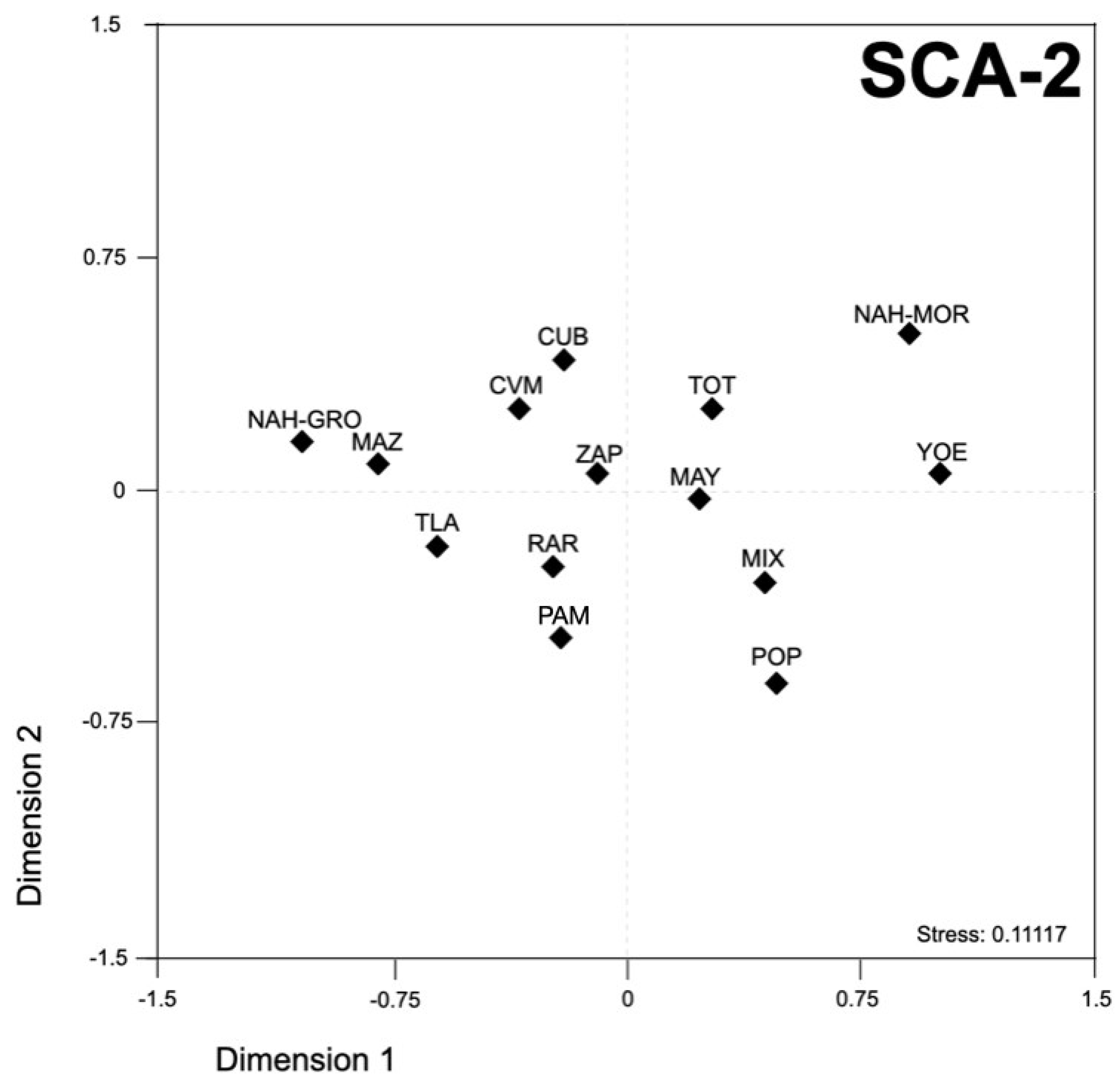
3.3. Genetic Interplay between Mexican Native American Populations
3.4. Comparison of the SCA Loci Allelic Distribution between Mexican Native American and Worldwide Populations
4. Discussion
4.1. Searching for Large SCA Alleles in Mexican Native Americans
4.2. Genetic Landscape of Mexican Native American Populations Using SCA Microsatellite Loci
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Benarroch, L.; Bonne, G.; Rivier, F.; Hamroun, D. The 2021 version of the gene table of neuromuscular disorders (nuclear genome). Neuromuscul. Disord. 2020, 30, 1008–1048. [Google Scholar] [CrossRef]
- Buijsen, R.A.M.; Toonen, L.J.A.; Gardiner, S.L.; van Roon-Mom, W.M.C. Genetics, Mechanisms, and Therapeutic Progress in Polyglutamine Spinocerebellar Ataxias. Neurother. J. Am. Soc. Exp. Neurother. 2019, 16, 263–286. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, S.L.; Boogaard, M.W.; Trompet, S.; de Mutsert, R.; Rosendaal, F.R.; Gussekloo, J.; Jukema, J.W.; Roos, R.A.C.; Aziz, N.A. Prevalence of Carriers of Intermediate and Pathological Polyglutamine Disease-Associated Alleles Among Large Population-Based Cohorts. JAMA Neurol. 2019, 76, 650–656. [Google Scholar] [CrossRef] [PubMed]
- Borbolla-Jiménez, F.V.; Del Prado-Audelo, M.L.; Cisneros, B.; Caballero-Florán, I.H.; Leyva-Gómez, G.; Magaña, J.J. New Perspectives of Gene Therapy on Polyglutamine Spinocerebellar Ataxias: From Molecular Targets to Novel Nanovectors. Pharmaceutics 2021, 13, 1018. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, T.; Mariotti, C.; Paulson, H.L. Spinocerebellar ataxia. Nat. Rev. Dis. Prim. 2019, 5, 24. [Google Scholar] [CrossRef]
- de Oliveira Scott, S.S.; Pedroso, J.L.; Barsottini, O.G.P.; França-Junior, M.C.; Braga-Neto, P. Natural history and epidemiology of the spinocerebellar ataxias: Insights from the first description to nowadays. J. Neurol. Sci. 2020, 417, 117082. [Google Scholar] [CrossRef]
- Rodríguez-Labrada, R.; Martins, A.C.; Magaña, J.J.; Vazquez-Mojena, Y.; Medrano-Montero, J.; Fernandez-Ruíz, J.; Cisneros, B.; Teive, H.; McFarland, K.N.; Saraiva-Pereira, M.L.; et al. Founder Effects of Spinocerebellar Ataxias in the American Continents and the Caribbean. Cerebellum 2020, 19, 446–458. [Google Scholar] [CrossRef]
- Magaña, J.J.; Gómez, R.; Maldonado-Rodríguez, M.; Velázquez-Pérez, L.; Tapia-Guerrero, Y.S.; Cortés, H.; Leyva-García, N.; Hernández-Hernández, O.; Cisneros, B. Origin of the spinocerebellar ataxia type 7 gene mutation in Mexican population. Cerebellum 2013, 12, 902–905. [Google Scholar] [CrossRef]
- Magaña, J.J.; Tapia-Guerrero, Y.S.; Velázquez-Pérez, L.; Cerecedo-Zapata, C.M.; Maldonado-Rodríguez, M.; Jano-Ito, J.S.; Leyva-García, N.; González-Piña, R.; Martínez-Cruz, E.; Hernández-Hernández, O.; et al. Analysis of CAG repeats in five SCA loci in Mexican population: Epidemiological evidence of a SCA7 founder effect. Clin. Genet. 2014, 85, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Sirugo, G.; Williams, S.M.; Tishkoff, S.A. The Missing Diversity in Human Genetic Studies. Cell 2019, 177, 26–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popejoy, A.B.; Fullerton, S.M. Genomics is failing on diversity. Nature 2016, 538, 161–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernández-López, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuña-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. Human genetics. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar] [CrossRef] [Green Version]
- Romero-Hidalgo, S.; Ochoa-Leyva, A.; Garcíarrubio, A.; Acuña-Alonzo, V.; Antúnez-Argüelles, E.; Balcazar-Quintero, M.; Barquera-Lozano, R.; Carnevale, A.; Cornejo-Granados, F.; Fernández-López, J.C.; et al. Demographic history and biologically relevant genetic variation of Native Mexicans inferred from whole-genome sequencing. Nat. Commun. 2017, 8, 1005. [Google Scholar] [CrossRef]
- Dorschner, M.O.; Barden, D.; Stephens, K. Diagnosis of five spinocerebellar ataxia disorders by multiplex amplification and capillary electrophoresis. J. Mol. Diagn. 2002, 4, 108–113. [Google Scholar] [CrossRef] [Green Version]
- Cagnoli, C.; Stevanin, G.; Michielotto, C.; Gerbino Promis, G.; Brussino, A.; Pappi, P.; Durr, A.; Dragone, E.; Viemont, M.; Gellera, C.; et al. Large pathogenic expansions in the SCA2 and SCA7 genes can be detected by fluorescent repeat-primed polymerase chain reaction assay. J. Mol. Diagn. 2006, 8, 128–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Nie, N.; Hull, C.; Bent, D.J.C.S.C. SPSS, Version 20. IBM statistical package for the social sciences. Computer Software: Chicago, IL, USA, 2011.
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing. 2019. Available online: http://www.r-project.org (accessed on 29 August 2021).
- Belkhir, K.; Chikhi, L.; Raufaste, N.B.F. Logiciel Sous Windows TM Pour la Génétique des Populations, GENETIX 4.05; Université Montpellier II: Montpellier, France, 2004.
- García-Ortiz, H.; Barajas-Olmos, F.; Contreras-Cubas, C.; Cid-Soto, M.Á.; Córdova, E.J.; Centeno-Cruz, F.; Mendoza-Caamal, E.; Cicerón-Arellano, I.; Flores-Huacuja, M.; Baca, P.; et al. The genomic landscape of Mexican Indigenous populations brings insights into the peopling of the Americas. Nat. Commun. 2021, 12, 5942. [Google Scholar] [CrossRef] [PubMed]
- Gómez, R.; Vilar, M.G.; Meraz-Ríos, M.A.; Véliz, D.; Zúñiga, G.; Hernández-Tobías, E.A.; Figueroa-Corona, M.D.P.; Owings, A.C.; Gaieski, J.B.; Schurr, T.G. Y chromosome diversity in Aztlan descendants and its implications for the history of Central Mexico. iScience 2021, 24, 102487. [Google Scholar] [CrossRef] [PubMed]
- Takano, H.; Cancel, G.; Ikeuchi, T.; Lorenzetti, D.; Mawad, R.; Stevanin, G.; Didierjean, O.; Dürr, A.; Oyake, M.; Shimohata, T.; et al. Close associations between prevalences of dominantly inherited spinocerebellar ataxias with CAG-repeat expansions and frequencies of large normal CAG alleles in Japanese and Caucasian populations. Am. J. Hum. Genet. 1998, 63, 1060–1066. [Google Scholar] [CrossRef] [Green Version]
- Laffita-Mesa, J.M.; Almaguer-Mederos, L.E.; Kourí, V.; Bauer, P.O.; Vázquez-Mojena, Y.; Cruz Mariño, T.; Velázquez-Pérez, L. Large normal alleles and SCA2 prevalence: Lessons from a nationwide study and analysis of the literature. Clin. Genet. 2014, 86, 96–98. [Google Scholar] [CrossRef]
- Ongaro, L.; Scliar, M.O.; Flores, R.; Raveane, A.; Marnetto, D.; Sarno, S.; Gnecchi-Ruscone, G.A.; Alarcón-Riquelme, M.E.; Patin, E.; Wangkumhang, P.; et al. The Genomic Impact of European Colonization of the Americas. Curr. Biol. 2019, 29, 3974–3986.e4. [Google Scholar] [CrossRef] [PubMed]
- González-Sobrino, B.Z.; Pintado-Cortina, A.P.; Sebastián-Medina, L.; Morales-Mandujano, F.; Contreras, A.V.; Aguilar, Y.E.; Chávez-Benavides, J.; Carrillo-Rodríguez, A.; Silva-Zolezzi, I.; Medrano-González, L. Genetic Diversity and Differentiation in Urban and Indigenous Populations of Mexico: Patterns of Mitochondrial DNA and Y-Chromosome Lineages. Biodemography Soc. Biol. 2016, 62, 53–72. [Google Scholar] [CrossRef]
- Santana, C.; Noris, G.; Meraz-Ríos, M.A.; Magaña, J.J.; Calderon-Aranda, E.S.; Muñoz, M.d.L.; Gómez, R. Genetic analysis of 17 Y-STRs in a Mestizo population from the Central Valley of Mexico. Hum. Biol. 2014, 86, 289–312. [Google Scholar] [CrossRef]
- Saiz, M.; Alvarez-Cubero, M.J.; Lorente, J.A.; Alvarez, J.C.; Martinez-Gonzalez, L.J. Genetic structure in the paternal lineages of South East Spain revealed by the analysis of 17 Y-STRs. Sci. Rep. 2019, 9, 5234. [Google Scholar] [CrossRef]
- Martínez-González, L.J.; Martínez-Espín, E.; Álvarez, J.C.; Albardaner, F.; Rickards, O.; Martínez-Labarga, C.; Calafell, F.; Lorente, J.A. Surname and Y chromosome in Southern Europe: A case study with Colom/Colombo. Eur. J. Hum. Genet. 2012, 20, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Márquez, E.A. Zapotecos del Istmo de Tehuantepec; Comisión Nacional para el Desarrollo de los Pueblos Indigenas: Ciudad de México, Mexico, 2007; ISBN 978-970-753-093-5. [Google Scholar]
- Encyclopaedia Britannica Inc. The Spanish Conquest. Available online: http://www.britannica.com (accessed on 7 September 2021).
- Martins, S.; Sequeiros, J. Origins and Spread of Machado-Joseph Disease Ancestral Mutations Events. Adv. Exp. Med. Biol. 2018, 1049, 243–254. [Google Scholar] [CrossRef]
- Andrés, A.M.; Lao, O.; Soldevila, M.; Calafell, F.; Bertranpetit, J. Dynamics of CAG repeat loci revealed by the analysis of their variability. Hum. Mutat. 2003, 21, 61–70. [Google Scholar] [CrossRef]
- Moctezuma-Zamarron, J.L. Yaquis. In Pueblos Indigenas del Mexico Contemporaneo; Comisión Nacional para el Desarrollo de los Pueblos Indigenas: Ciudad de México, México, 2007. [Google Scholar]
- Cahua-Pablo, J.Á.; Cruz, M.; Tello-Almaguer, P.V.; Del Alarcón-Romero, L.C.; Parra, E.J.; Villerías-Salinas, S.; Valladares-Salgado, A.; Tello-Flores, V.A.; Méndez-Palacios, A.; Pérez-Macedonio, C.P.; et al. Analysis of admixture proportions in seven geographical regions of the state of Guerrero, Mexico. Am. J. Hum. Biol. Off. J. Hum. Biol. Counc. 2017, 29, e23032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, C.; Gutiérrez, G.; Parra, E.J.; Kline, C.; Shriver, M.D. Admixture analysis of a rural population of the state of Guerrero, Mexico. Am. J. Phys. Anthropol. 2005, 128, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Silva-Zolezzi, I.; Hidalgo-Miranda, A.; Estrada-Gil, J.; Fernandez-Lopez, J.C.; Uribe-Figueroa, L.; Contreras, A.; Balam-Ortiz, E.; del Bosque-Plata, L.; Velazquez-Fernandez, D.; Lara, C.; et al. Analysis of genomic diversity in Mexican Mestizo populations to develop genomic medicine in Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 8611–8616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandoval, K.; Moreno-Estrada, A.; Mendizabal, I.; Underhill, P.A.; Lopez-Valenzuela, M.; Peñaloza-Espinosa, R.; Lopez-Lopez, M.; Buentello-Malo, L.; Avelino, H.; Calafell, F.; et al. Y-chromosome diversity in Native Mexicans reveals continental transition of genetic structure in the Americas. Am. J. Phys. Anthropol. 2012, 148, 395–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, D.; Patterson, N.; Campbell, D.; Tandon, A.; Mazieres, S.; Ray, N.; Parra, M.V.; Rojas, W.; Duque, C.; Mesa, N.; et al. Reconstructing Native American population history. Nature 2012, 488, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Perez-Benedico, D.; La Salvia, J.; Zeng, Z.; Herrera, G.A.; Garcia-Bertrand, R.; Herrera, R.J. Mayans: A Y chromosome perspective. Eur. J. Hum. Genet. 2016, 24, 1352–1358. [Google Scholar] [CrossRef] [Green Version]
- Jorge, V.O. Piratas en tierra adentro. Estrategia defensiva de una ciudad novohispana. Siglos XVI al XVIII. In Millars Espai i Història; Universitat Jaume I: Castelló de la Plana, España, 2003; Volume 26. [Google Scholar] [CrossRef]
- Martínez-Cortés, G.; Nuño-Arana, I.; Rubi-Castellanos, R.; Vilchis-Dorantes, G.; Luna-Vázquez, A.; Coral-Vázquez, R.M.; Canto-Cetina, T.; Salazar-Flores, J.; Muñoz-Valle, J.F.; Sandoval-Mendoza, K.; et al. Origin and genetic differentiation of three Native Mexican groups (Purépechas, Triquis and Mayas): Contribution of CODIS-STRs to the history of human populations of Mesoamerica. Ann. Hum. Biol. 2010, 37, 801–819. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Villalobos, H.; Muñoz-Valle, J.F.; González-Martín, A.; Gorostiza, A.; Magaña, M.T.; Páez-Riberos, L.A. Genetic admixture, relatedness, and structure patterns among Mexican populations revealed by the Y-chromosome. Am. J. Phys. Anthropol. 2008, 135, 448–461. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Variation (%) | p-Value |
|---|---|---|
| Global | ||
| Among populations | 3.29 | 0.004 |
| Among individuals within population | 52.43 | ≤0.0001 |
| Within individuals | 44.27 | ≤0.0001 |
| Geography | ||
| Among populations | 4.02 | 0.0001 |
| Among individuals within population | 52.04 | ≤0.0001 |
| Within individuals | 43.94 | ≤0.0001 |
| Language | ||
| Among populations | 3.99 | ≤0.0001 |
| Among individuals within population | 51.98 | ≤0.0001 |
| Within individuals | 44.03 | ≤0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, R.; Tapia-Guerrero, Y.S.; Cisneros, B.; Orozco, L.; Cerecedo-Zapata, C.; Mendoza-Caamal, E.; Leyva-Gómez, G.; Leyva-García, N.; Velázquez-Pérez, L.; Magaña, J.J. Genetic Distribution of Five Spinocerebellar Ataxia Microsatellite Loci in Mexican Native American Populations and Its Impact on Contemporary Mestizo Populations. Genes 2022, 13, 157. https://doi.org/10.3390/genes13010157
Gómez R, Tapia-Guerrero YS, Cisneros B, Orozco L, Cerecedo-Zapata C, Mendoza-Caamal E, Leyva-Gómez G, Leyva-García N, Velázquez-Pérez L, Magaña JJ. Genetic Distribution of Five Spinocerebellar Ataxia Microsatellite Loci in Mexican Native American Populations and Its Impact on Contemporary Mestizo Populations. Genes. 2022; 13(1):157. https://doi.org/10.3390/genes13010157
Chicago/Turabian StyleGómez, Rocío, Yessica S. Tapia-Guerrero, Bulmaro Cisneros, Lorena Orozco, César Cerecedo-Zapata, Elvia Mendoza-Caamal, Gerardo Leyva-Gómez, Norberto Leyva-García, Luis Velázquez-Pérez, and Jonathan J. Magaña. 2022. "Genetic Distribution of Five Spinocerebellar Ataxia Microsatellite Loci in Mexican Native American Populations and Its Impact on Contemporary Mestizo Populations" Genes 13, no. 1: 157. https://doi.org/10.3390/genes13010157