
LncRNAs and the Angiogenic Switch in Cancer: Clinical Significance and Therapeutic Opportunities


Abstract
:1. Introduction
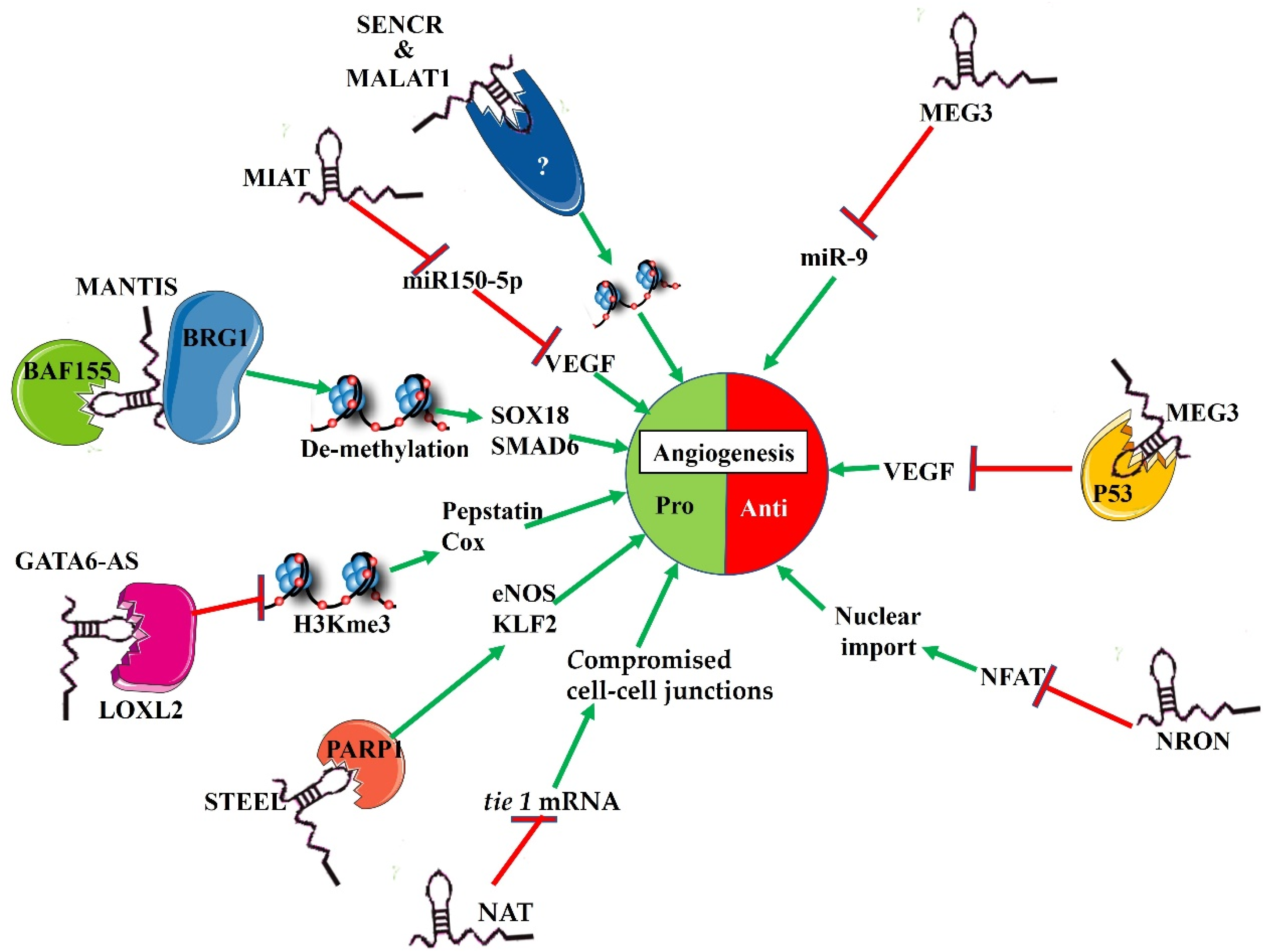
2. Expression Patterns of lncRNAs in Endothelial Cells
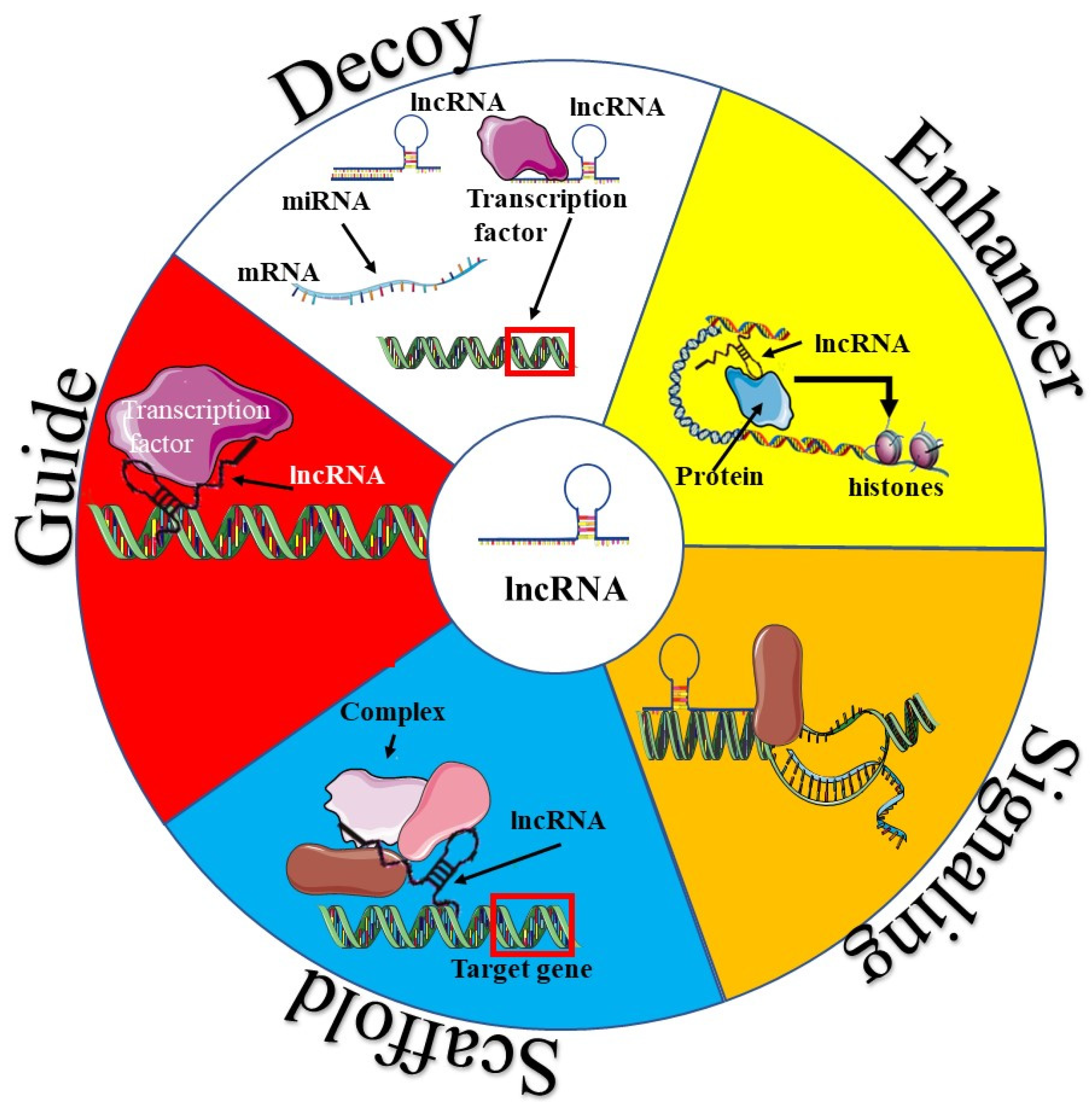
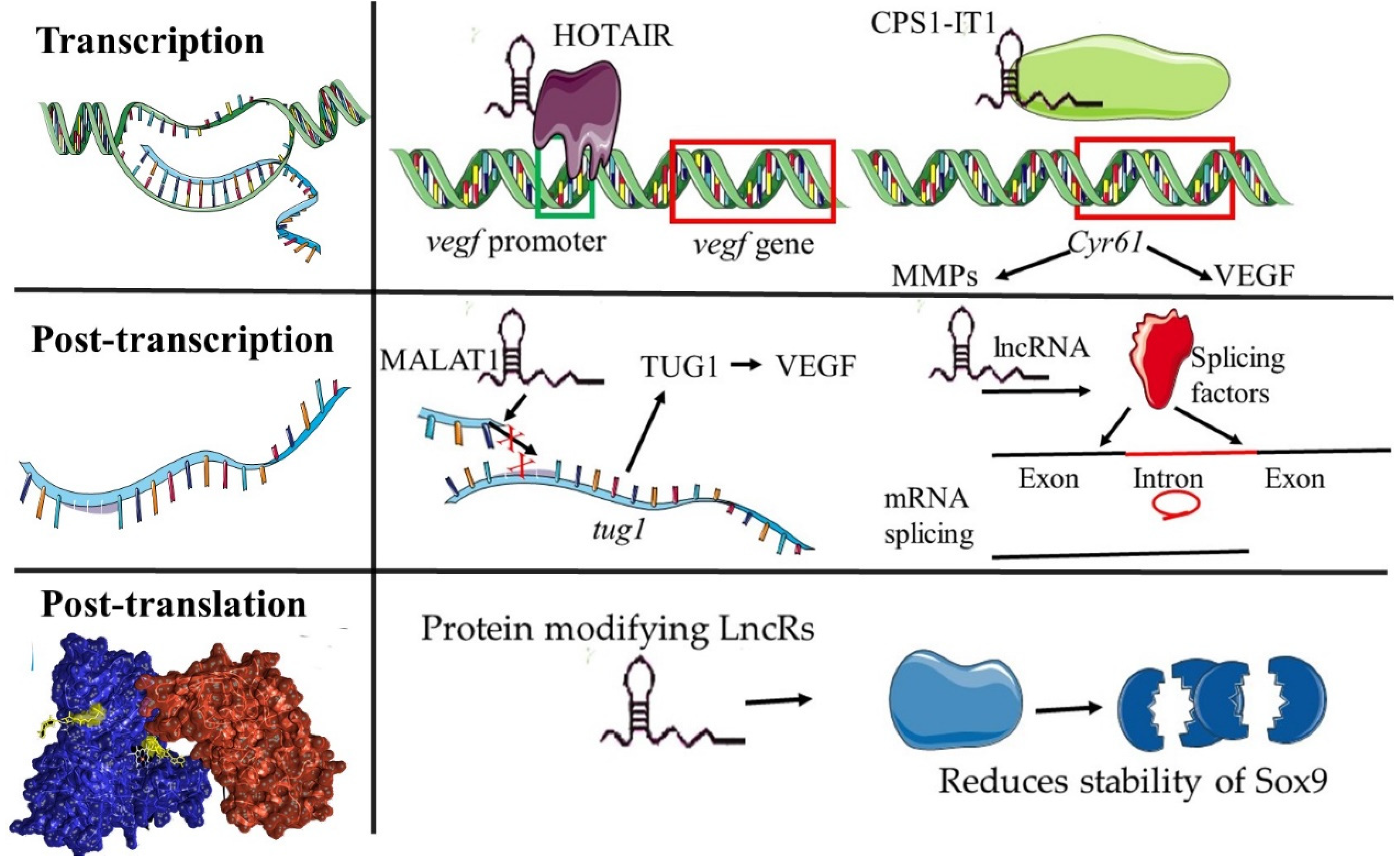
3. Functional Mechanisms of lncRNAs in Angiogenesis
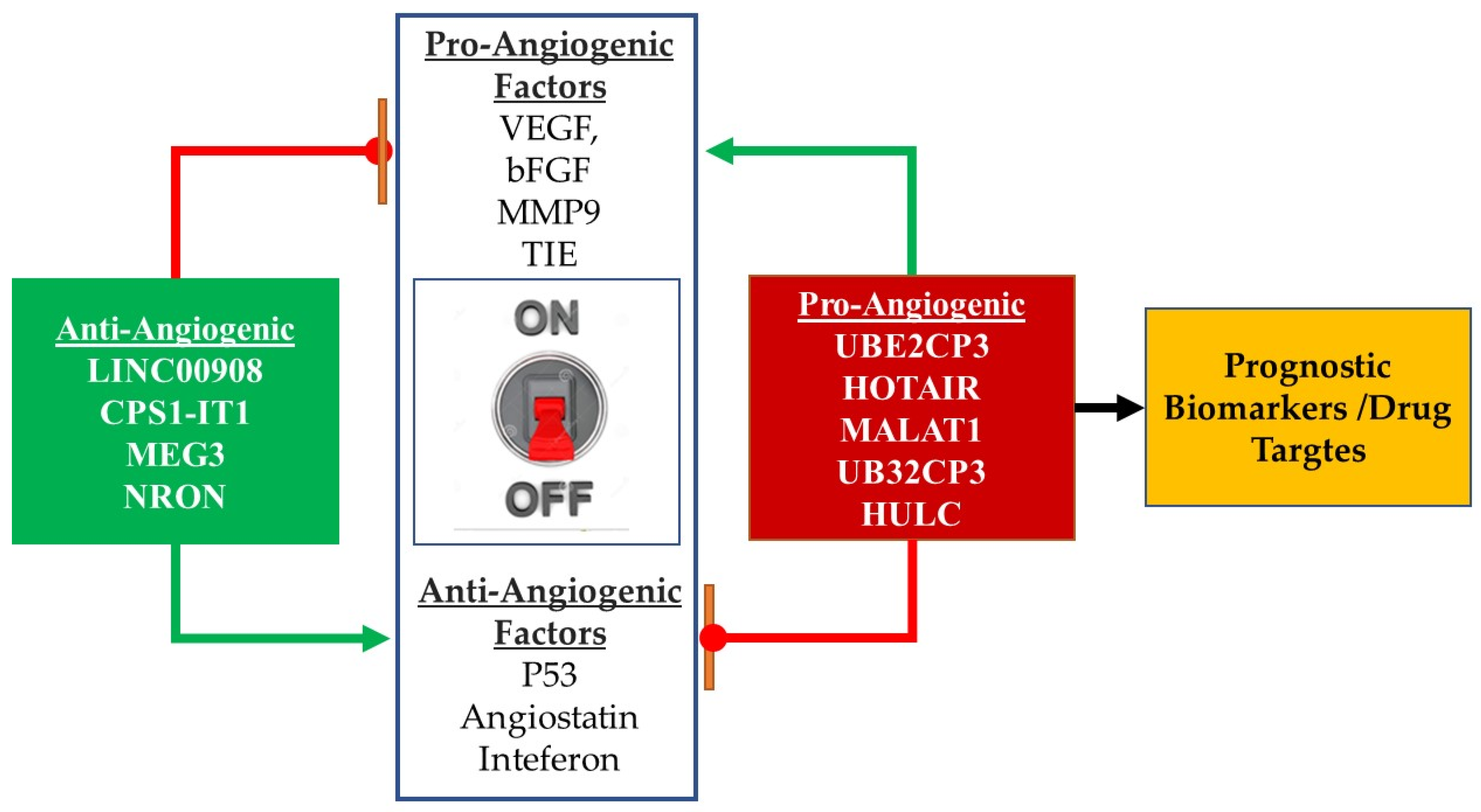
4. Clinical Significance of Angiogenesis Regulating lncRNAs in Cancer
4.1. Angiogenesis Regulating lncRNAs: Role as Cancer Biomarkers
4.2. Therapeutic Targeting of lncRNAs in Cancer Angiogenesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mabeta, P. Paradigms of vascularization in melanoma: Clinical significance and potential for therapeutic targeting. Biomed. Pharmacother. 2020, 127, 110135. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Mabeta, P.; Pepper, M.S. Manipulating the tumor microenvironment: Opportunities for therapeutic targeting. Front. Anti Cancer Drug Discov. 2017, 8, 46–71. [Google Scholar]
- Boulberdaa, M.; Scott, E.; Ballantyne, M.; Garcia, R.; Descamps, B.; Angelini, G.D.; Brittan, M.; Hunter, A.; McBride, M.; McClure, J.; et al. A role for the long noncoding RNA SENCR in commitment and function of endothelial cells. Mol. Ther. 2016, 24, 978–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, P.; Cai, H.; Liu, X.; Chen, J.; Ma, J.; Wang, P.; Liu, Y.; Zheng, J.; Xue, Y. Long non-coding RNA H19 regulates glioma angi-ogenesis and the biological behavior of glioma-associated endothelial cells by inhibiting microRNA-29a. Cancer Lett. 2016, 381, 359–369. [Google Scholar] [CrossRef]
- Josipovic, I.; Pflüger, B.; Fork, C.; Vasconez, A.E.; Oo, J.A.; Hitzel, J.; Seredinski, S.; Gamen, E.; zu Heringdorf, D.M.; Chen, W.; et al. Long noncoding RNA LISPR1 is required for S1P signaling and endothelial cell function. J. Mol. Cell. Cardiol. 2018, 116, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, J.; Tang, N. Long noncoding RNA MALAT1 is a potent autophagy inducer protecting brain microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury by sponging miR-26b and upregulating ULK2 ex-pression. Neuroscience 2017, 354, 1–10. [Google Scholar] [CrossRef]
- Song, Y.; Yang, L.; Guo, R.; Lu, N.; Shi, Y.; Wang, X. Long noncoding RNA MALAT1 promotes high glucose-induced human endothelial cells pyroptosis by affecting NLRP3 expression through competitively binding miR-22. Biochem. Biophys. Res. Commun. 2019, 509, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Puthanveetil, P.; Chen, S.; Feng, B.; Gautam, A.; Chakrabarti, S. Long non-coding RNA MALAT1 regulates hyperglycaemia induced inflammatory process in the endothelial cells. J. Cell. Mol. Med. 2015, 19, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Chen, W.; Hu, X.; Li, J.; Liu, C. Regulatory roles of long noncoding RNAs implicated in cancer hallmarks. Int. J. Cancer 2020, 146, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Bai, X.; Liu, C.; Hu, Z. Long noncoding RNA XIST participates hypoxia-induced angiogenesis in human brain mi-crovascular endothelial cells through regulating MIR-485/SOX7 axis. Am. J. Transl. Res. 2019, 11, 6487–6497. [Google Scholar] [PubMed]
- Khyzha, N.; Khor, M.; DiStefano, P.V.; Wang, L.; Matic, L.; Hedin, U.; Wilson, M.D.; Maegdefessel, L.; Fish, J.E. Regulation of CCL2 expression in human vascular endothelial cells by a neighboring divergently transcribed long noncoding RNA. Proc. Natl. Acad. Sci. USA 2019, 116, 16410–16419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammad, H.M.; Abdelghany, A.A.; Al Ageeli, E.; Kattan, S.W.; Hassan, R.; Toraih, E.A.; Fawzy, M.S.; Mokhtar, N. Long Non-Coding RNAs Gene Variants as Molecular Markers for Diabetic Retinopathy Risk and Response to Anti-VEGF Therapy. Pharm. Pers. Med. 2021, 14, 997–1014. [Google Scholar] [CrossRef] [PubMed]
- Islam Khan, Z.; Tam, S.Y.; Law, H.K. Autophagy-Modulating Long Non-coding RNAs (LncRNAs) and Their Molecular Events in Cancer. Front. Genet. 2019, 9, 750. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F. Autophagy in vascular endothelial cells. Clin. Exp. Pharmacol. Physiol. 2016, 43, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- Bao, H.; Su, H. Long Noncoding RNAs Act as Novel Biomarkers for Hepatocellular Carcinoma: Progress and Prospects. BioMed Res. Int. 2017, 2017, 6049480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, L.; Tang, Q.; Li, G.; Chen, K. Long non-coding RNAs as biomarkers and therapeutic targets: Recent insights into hepa-tocellular carcinoma. Life Sci. 2017, 191, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Bolha, L.; Ravnik-Glavač, M.; Glavač, D. Long Noncoding RNAs as Biomarkers in Cancer. Dis. Markers 2017, 2017, 7243968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossu, A.M.; Mosca, L.; Zappavigna, S.; Misso, G.; Bocchetti, M.; De Micco, F.; Quagliuolo, L.; Porcelli, M.; Caraglia, M.; Boc-cellino, M. Long non-coding RNAs as important biomarkers in laryngeal cancer and other head and neck tumours. Int. J. Mol. Sci. 2019, 20, 3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Gao, G.; Cao, Y. Long Noncoding RNAs as Novel Biomarkers Have a Promising Future in Cancer Diagnostics. Dis. Markers 2016, 2016, 9085195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, R.; Akhade, V.S.; Pal, D.; Rao, S.M. Long noncoding RNAs in development and cancer: Potential biomarkers and therapeutic targets. Mol. Cell. Ther. 2015, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Li, X.; Zhao, H.; Liu, H. Long non-coding RNAs: Potential new biomarkers for predicting tumor invasion and me-tastasis. Mol. Cancer 2016, 15, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, L.; Han, Z.-Y.; Wang, Z.-X.; Qin, L.-X. Long noncoding RNAs, emerging and versatile regulators of tumor-induced angiogenesis. Am. J. Cancer Res. 2019, 9, 1367–1381. [Google Scholar] [PubMed]
- Garajová, I.; Ferracin, M.; Porcellini, E.; Palloni, A.; Abbati, F.; Biasco, G.; Brandi, G. Non-coding RNAs as predictive bi-omarkers to current treatment in metastatic colorectal cancer. Int. J. Mol. Sci. 2017, 18, 1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra Gupta, S.; Nandan Tripathi, Y. Potential of long non-coding RNAs in cancer patients: From biomarkers to therapeutic targets. Int. J. Cancer 2017, 140, 1955–1967. [Google Scholar] [CrossRef] [PubMed]
- Man, H.S.J.; Sukumar, A.N.; Lam, G.C.; Turgeon, P.J.; Yan, M.S.; Ku, K.H.; Dubinsky, M.K.; Ho, J.J.D.; Wang, J.J.; Das, S.; et al. Angiogenic patterning by STEEL, an endothelial-enriched long noncoding RNA. Proc. Natl. Acad. Sci. USA 2018, 115, 2401–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, B.; Wang, S. Angio-lncrs: Lncrnas that regulate angiogenesis and vascular disease. Theranostics 2018, 8, 3654–3675. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lian, Y.; Zhang, Y.; Huang, S.; Zuo, Q.; Yang, N.; Chen, Y.; Wu, D.; Sun, L. The long non-coding RNA PVT1 represses ANGPTL4 transcription through binding with EZH2 in trophoblast cell. J. Cell. Mol. Med. 2018, 22, 1272–1282. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Bao, L.; Chen, Y.; Wang, C.; Luo, M.; Zhang, B.; Zhou, M.; Wang, J.E.; Fang, Y.V.; Kumar, A.; et al. HIF2-Induced Long Noncoding RNA RAB11B-AS1 Promotes Hypoxia-Mediated Angiogenesis and Breast Cancer Metastasis. Cancer Res. 2020, 80, 964–975. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, C.; Cui, H. Long non-coding RNA SNHG22 facilitates hepatocellular carcinoma tumorigenesis and angiogenesis via DNA methylation of microRNA miR-16-5p. Bioengineered 2021, 12, 7446–7458. [Google Scholar] [CrossRef]
- Kondo, A.; Nonaka, A.; Shimamura, T.; Yamamoto, S.; Yoshida, T.; Kodama, T.; Aburatani, H.; Osawa, T. Long Noncoding RNA JHDM1D-AS1 Promotes Tumor Growth by Regulating Angiogenesis in Response to Nutrient Starvation. Mol. Cell. Biol. 2017, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Tang, X.; Hamblin, M.H.; Yin, K.-J. Long non-coding RNA MALAT1 regulates angiogenesis in hindlimb ischemia. Int. J. Mol. Sci. 2018, 19, 1723. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Cao, S.; Wang, Y.; Hu, Y.; Liu, H.; Li, J.; Wang, Q.; Zheng, L. Long non-coding RNA UBE2CP3 enhances HCC cell secretion of VEGFA and promotes angiogenesis by activating ERK1/2/HIF-1α/VEGFA signalling in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Li, Z.; Ma, K.; Li, X.; Tian, N.; Duan, J.; Xiao, X.; Wang, Y. Long non-coding RNA XIST promotes glioma tumor-igenicity and angiogenesis by acting as a molecular sponge of miR-429. J. Cancer 2017, 8, 4106–4116. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, H.; Maheronnaghsh, M.; Molaei, F.; Mashouri, L.; Aref, A.R.; Momeny, M.; Alahari, S.K. Long noncoding RNAs and exosomal lncRNAs: Classification, and mechanisms in breast cancer metastasis and drug resistance. Oncogene 2020, 39, 953–974. [Google Scholar] [CrossRef] [PubMed]
- Bartonicek, N.; Maag, J.L.; Dinger, M.E. Long noncoding RNAs in cancer: Mechanisms of action and technological advance-ments. Mol. Cancer 2016, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.; Xiao, Y.; Ma, J.; Cao, D.; Zhou, Y.; Wang, H.; Liao, Q.; Wang, W. Long non-coding RNAs in esophageal cancer: Molecular mechanisms, functions, and potential applications. J. Hematol. Oncol. 2018, 11, 118. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Dai, X.; Yang, T.; Zhang, N.; Liu, Z.; Jiang, Y. Low long noncoding RNA Growth Arrest-Specific Transcript 5 expression in the exosomes of lung cancer cells promotes tumor angiogenesis. J. Oncol. 2019, 2019, 2476175. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Sun, W.; Guo, Z.; Zhang, J.; Yu, H.; Liu, B. Mechanisms of lncRNA/microRNA interactions in angiogenesis. Life Sci. 2020, 254, 116900. [Google Scholar] [CrossRef]
- Wang, S.-W.; Liu, Z.; Shi, Z.-S. Non-coding RNA in acute ischemic stroke: Mechanisms, biomarkers and therapeutic targets. Cell Transplant. 2018, 27, 1763–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-C.; Tang, C.; Dong, Y.; Zhang, J.; Yuan, T.; Tao, S.-C.; Li, X.-L. Targeting the long noncoding RNA MALAT1 blocks the pro-angiogenic effects of osteosarcoma and suppresses tumour growth. Int. J. Biol. Sci. 2017, 13, 1398–1408. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zu, Y.; Huang, W.; Chen, H.; Xie, H.; Yang, Y. LncRNA CALML3-AS1 promotes tumorigenesis of bladder cancer via regulating ZBTB2 by suppression of microRNA-4316. Biochem. Biophys. Res. Commun. 2018, 504, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Xu, Y.M.; Qin, S.H.; Chen, G.F.; Lau, A.T. Epigenetic regulation of angiogenesis in lung cancer. J. Cell. Physiol. 2021, 236, 3194–3206. [Google Scholar] [CrossRef] [PubMed]
- Kok, F.O.; Baker, A.H. The function of long non-coding RNAs in vascular biology and disease. Vasc. Pharmacol. 2019, 114, 23–30. [Google Scholar] [CrossRef]
- Lecerf, C.; Le Bourhis, X.; Adriaenssens, E. The long non-coding RNA H19: An active player with multiple facets to sustain the hallmarks of cancer. Cell. Mol. Life Sci. 2019, 76, 4673–4687. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Hu, W.; Chen, H.; Wang, Y.; Li, Q.; Xiao, C.; Fan, L.; Zhong, Z.; Chen, X.; Lv, K.; et al. A Novel Human Long Noncoding RNA SCDAL Promotes Angiogenesis through SNF5-Mediated GDF6 Expression. Adv. Sci. 2021, 8, 2004629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yuan, W.; Song, J.; Wang, S.; Gu, X. LncRNA CPS1-IT1 suppresses EMT and metastasis of colorectal cancer by inhibiting hypoxia-induced autophagy through inactivation of HIF-1α. Biochimie 2018, 144, 21–27. [Google Scholar] [CrossRef]
- Peng, Z.; Wang, J.; Shan, B.; Li, B.; Peng, W.; Dong, Y.; Shi, W.; Zhao, W.; He, D.; Duan, M.; et al. The long noncoding RNA LINC00312 induces lung adenocarcinoma migration and vasculogenic mimicry through directly binding YBX1. Mol. Cancer 2018, 17, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Zheng, Y.; Xiao, X.; Liu, C.; Lin, J.; Zheng, S.; Yang, B.; Ou, Q. A circulating long noncoding RNA panel serves as a diagnostic marker for hepatocellular carcinoma. Dis. Markers 2020, 2020, 5417598. [Google Scholar] [CrossRef] [PubMed]
- Lei, D.; Chen, Y.; Zhou, Y.; Hu, G.; Luo, F. An angiogenesis-related long noncoding RNA signature correlates with prognosis in patients with hepatocellular carcinoma. Biosci. Rep. 2021, 41, bsr20204442. [Google Scholar] [CrossRef]
- Liu, C.-G.; Li, J.; Xu, Y.; Li, W.; Fang, S.-X.; Zhang, Q.; Xin, H.-W.; Ma, Z. Long non-coding RNAs and circular RNAs in tumor angiogenesis: From mechanisms to clinical significance. Mol. Ther. Oncolytics 2021, 22, 336–354. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Qiu, M.; Xu, Y.; Mao, Q.; Wang, J.; Dong, G.; Xia, W.; Yin, R.; Xu, L. Differentially expressed protein-coding genes and long noncoding RNA in early-stage lung cancer. Tumor. Biol. 2015, 36, 9969–9978. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.J.; Yin, H.; Hu, J.J.; Wei, X.Z. Long noncoding RNA LINC00313 modulates papillary thyroid cancer tumorigenesis via sponging miR-4429. Neoplasma 2018, 65, 933–942. [Google Scholar] [CrossRef]
- Zhou, X.; Rao, Y.; Sun, Q.; Liu, Y.; Chen, J.; Bu, W. Long noncoding RNA CPS1-IT1 suppresses melanoma cell metastasis through inhibiting Cyr61 via competitively binding to BRG1. J. Cell Physiol. 2019, 234, 22017–22027. [Google Scholar] [CrossRef]
- Dong, R.; Liu, X.-Q.; Zhang, B.-B.; Liu, B.-H.; Zheng, S.; Dong, K.-R. Long non-coding RNA-CRNDE: A novel regulator of tumor growth and angiogenesis in hepatoblastoma. Oncotarget 2017, 8, 42087–42097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Liu, Y.; Chen, Q.; Yu, G.; Chen, J.; Chen, K.; Yang, N.; Zeng, T.; Yan, S.; Huang, A.; et al. Long-noncoding RNA colorectal neoplasia differentially expressed gene as a potential target to upregulate the expression of IRX5 by miR-136-5P to promote oncogenic properties in hepatocellular carcinoma. Cell Physiol. Biochem. 2018, 50, 2229–2248. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.-M.; Lu, Y.-F.; Hu, B.-G.; Liang, W.-C.; Zhu, X.; Yang, H.-D.; Li, G.; Zhang, J.-F. Long noncoding RNA HOTAIR mediated angiogenesis in nasopharyngeal carcinoma by direct and indirect signaling pathways. Oncotarget 2016, 7, 4712–4723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.D.; Yuan, L.L.; Lin, L.Q. LncRNA HOTAIR contributes to the tumorigenesis of nasopharyngeal carcinoma via up-regulating FASN. Eur Rev Med Pharmacol Sci 2017, 21, 5143–5152. [Google Scholar]
- Qu, X.; Alsager, S.; Zhuo, Y.; Shan, B. HOX transcript antisense RNA (HOTAIR) in cancer. Cancer Lett. 2019, 454, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Toy, H.I.; Okmen, D.; Kontou, P.I.; Georgakilas, A.G.; Pavlopoulou, A. HOTAIR as a Prognostic Predictor for Diverse Human Cancers: A Meta- and Bioinformatics Analysis. Cancers 2019, 11, 778. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xia, Y.; Wang, Y.; Xue, C. Pathogenic roles of long noncoding RNAs in melanoma: Implications in diagnosis and therapies. Genes Dis. 2021. [Google Scholar] [CrossRef]
- Zhao, J.; Du, P.; Cui, P.; Qin, Y.; Wu, J.; Zhou, Z.; Zhang, W.; Qin, L.; Huang, G. LncRNA PVT1 promotes angiogenesis via activating the STAT3/VEGFA axis in gastric cancer. Oncogene 2018, 37, 4094–4109. [Google Scholar] [CrossRef] [PubMed]
- Ilboudo, A.; Chouhan, J.; McNeil, B.K.; Osborne, J.R.; Ogunwobi, O.O. PVT1 exon 9: A potential biomarker of aggressive prostate cancer? Int. J. Environ. Res. Public Health 2016, 13, 12. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Li, D.; Gong, M.; Wang, J.; Huang, X.; Wu, T.; Wang, C. Expression and clinical significance of the long non-coding RNA PVT1 in human gastric cancer. OncoTargets Ther. 2014, 7, 1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Ou, C.; Liu, J.; Chen, C.; Zhou, Q.; Yang, S.; Li, G.; Wang, G.; Song, J.; Li, Z.; et al. YAP1-induced MALAT1 promotes epithelial–mesenchymal transition and angiogenesis by sponging miR-126-5p in colorectal cancer. Oncogene 2019, 38, 2627–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, B.; Yadav, S.R.M.; Awasthee, N.; Gupta, S.; Kunnumakkara, A.B.; Gupta, S.C. Diagnostic, prognostic, and therapeutic significance of long non-coding RNA MALAT1 in cancer. Biochim. Biophys. Acta Bioenerg. 2021, 1875, 188502. [Google Scholar] [CrossRef]
- Yu, X.; Hu, L.; Li, S.; Shen, J.; Wang, D.; Xu, R.; Yang, H. Long non-coding RNA Taurine upregulated gene 1 promotes osteo-sarcoma cell metastasis by mediating HIF-1α via miR-143-5p. Cell Death Dis. 2019, 10, 280. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Liu, X.; Zheng, J.; Xue, Y.; Ma, J.; Li, Z.; Bao, M.; Liu, Y. Long non-coding RNA Taurine upregulated 1 enhances tu-mor-induced angiogenesis through inhibiting microRNA-299 in human glioblastoma. Oncogene 2017, 36, 318–331. [Google Scholar] [CrossRef]
- Yang, C.; Zheng, J.; Liu, X.; Xue, Y.; He, Q.; Dong, Y.; Wang, D.; Li, Z.; Liu, L.; Ma, J.; et al. Role of ANKHD1/LINC00346/ZNF655 feedback loop in regulating the glioma angiogenesis via staufen1-mediated mRNA decay. Mol. Ther. Nucleic Acids 2020, 20, 866–878. [Google Scholar] [CrossRef]
- Zhang, B.; Li, C.; Sun, Z. Long non-coding RNA LINC00346, LINC00578, LINC00673, LINC00671, LINC00261, and SNHG9 are novel prognostic markers for pancreatic cancer. Am. J. Transl. Res. 2018, 10, 2648. [Google Scholar]
- Pichler, M.; Rodriguez-Aguayo, C.; Nam, S.Y.; Dragomir, M.P.; Bayraktar, R.; Anfossi, S.; Knutsen, E.; Ivan, C.; Fuentes-Mattei, E.; Kil Lee, S.; et al. Therapeutic potential of FLANC, a novel primate-specific long non-coding RNA in colorectal cancer. Gut 2020, 69, 1818–1831. [Google Scholar] [CrossRef]
- Teppan, J.; Barth, D.A.; Prinz, F.; Jonas, K.; Pichler, M.; Klec, C. Involvement of long non-coding RNAs (lncRNAs) in tumor angiogenesis. Non-coding RNA 2020, 6, 42. [Google Scholar] [CrossRef]
- Chong, Z.X.; Yeap, S.K.; Ho, W.Y. Angiogenesis regulation by microRNAs and long non-coding RNAs in human breast cancer. Pathol. Res. Pract. 2021, 13, 153326. [Google Scholar] [CrossRef]
- Hu, X.; Li, Q.; Zhang, J. The long noncoding RNA LINC00908 facilitates hepatocellular carcinoma progression via interaction with Sox-4. Cancer Manag. Res. 2019, 11, 8789. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Huang, C.; Gong, Z.; Zhao, Y.; Tang, K.; Li, X.; Fan, S.; Shi, L.; Li, X.; Zhang, P.; et al. Expression of LINC00312, a long intergenic non-coding RNA, is negatively correlated with tumor size but positively correlated with lymph node metastasis in nasopharyngeal carcinoma. J. Mol. Histol. 2013, 44, 545–554. [Google Scholar] [CrossRef]
- Yoshimura, H.; Matsuda, Y.; Yamamoto, M.; Kamiya, S.; Ishiwata, T. Expression and role of long non-coding RNA H19 in carcinogenesis. Front. Biosci. 2018, 23, 614–625. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Esmaeili, M.; Taheri, M. H19 lncRNA: Roles in tumorigenesis. Biomed. Pharmacother. 2020, 123, 109774. [Google Scholar] [CrossRef]
- Ariel, I.; Sughayer, M.; Fellig, Y.; Pizov, G.; Ayesh, S.; Podeh, D.; Libdeh, B.A.; Levy, C.; Birman, C.T.; Tykocinski, M.L.; et al. The imprinted H19 gene is a marker of early recurrence in human bladder carcinoma. Mol. Pathol. 2000, 53, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, Y.; Zhang, X.; Lv, J.; Li, Y.; Liu, X.; Xu, T.P. H19 serves as a diagnostic biomarker and up-regulation of H19 expression contributes to poor prognosis in patients with gastric cancer. Neoplasma 2016, 63, 223–230. [Google Scholar] [CrossRef]
- Peng, W.; Gao, W.; Feng, J. Long noncoding RNA HULC is a novel biomarker of poor prognosis in patients with pancreatic cancer. Med. Oncol. 2014, 31, 346. [Google Scholar] [CrossRef]
- Xian, H.P.; Zhuo, Z.L.; Sun, Y.J.; Liang, B.; Zhao, X.T. Circulating long non-coding RNAs HULC and ZNFX1-AS1 are potential biomarkers in patients with gastric cancer. Oncol. Lett. 2018, 16, 4689–4698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Ota, Y.; Kogure, T.; Suzuki, Y.; Iwamoto, H.; Yamakita, K.; Kitano, Y.; Fujii, S.; Haneda, M.; Patel, T.; et al. Circulating extracellular vesicle-encapsulated HULC is a potential biomarker for human pancreatic cancer. Cancer Sci. 2020, 111, 98–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Chen, Z.; Guo, S.; Liao, X.; Xie, H.; Zheng, Y.; Cai, B.; Huang, P.; Liu, Y.; Zhou, Q.; et al. TUG1, SPRY4-IT1, and HULC as valuable prognostic biomarkers of survival in cancer: A prisma-compliant meta-analysis. Medicine 2017, 96, e8583. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Zheng, Q.; Xiong, J.; Wu, H.; Wang, W.; Zhou, W. Long non-coding RNA MVIH promotes cell proliferation, migration, invasion through regulating multiple cancer-related pathways, and correlates with worse prognosis in pancreatic ductal adenocarcinomas. Am. J. Transl. Res. 2020, 12, 2118–2135. [Google Scholar] [PubMed]
- Zhang, Y.; Lin, S.; Yang, X.; Zhang, X. Prognostic and Clinicopathological significance of lncRNA MVIH in cancer patients. J. Cancer 2019, 10, 1503–1510. [Google Scholar] [CrossRef]
- Wang, Y.; Han, D.; Pan, L.; Sun, J. The positive feedback between lncRNA TNK2-AS1 and STAT3 enhances angiogenesis in non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2018, 507, 185–192. [Google Scholar] [CrossRef]
- Luo, Z.; Pan, J.; Ding, Y.; Zhang, Y.-S.; Zeng, Y. The function and clinical relevance of lncRNA UBE2CP3-001 in human gliomas. Arch. Med Sci. 2018, 14, 1308–1320. [Google Scholar] [CrossRef]
- Umeda, S.; Kanda, M.; Kodera, Y. Recent advances in molecular biomarkers for patients with hepatocellular carcinoma. Expert Rev. Mol. Diagn. 2019, 19, 725–738. [Google Scholar] [CrossRef]
- Yuan, S.X.; Yang, F.; Yang, Y.; Tao, Q.F.; Zhang, J.; Huang, G.; Yang, Y.; Wang, R.-Y.; Yang, S.; Huo, X.-S.; et al. Long noncoding RNA associated with microvascular invasion in hepatocellular carcinoma promotes angiogenesis and serves as a predictor for hepatocellular carcinoma patients’ poor recurrence-free survival after hepatectomy. Hepatology 2012, 56, 2231–2241. [Google Scholar] [CrossRef]
- Nie, F.-Q.; Zhu, Q.; Xu, T.-P.; Zou, Y.-F.; Xie, M.; Sun, M.; Xia, R.; Lu, K.-H. Long non-coding RNA MVIH indicates a poor prognosis for non-small cell lung cancer and promotes cell proliferation and invasion. Tumor Biol. 2014, 35, 7587–7594. [Google Scholar] [CrossRef]
- Lei, B.; Xu, S.-P.; Liang, X.-S.; Li, Y.-W.; Zhang, J.-F.; Zhang, G.-Q.; Pang, D. Long non-coding RNA MVIH is associated with poor prognosis and malignant biological behavior in breast cancer. Tumor Biol. 2016, 37, 5257–5264. [Google Scholar] [CrossRef]
- Kumar, M.M.; Goyal, R. LncRNA as a Therapeutic Target for Angiogenesis. Curr. Top. Med. Chem. 2017, 17, 1750–1757. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; He, X.; Zheng, Z.; Ma, X.; Hu, X.; Wu, D.; Wang, M. Serum HOTAIR as a novel diagnostic biomarker for esophageal squamous cell carcinoma. Mol. Cancer 2017, 16, 75. [Google Scholar] [CrossRef] [Green Version]
- Botti, G.; De Chiara, A.; Di Bonito, M.; Cerrone, M.; Malzone, M.G.; Collina, F.; Cantile, M. Noncoding RNAs within the HOX gene network in tumor pathogenesis and progression. J. Cell. Physiol. 2019, 234, 395–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Zhou, L.; Wu, L.-M.; Lai, M.-C.; Xie, H.-Y.; Zhang, F.; Zheng, S.-S. Overexpression of Long Non-coding RNA HOTAIR Predicts Tumor Recurrence in Hepatocellular Carcinoma Patients Following Liver Transplantation. Ann. Surg. Oncol. 2011, 18, 1243–1250. [Google Scholar] [CrossRef]
- Li, N.; Wang, Y.; Liu, X.; Luo, P.; Jing, W.; Zhu, M.; Jiancheng, T. Identification of Circulating Long Noncoding RNA HOTAIR as a Novel Biomarker for Diagnosis and Monitoring of Non–Small Cell Lung Cancer. Technol. Cancer Res. Treat. 2017, 16, 1060–1066. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Cao, Y.; Gong, X.; Li, H. Long noncoding RNAs in head and neck cancer. Oncotarget 2017, 8, 10726–10740. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Chen, X.; Li, J.; Guo, Y.; Li, H.; Pan, X.; Jiang, J.; Liu, H.; Wu, B. Salivary HOTAIR and PVT1 as novel biomarkers for early pancreatic cancer. Oncotarget 2016, 7, 25408–25419. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.; Wu, L.-X.; Yang, Y.; Li, S.; Mei, Y.; Liu, Y.-B.; Zhang, L.; Cheng, Y.; Zhou, H.-H. lncRNAs PVT1 and HAR1A are prognosis biomarkers and indicate therapy outcome for diffuse glioma patients. Oncotarget 2017, 8, 78767–78780. [Google Scholar] [CrossRef] [Green Version]
- Yim, G.W.; Kim, H.J.; Kim, L.K.; Kim, S.W.; Kim, S.; Nam, E.J.; Kim, Y.T. Long Non-coding RNA HOXA11 Antisense Promotes Cell Proliferation and Invasion and Predicts Patient Prognosis in Serous Ovarian Cancer. Cancer Res. Treat. 2017, 49, 656–668. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long Noncoding RNA and Cancer: A New Paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.-L.; Lu, R.-Y.; Wang, L.-K.; Wang, Y.-Y.; Dai, Y.-J.; Wang, C.-Y.; Yang, Y.-J.; Guo, F.; Xue, J.; Yang, D.-D. Long noncoding RNA HOTAIR silencing inhibits invasion and proliferation of human colon cancer LoVo cells via regulating IGF2BP2. J. Cell. Biochem. 2019, 120, 1221–1231. [Google Scholar] [CrossRef]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long Noncoding RNA MALAT1 Regulates Endothelial Cell Function and Vessel Growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef]
- Han, P.; Li, J.-W.; Zhang, B.-M.; Lv, J.-C.; Li, Y.-M.; Gu, X.-Y.; Yu, Z.-W.; Jia, Y.-H.; Bai, X.-F.; Li, L.; et al. The lncRNA CRNDE promotes colorectal cancer cell proliferation and chemoresistance via miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol. Cancer 2017, 16, 9. [Google Scholar] [CrossRef] [Green Version]
- Spencer, H.L.; Sanders, R.; Boulberdaa, M.; Meloni, M.; Cochrane, A.; Spiroski, A.-M.; Mountford, J.; Emanueli, C.; Caporali, A.; Brittan, M.; et al. The LINC00961 transcript and its encoded micropeptide, small regulatory polypeptide of amino acid response, regulate endothelial cell function. Cardiovasc. Res. 2020, 116, 1981–1994. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LncR | Cancer | Expression | Mechanism of Action | Potential Application | Reference |
|---|---|---|---|---|---|
| LINC00313 | Lung, thyroid | Upregulated | Inhibits the transcription of genes regulating cell motility | Prognosis | [52,53] |
| CPS1-IT1 | Multiple | Upregulated | Inhibits VEGF, MMP-9 and Cyr61 | Prognosis | [54,55] |
| CRNDE | hepatoblastoma, leukemia | Upregulated | Modulates the PI3K/PKB/mTOR pathway | Prognosis, Identification of subtype (in Leukemia) | [56] |
| HOTAIR | Nasopharyngeal carcinoma | Upregulated | targets the VEGF promoter and activates the transcription of VEGF; modulates Ang2 expression through the upregulation of GRP78 | prognosis, recurrence | [57,58,59,60] |
| HOTAIR | Melanoma | downregulated | prognosis | [59,61] | |
| PVT1 | gastric cancer | Upregulated | activates VEGF via STAT3 | aggressiveness | [62,63,64] |
| MALAT1 | Multiple | Upregulated | promotes the expression of VEGF, SLUG and Twist | detection, risk of metastasis, prognosis | [65,66] |
| TUG1 | Multiple | Upregulated | modulates HIF-1α expression, promotes VEGF expression | prognosis | [67,68] |
| LINC00346 | Glioma | Upregulated | induces ZNF655 degradation | prognosis | [69,70] |
| FLANC | CRC | Upregulated | induces VEGF expression via STAT3 | prognosis | [71,72] |
| LINC00908 | TNBC HCC | downregulated | inhibits STAT3 phosphorylation, decreases VEGF expression | prognosis | [73,74] |
| LINC00312 | lung cancer Nasopharyngeal carcinoma | Upregulated | induces VEGF expression | prognosis | [48,75] |
| H19 | bladder cancer, gastric cancer | Upregulated | increases VEGF expression | early recurrence, prognosis | [76,77,78,79] |
| HULC | HCC | Upregulated | promotes SPHKI expression | metastasis | [80,81,82,83] |
| MVIH | HCC | Upregulated | interacts with PGK1 | prognosis | [84,85] |
| TNK2-AS1 | NSCLC | Upregulated | prognosis | [86] | |
| UBE2CP3 | glioma, HCC | Upregulated | activates the ERK1/2/HIF-1α/VEGF pathway | prognosis | [49,87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mabeta, P.; Hull, R.; Dlamini, Z. LncRNAs and the Angiogenic Switch in Cancer: Clinical Significance and Therapeutic Opportunities. Genes 2022, 13, 152. https://doi.org/10.3390/genes13010152
Mabeta P, Hull R, Dlamini Z. LncRNAs and the Angiogenic Switch in Cancer: Clinical Significance and Therapeutic Opportunities. Genes. 2022; 13(1):152. https://doi.org/10.3390/genes13010152
Chicago/Turabian StyleMabeta, Peace, Rodney Hull, and Zodwa Dlamini. 2022. "LncRNAs and the Angiogenic Switch in Cancer: Clinical Significance and Therapeutic Opportunities" Genes 13, no. 1: 152. https://doi.org/10.3390/genes13010152