
Novel Pathogenic Variants in PJVK, the Gene Encoding Pejvakin, in Subjects with Autosomal Recessive Non-Syndromic Hearing Impairment and Auditory Neuropathy Spectrum Disorder

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Clinical Tests
2.3. DNA Purification, Genotyping and Sequencing
2.4. Assessment of Pathogenicity of DNA Variants
3. Results
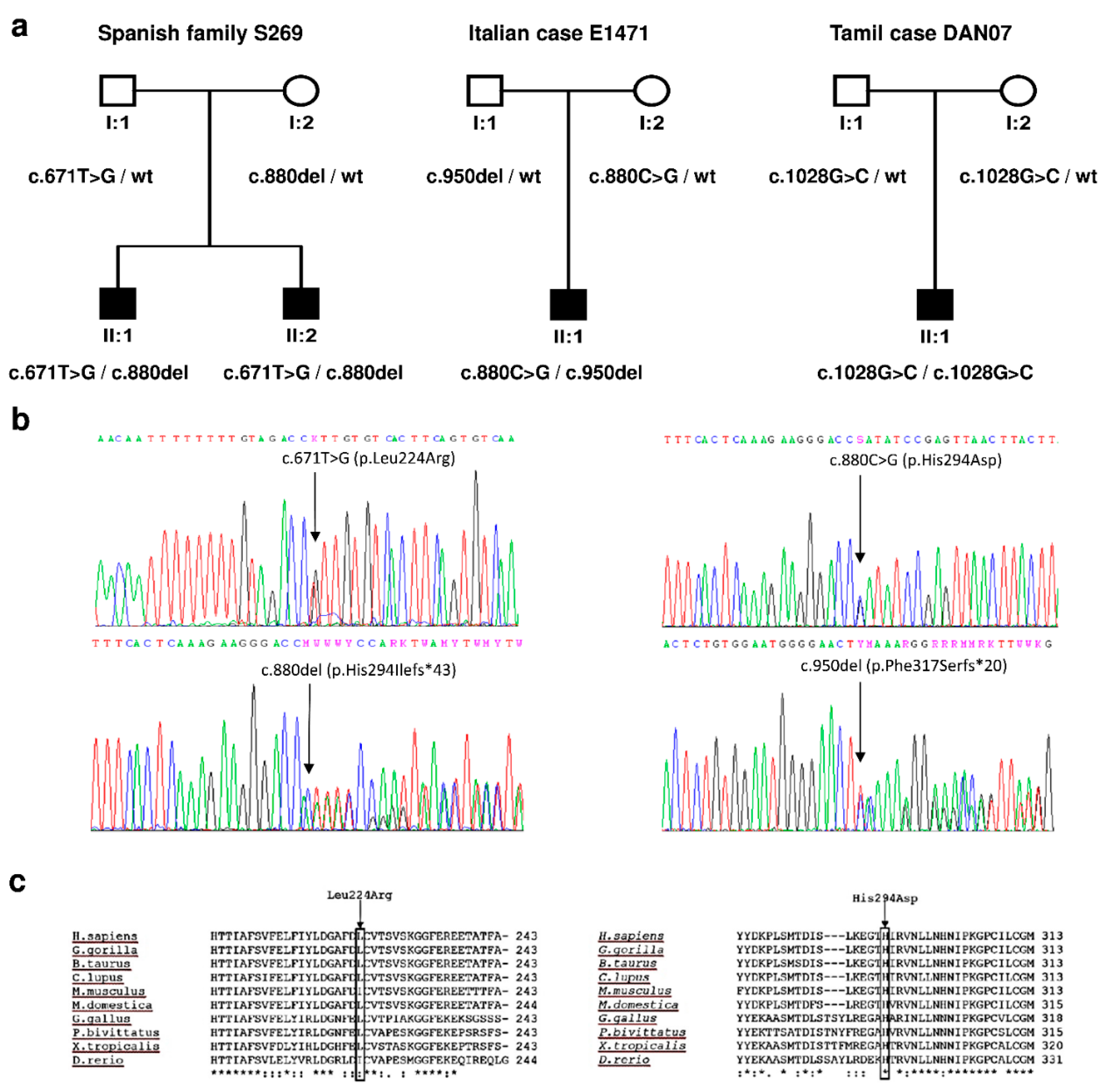
3.1. Genetic Study
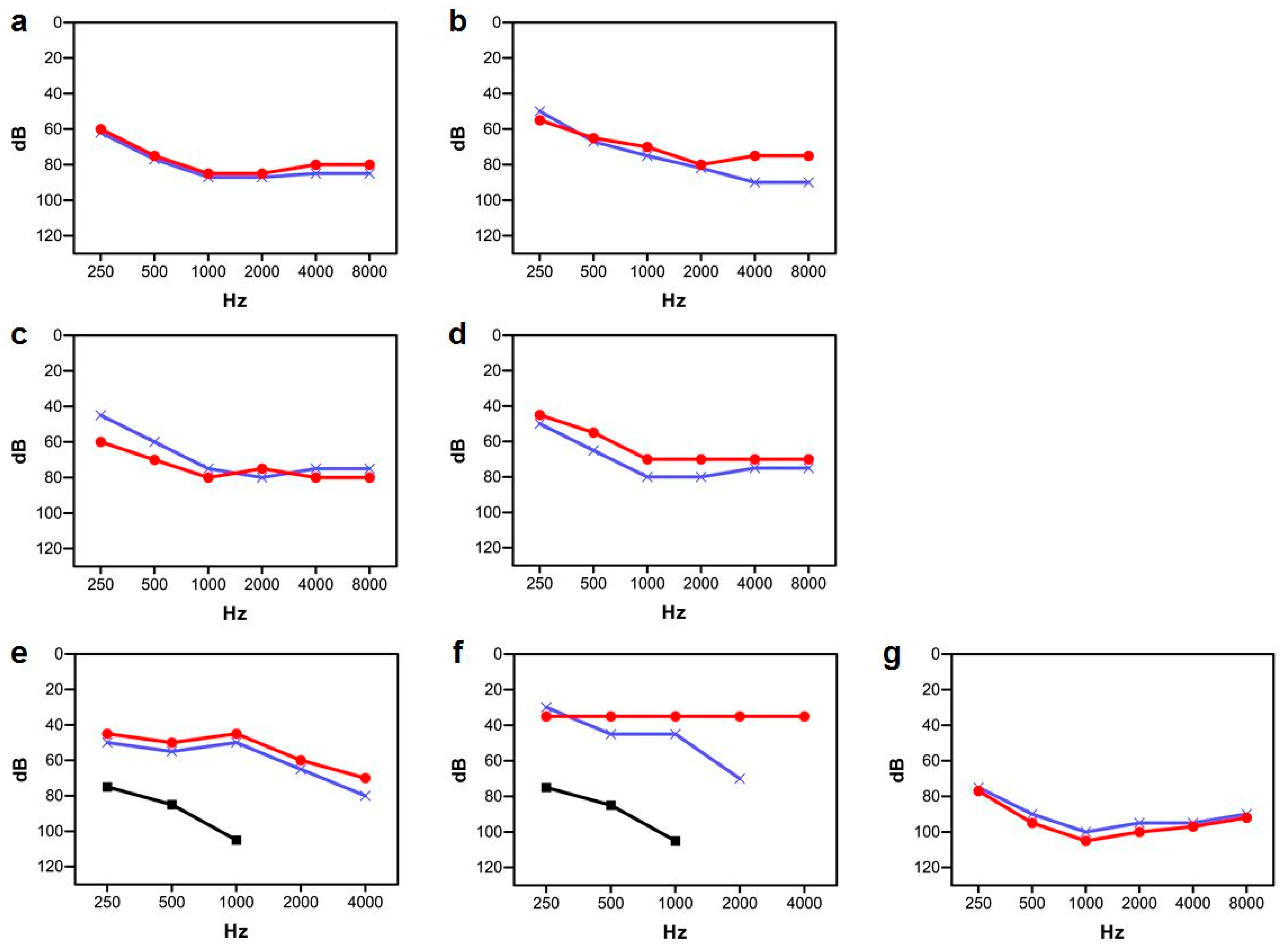
3.2. Clinical Study
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dror, A.A.; Avraham, K.B. Hearing impairment: A panoply of genes and functions. Neuron 2010, 68, 293–308. [Google Scholar] [CrossRef] [Green Version]
- Van Camp, G.; Smith, R.J. Hereditary Hearing Loss Homepage. Available online: https://hereditaryhearingloss.org (accessed on 15 December 2021).
- Hoefsloot, L.H.; Feenstra, I.; Kunst, H.P.; Kremer, H. Genotype phenotype correlations for hearing impairment: Approaches to management. Clin. Genet. 2014, 85, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Delmaghani, S.; del Castillo, F.J.; Michel, V.; Leibovici, M.; Aghaie, A.; Ron, U.; Van Laer, L.; Ben-Tal, N.; Van Camp, G.; Weil, D.; et al. Mutations in the gene encoding pejvakin, a newly identified protein of the afferent auditory pathway, cause DFNB59 auditory neuropathy. Nat. Genet. 2006, 38, 770–778. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, E.; Roelandt, R.; Riquet, F.B.; Van Camp, G.; Wullaert, A.; Vandenabeele, P. Punching Holes in Cellular Membranes: Biology and Evolution of Gasdermins. Trends Cell Biol. 2021, 31, 500–513. [Google Scholar] [CrossRef]
- Rogers, C.; Alnemri, E.S. Gasdermins in apoptosis: New players in an old game. Yale J. Biol. Med. 2019, 92, 603–617. [Google Scholar] [PubMed]
- Kazmierczak, M.; Kazmierczak, P.; Peng, A.W.; Harris, S.L.; Shah, P.; Puel, J.L.; Lenoir, M.; Franco, S.J.; Schwander, M. Pejvakin, a Candidate Stereociliary Rootlet Protein, Regulates Hair Cell Function in a Cell-Autonomous Manner. J. Neurosci. 2017, 37, 3447–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmaghani, S.; Defourny, J.; Aghaie, A.; Beurg, M.; Dulon, D.; Thelen, N.; Perfettini, I.; Zelles, T.; Aller, M.; Meyer, A.; et al. Hypervulnerability to Sound Exposure through Impaired Adaptive Proliferation of Peroxisomes. Cell 2015, 163, 894–906. [Google Scholar] [CrossRef] [Green Version]
- Defourny, J.; Aghaie, A.; Perfettini, I.; Avan, P.; Delmaghani, S.; Petit, C. Pejvakin-mediated pexophagy protects auditory hair cells against noise-induced damage. Proc. Natl. Acad. Sci. USA 2019, 116, 8010–8017. [Google Scholar] [CrossRef] [Green Version]
- Chaleshtori, M.H.; Simpson, M.A.; Farrokhi, E.; Dolati, M.; Hoghooghi-Rad, L.; Amani Geshnigani, S.; Crosby, A.H. Novel mutations in the pejvakin gene are associated with autosomal recessive non-syndromic hearing loss in Iranian families. Clin. Genet. 2007, 72, 261–263. [Google Scholar] [CrossRef]
- Ebermann, I.; Walger, M.; Scholl, H.P.; Charbel Issa, P.; Lüke, C.; Nürnberg, G.; Lang-Roth, R.; Becker, C.; Nürnberg, P.; Bolz, H.J. Truncating mutation of the DFNB59 gene causes cochlear hearing impairment and central vestibular dysfunction. Hum. Mutat. 2007, 28, 571–577. [Google Scholar] [CrossRef]
- Collin, R.W.; Kalay, E.; Oostrik, J.; Caylan, R.; Wollnik, B.; Arslan, S.; den Hollander, A.I.; Birinci, Y.; Lichtner, P.; Strom, T.M.; et al. Involvement of DFNB59 mutations in autosomal recessive nonsyndromic hearing impairment. Hum. Mutat. 2007, 28, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Schwander, M.; Sczaniecka, A.; Grillet, N.; Bailey, J.S.; Avenarius, M.; Najmabadi, H.; Steffy, B.M.; Federe, G.C.; Lagler, E.A.; Banan, R.; et al. A forward genetics screen in mice identifies recessive deafness traits and reveals that pejvakin is essential for outer hair cell function. J. Neurosci. 2007, 27, 2163–2175. [Google Scholar] [CrossRef] [Green Version]
- Shahin, H.; Walsh, T.; Rayyan, A.A.; Lee, M.K.; Higgins, J.; Dickel, D.; Lewis, K.; Thompson, J.; Baker, C.; Nord, A.S.; et al. Five novel loci for inherited hearing loss mapped by SNP-based homozygosity profiles in Palestinian families. Eur. J. Hum. Genet. 2010, 18, 407–413. [Google Scholar] [CrossRef] [Green Version]
- Borck, G.; Rainshtein, L.; Hellman-Aharony, S.; Volk, A.E.; Friedrich, K.; Taub, E.; Magal, N.; Kanaan, M.; Kubisch, C.; Shohat, M.; et al. High frequency of autosomal-recessive DFNB59 hearing loss in an isolated Arab population in Israel. Clin. Genet. 2012, 82, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Babanejad, M.; Fattahi, Z.; Bazazzadegan, N.; Nishimura, C.; Meyer, N.; Nikzat, N.; Sohrabi, E.; Najmabadi, A.; Jamali, P.; Habibi, F.; et al. A comprehensive study to determine heterogeneity of autosomal recessive nonsyndromic hearing loss in Iran. Am. J. Med. Genet. 2012, 158A, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Mujtaba, G.; Bukhari, I.; Fatima, A.; Naz, S. A p.C343S missense mutation in PJVK causes progressive hearing loss. Gene 2012, 504, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Sloan-Heggen, C.M.; Babanejad, M.; Beheshtian, M.; Simpson, A.C.; Booth, K.T.; Ardalani, F.; Frees, K.L.; Mohseni, M.; Mozafari, R.; Mehrjoo, Z.; et al. Characterising the spectrum of autosomal recessive hereditary hearing loss in Iran. J. Med. Genet. 2015, 52, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.J.; Lan, L.; Li, N.; Qi, Y.; Zong, L.; Shi, W.; Yu, L.; Wang, H.; Yang, J.; Xie, L.Y.; et al. Identification of a novel mutation of PJVK in the Chinese non-syndromic hearing loss population with low prevalence of the PJVK mutations. Acta Otolaryngol. 2015, 135, 211–216. [Google Scholar] [CrossRef]
- Bademci, G.; Foster, J.; Mahdieh, N.; Bonyadi, M.; Duman, D.; Cengiz, F.B.; Menendez, I.; Diaz-Horta, O.; Shirkavand, A.; Zeinali, S.; et al. Comprehensive analysis via exome sequencing uncovers genetic etiology in autosomal recessive nonsyndromic deafness in a large multiethnic cohort. Genet. Med. 2016, 18, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Tekin, D.; Bademci, G.; Foster, J.; Cengiz, F.B.; Kannan-Sundhari, A.; Guo, S.; Mittal, R.; Zou, B.; Grati, M.; et al. Spectrum of DNA variants for non-syndromic deafness in a large cohort from multiple continents. Hum. Genet. 2016, 135, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Alimardani, M.; Hosseini, S.M.; Khaniani, M.S.; Haghi, M.R.; Eslahi, A.; Farjami, M.; Chezgi, J.; Derakhshan, S.M.; Mojarrad, M. Targeted Mutation Analysis of the SLC26A4, MYO6, PJVK and CDH23 Genes in Iranian Patients with AR Nonsyndromic Hearing Loss. Fetal Pediatr. Pathol. 2019, 38, 93–102. [Google Scholar] [CrossRef]
- Khan, A.; Han, S.; Wang, R.; Ansar, M.; Ahmad, W.; Zhang, X. Sequence variants in genes causing nonsyndromic hearing loss in a Pakistani cohort. Mol. Genet. Genomic Med. 2019, 7, e917. [Google Scholar] [CrossRef] [PubMed]
- Richard, E.M.; Santos-Cortez, R.L.P.; Faridi, R.; Rehman, A.U.; Lee, K.; Shahzad, M.; Acharya, A.; Khan, A.A.; Imtiaz, A.; Chakchouk, I.; et al. Global genetic insight contributed by consanguineous Pakistani families segregating hearing loss. Hum. Mutat. 2019, 40, 53–72. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Tariq, M.; He, S.; Abdullah, U.; Zhang, J.; Baig, S.M. Whole exome sequencing identified mutations causing hearing loss in five consanguineous Pakistani families. BMC Med. Genet. 2020, 21, 151. [Google Scholar] [CrossRef] [PubMed]
- Salime, S.; Charif, M.; Bousfiha, A.; Elrharchi, S.; Bakhchane, A.; Charoute, H.; Kabine, M.; Snoussi, K.; Lenaers, G.; Barakat, A. Homozygous mutations in PJVK and MYO15A genes associated with non-syndromic hearing loss in Moroccan families. Int. J. Pediatr. Otorhinolaryngol. 2017, 101, 25–29. [Google Scholar] [CrossRef]
- Starr, A.; Picton, T.W.; Sininger, Y.; Hood, L.J.; Berlin, C.I. Auditory neuropathy. Brain 1996, 119, 741–753. [Google Scholar] [CrossRef]
- Arslan, E.; Genovese, E.; Orzan, E.; Turrini, M. Valutazione della Percezione Verbale nel Bambino Ipoacusico; Ecumenica: Roma, Italy, 1997; pp. 1–79. [Google Scholar]
- Elliott, L.L.; Katz, D. Development of a New Children’s Test of Speech Discrimination (Technical Manual); Auditec: St. Louis, MO, USA, 1980. [Google Scholar]
- Dib, C.; Fauré, S.; Fizames, C.; Samson, D.; Drouot, N.; Vignal, A.; Millasseau, P.; Marc, S.; Hazan, J.; Seboun, E.; et al. A comprehensive genetic map of the human genome based on 5,264 microsatellites. Nature 1996, 380, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. ACMG Laboratory Quality Assurance Committee. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- VarSome: The Human Genomic Variant Search Engine. Available online: https://varsome.com/ (accessed on 15 December 2021).
- Oza, A.M.; DiStefano, M.T.; Hemphill, S.E.; Cushman, B.J.; Grant, A.R.; Siegert, R.K.; Shen, J.; Chapin, A.; Boczek, N.J.; Schimmenti, L.A.; et al. ClinGen Hearing Loss Clinical Domain Working Group. Expert specification of the ACMG/AMP variant interpretation guidelines for genetic hearing loss. Hum. Mutat. 2018, 39, 1593–1613. [Google Scholar] [CrossRef] [PubMed]
- Genome Aggregation Database (gnomAD). Available online: https://gnomad.broadinstitute.org/ (accessed on 15 December 2021).
- Boothroyd, A. The acoustic speech signal. In Pediatric audiology: Diagnosis, technology, and management; Madell, J.R., Flexer, C., Eds.; Thieme: New York, NY, USA, 2008; pp. 159–167. [Google Scholar]
- Del Castillo, I.; Morín, M.; Domínguez-Ruiz, M.; Moreno-Pelayo, M.A. Genetic Etiology of Non-Syndromic Hearing Loss in Europe. Hum. Genet. 2022; in press. [Google Scholar]



{kind=link}
{kind=link}
| Exon | Primer Sequences (5′-3′) | [MgCl2] |
|---|---|---|
| 1 | F: CTAGGCCGCAGTTCTTTGTCCTTAG R: TCCCAGGCAAACGCCATTACA | 2.5 mM |
| 2 | F: GCAGAGGCAGGGAATTATACAGT R: ACAAACTTTTGGCATTGTTAATCTT | 2.0 mM |
| 3 | F: TGGTGAGTCATGTTGCCTTTCT R: CAACCTCAATGTTTTAAGCATTCTT | 1.5 mM |
| 4 | F: CTGACTATTAGGATTGCCTTGATTT R: CAGCTCTTTCATCAGAACATTTCA | 1.5 mM |
| 5 | F: TTGTTTTTGGTAGGATTATAGGAAA R: GAGAGCACATGCCCTAATGAAT | 2.5 mM |
| 6 | F: TCATCACCCCATCAAACAATAA R: GAATAGAAAACCTCATGTGTTAAGC | 1.5 mM |
| 7 | F: GCTGTTTGCATTATGTATTTTTCA R: TGTGGCACAACTGCACCTAA | 2.0 mM |
| Variant | SIFT Score | Polyphen-2 Score | Minor Allele Frequency (MAF) [31] | ACMG Criteria | Classification | |
|---|---|---|---|---|---|---|
| DNA | Protein | |||||
| c.671T->G | p.Leu224Arg | 0.01 (deleterious) | 0.959 (Probably damaging) | 2 × 10−5 (global) 4 × 10−5 (Non-Finnish Europeans) | PM2 (strong), PM3 (strong), PP1 (supporting) | Likely pathogenic |
| c.880C>G | p.His294Asp | 0.00 (deleterious) | 0.981 (Probably damaging) | 4 × 10−6 (global) 8 × 10−6 (Non-Finnish Europeans) | PM2 (strong), PM3 (moderate) | Likely pathogenic |
| Exon | DNA Level | Protein Level | Reference |
|---|---|---|---|
| 2 | c.113dup | p.Lys41Glufs*8 | [11,26] |
| 2 | c.122del | p.Lys41Serfs*18 | [13,16] |
| 2 | c.147T>A | p.Tyr49* | [23] |
| 2 | c.158C>G | p.Ser53* | [24] |
| 2 | c.161C>T | p.Thr54Ile | [4] |
| 2 | c.162_172del | p.Pro55fs*23 | [24] |
| intron 2 | c.211+1G>T | [18] | |
| 3 | c.274C>T | p.Arg92* | [16,18] |
| 3 | c.406C>T | p.Arg136* | [14,15,24,26] |
| 4 | c.485G>A | p.Ser162Asn | [25] |
| 4 | c.499C>T | p.Arg167* | [12,18,20] |
| 4 | c.547C>T | p.Arg183Trp | [4,12,21,22] |
| 6 | c.671T>G | p.Leu224Arg | This work |
| 6 | c.726del | p.Phe242Leufs*7 | [10] |
| 6 | deletion of whole exon | [18] | |
| 7 | c.880del | p.His294Ilefs*43 | This work |
| 7 | c.880C>G | p.His294Asp | This work |
| 7 | c.908_910del | p.Asn303del | [24] |
| 7 | c.930_931del | p.Cys312Trpfs*19 | [19] |
| 7 | c.950del | p.Phe317Serfs*20 | This work |
| 7 | c.970G>T | p.Gly324Trp | [18] |
| 7 | c.988del | p.Val330Leufs*7 | [10] |
| 7 | c.1028G>C | p.Cys343Ser | [17], This work |
| Genotype | Families | Features of the Hearing Loss | Reference | |||
|---|---|---|---|---|---|---|
| Onset | Severity | Evolution | AN | |||
| p.[Lys41Glufs*8];[Lys41Glufs*8] | 1 (Moroccan) | Prelingual | S-P | Progressive | No | [11] |
| 1 (Moroccan) | Prelingual | P | NR | NT | [26] | |
| p.[Lys41Serfs*18];[Lys41Serfs*18] | 1 (Iranian) | NR | M-P | Progressive | NT | [13] |
| 1 (Iranian) | Prelingual | P | Progressive | NT | [16] | |
| p.[Tyr49*];[Tyr49*] | 1 (Pakistani) | Prelingual | NR | NR | NT | [23] |
| p.[Ser53*];[Ser53*] | 1 (Pakistani) | NR | NR | NR | NT | [24] |
| p.[Thr54Ile];[Thr54Ile] | 1 (Iranian) | Prelingual | S | NR | Yes | [4] |
| p.[Pro55fs*23];[Pro55fs*23] | 1 (Pakistani) | NR | NR | NR | NT | [24] |
| c.[211 + 1G > T];[211 + 1G > T] | 1 (Iranian) | Prelingual | NR | NR | NT | [18] |
| p.[Arg92*];[Arg92*] | 1 (Iranian) | Prelingual | S-P | Stable | NT | [16] |
| 1 (Iranian) | Prelingual | S | NR | NT | [18] | |
| p.[Arg136*];[Arg136*] | 1 (Palestinian) | Prelingual | P | NR | No | [14] |
| 3 (Israeli Arab) | Prelingual | M-S | Stable | No | [15] | |
| 1 (Pakistani) | NR | NR | NR | NT | [24] | |
| 1 (Moroccan) | Prelingual | P | NR | NT | [26] | |
| p.[Ser162Asn];[p.Ser162Asn] | 1 (Pakistani) | Prelingual | P | NR | NT | [25] |
| p.[Arg167*];[Arg167*] | 1 (Turkish) | NR | S-P | NR | No | [12] |
| 1 (Iranian) | Prelingual | P | NR | NT | [18] | |
| 1 (Turkish) | Prelingual | NR | NR | NT | [20] | |
| p.[Arg183Trp];[Arg183Trp] | 3 (Iranian) | Prelingual | P | NR | Yes | [4] |
| 1 (Turkish) | Prelingual | S-P | NR | No | [12] | |
| 1 (Iranian) | Prelingual | NR | NR | NT | [21] | |
| 1 (Iranian) | NR | NR | NR | NT | [22] | |
| p.[Leu224Arg];[His294Ilefs*43] | 1 (Spanish) | Early childhood | S | Stable | No | This work |
| p.[Phe242Leufs*7];[Phe242Leufs*7] | 1 (Iranian) | NR | P | NR | NT | [10] |
| Homozygous deletion of exon 6 | 1 (Iranian) | Prelingual | NR | NR | NT | [18] |
| p.[His294Asp];[Phe317Serfs*20] | 1 (Italian) | Early childhood | P | Stable | Yes | This work |
| p.[Asn303del];[Asn303del] | 1 (Pakistani) | NR | NR | NR | NT | [24] |
| p.[Cys312Trpfs*19];[Cys312Trpfs*19] | 1 (Chinese) | Prelingual | S-P | Progressive | No | [19] |
| p.[Gly324Trp];[Gly324Trp] | 1 (Iranian) | Prelingual | S-P | NR | NT | [18] |
| p.[Val330Leufs*7];[Val330Leufs*7] | 1 (Iranian) | NR | P | NR | No | [10] |
| p.[Cys343Ser];[Cys343Ser] | 1 (Pakistani) | Early childhood | S-P | Progressive | NT | [17] |
| 1 (Tamil) | Prelingual | P | Stable | Yes | This work | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Ruiz, M.; Rodríguez-Ballesteros, M.; Gandía, M.; Gómez-Rosas, E.; Villamar, M.; Scimemi, P.; Mancini, P.; Rendtorff, N.D.; Moreno-Pelayo, M.A.; Tranebjaerg, L.; et al. Novel Pathogenic Variants in PJVK, the Gene Encoding Pejvakin, in Subjects with Autosomal Recessive Non-Syndromic Hearing Impairment and Auditory Neuropathy Spectrum Disorder. Genes 2022, 13, 149. https://doi.org/10.3390/genes13010149
Domínguez-Ruiz M, Rodríguez-Ballesteros M, Gandía M, Gómez-Rosas E, Villamar M, Scimemi P, Mancini P, Rendtorff ND, Moreno-Pelayo MA, Tranebjaerg L, et al. Novel Pathogenic Variants in PJVK, the Gene Encoding Pejvakin, in Subjects with Autosomal Recessive Non-Syndromic Hearing Impairment and Auditory Neuropathy Spectrum Disorder. Genes. 2022; 13(1):149. https://doi.org/10.3390/genes13010149
Chicago/Turabian StyleDomínguez-Ruiz, María, Montserrat Rodríguez-Ballesteros, Marta Gandía, Elena Gómez-Rosas, Manuela Villamar, Pietro Scimemi, Patrizia Mancini, Nanna D. Rendtorff, Miguel A. Moreno-Pelayo, Lisbeth Tranebjaerg, and et al. 2022. "Novel Pathogenic Variants in PJVK, the Gene Encoding Pejvakin, in Subjects with Autosomal Recessive Non-Syndromic Hearing Impairment and Auditory Neuropathy Spectrum Disorder" Genes 13, no. 1: 149. https://doi.org/10.3390/genes13010149