
Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica



Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of WAK/WAKL Members in J. regia and Its Wild Related Species J. mandshurica
2.2. Chromosome Localization and Gene Duplication Mode Analysis
2.3. Physicochemical Properties and Subcellular Localization Prediction
2.4. Phylogenetic Tree Construction and Gene Structure Analysis
2.5. Cis-Acting Element Prediction and Gene Expression Analysis
3. Results
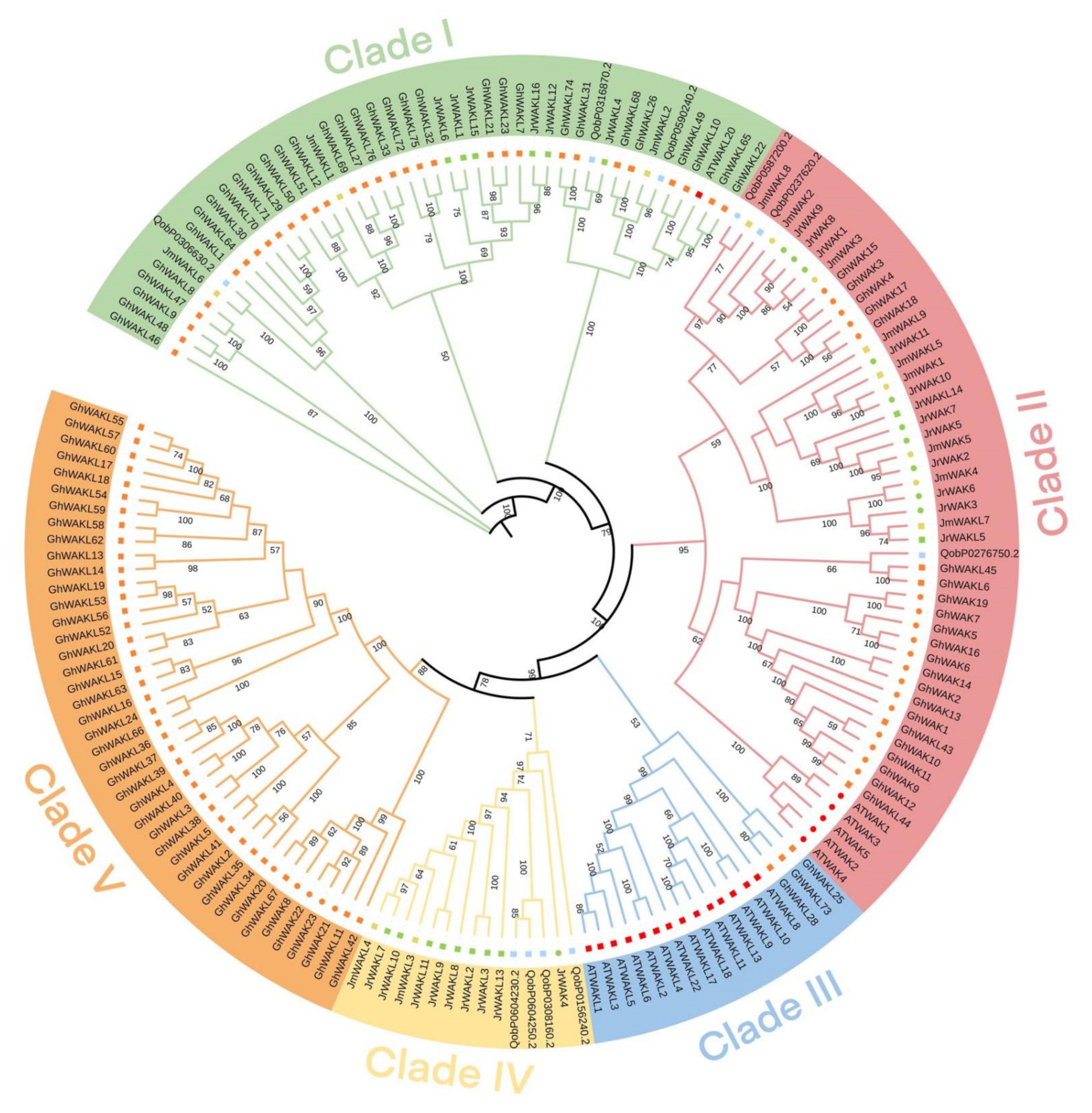
3.1. Genome-Wide Identification and Phylogenetic Analysis of the WAK/WAKL Gene Family in J. regia and Its Wild Related Species J. mandshurica
3.2. Physicochemical Property and Subcellular Localization Analysis of the WAK/WAKL Proteins in J. regia and Its Wild Related Species J. mandshurica
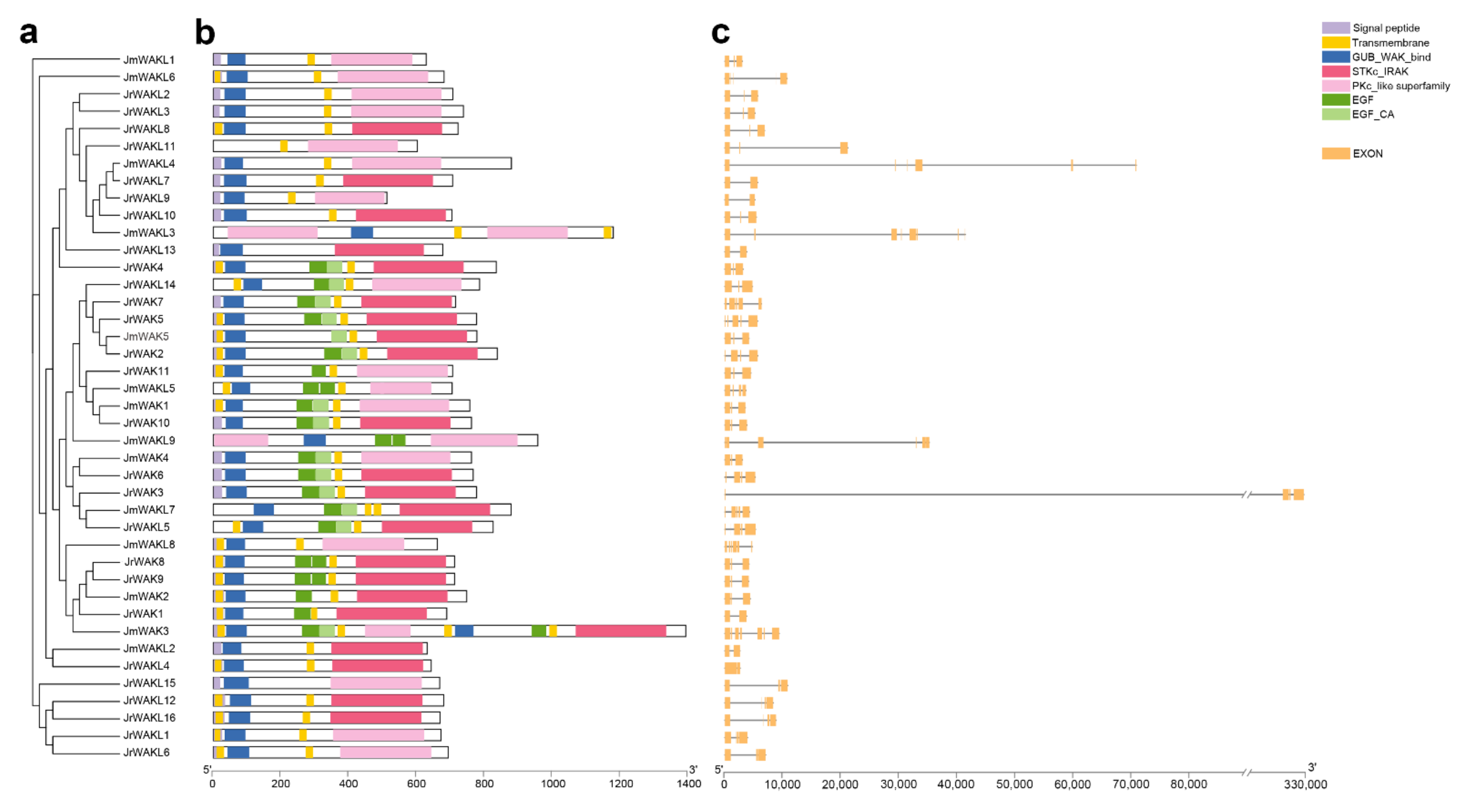
3.3. Protein Domain and Gene Structure Analysis of the WAK/WAKL Members in J. regia and Its Wild Related Species J. mandshurica
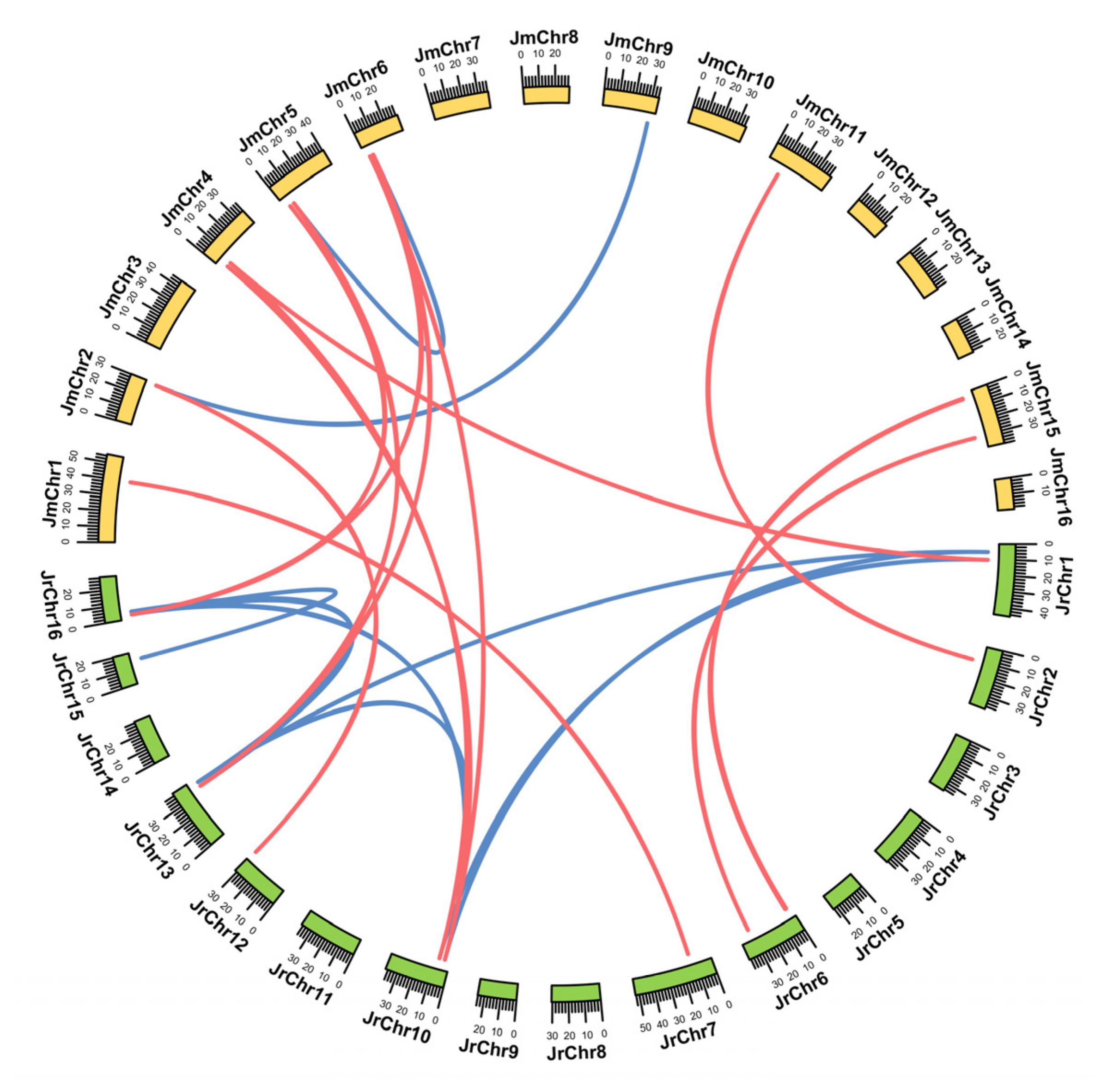
3.4. Chromosomal Distribution and Duplication Mode Analysis of the WAK/WAKL Gene Family in J. regia and Its Wild Related Species J. mandshurica
3.5. Collinearity and Selective Pressure Analysis of the WAK/WAKL Members in J. regia and Its Wild Related Species J. mandshurica
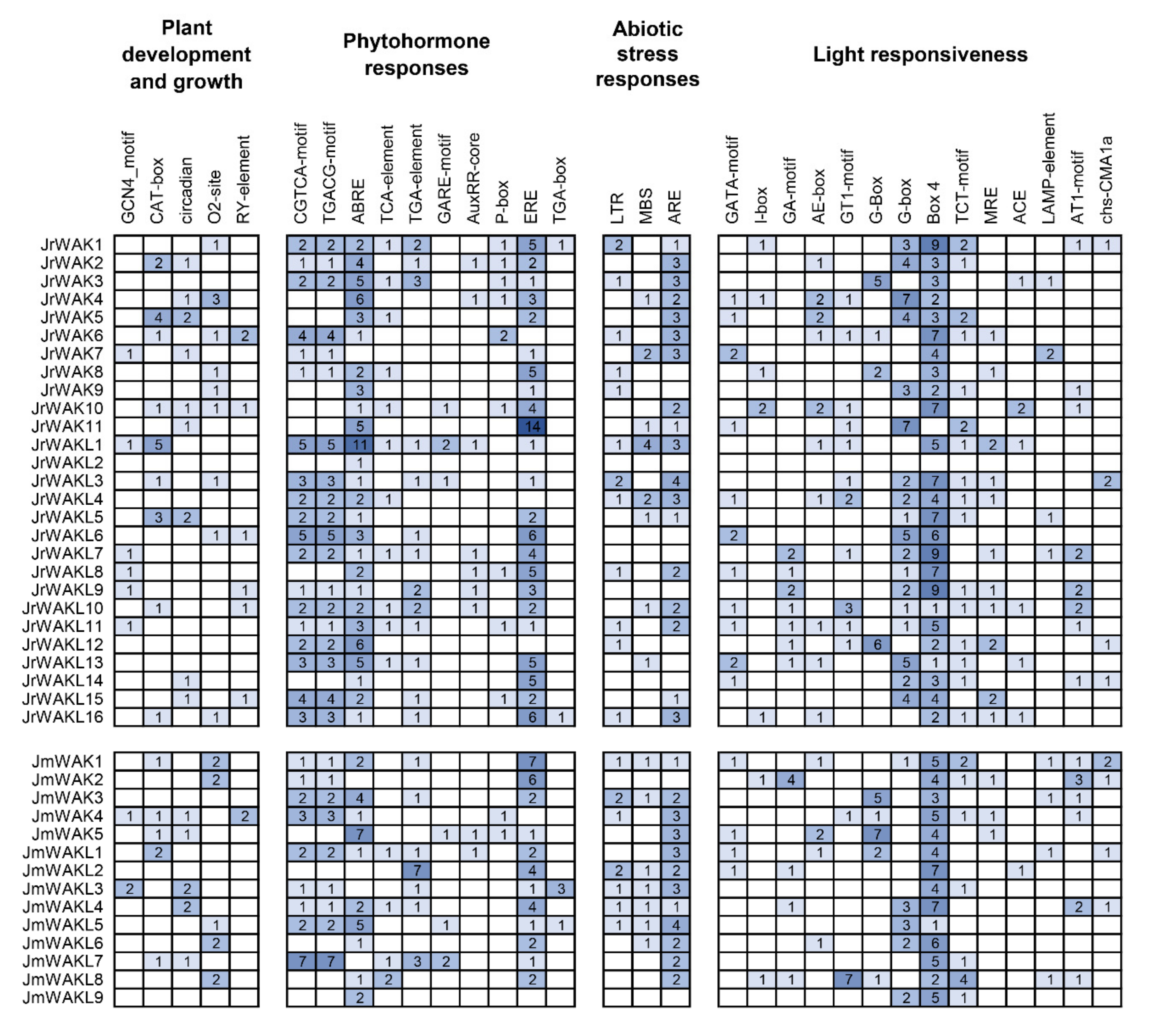
3.6. Cis-Acting Elements Analysis of Promoters of WAK/WAKL Genes in J. regia and Its Wild Related Species J. mandshurica
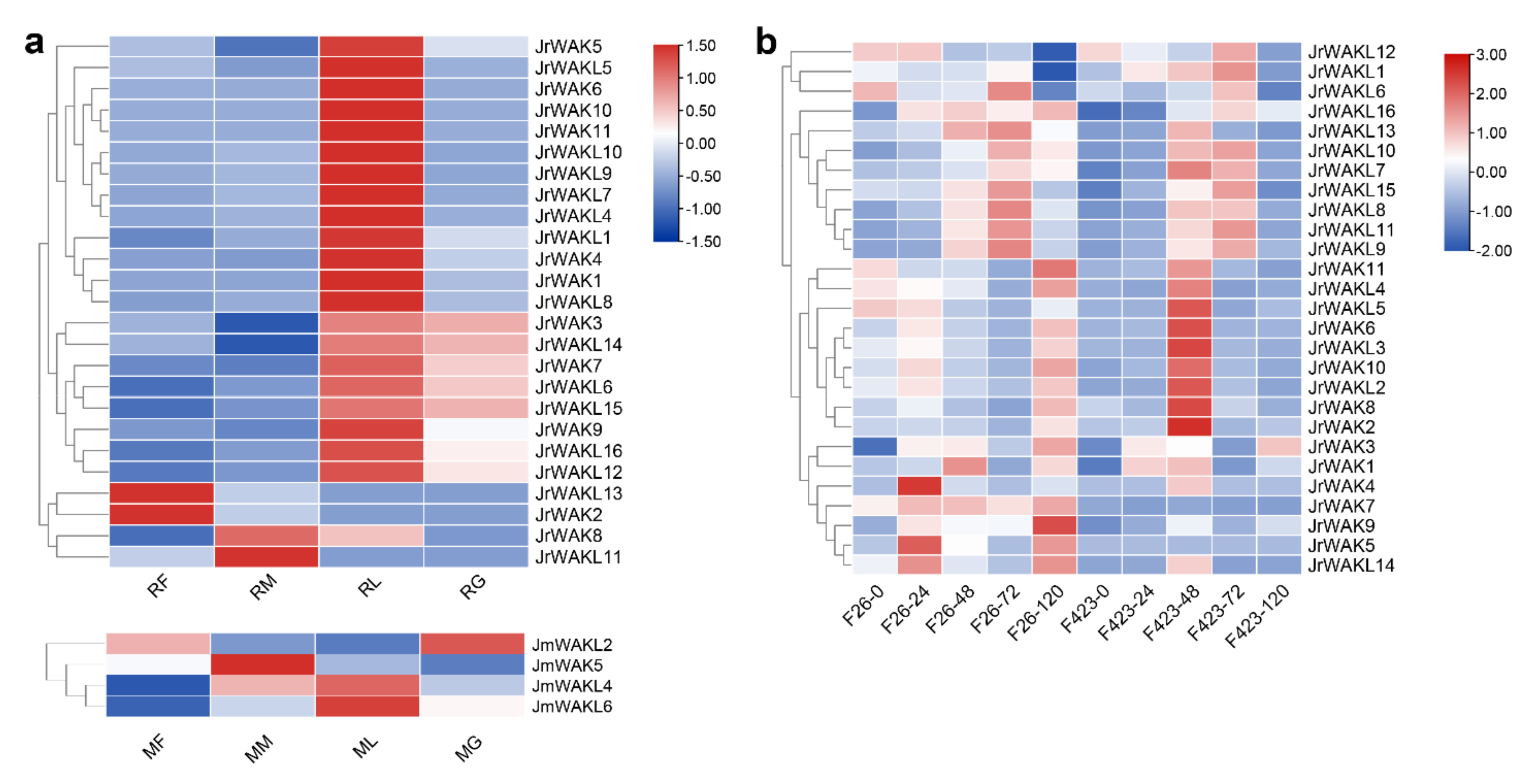
3.7. Expression Profile Analysis of the WAK/WAKL Genes in J. regia and Its Wild Related Species J. mandshurica
4. Discussion
4.1. Characteristics of WAK/WAKL Genes in J. regia and J. mandshurica
4.2. The Tandem Duplication of the WAK/WAKL Gene Family in J. regia and J. mandshurica
4.3. The Conserved Domain, Gene Structure, and cis-Acting Elements of the WAK/WAKL Genes in J. regia and J. mandshurica
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kohorn, B.D.; Kobayashi, M.; Johansen, S.; Riese, J.; Huang, L.F.; Koch, K.; Fu, S.; Dotson, A.; Byers, N. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 2006, 46, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Underwood, W. The plant cell wall: A dynamic barrier against pathogen invasion. Front Plant Sci. 2012, 3, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; Li, Z.; Shen, Q.; Shi, H.; Li, H.; Wang, W.; Zou, C.; Shang, H.; Li, H.; Xiao, G. Genome-wide characterization of the WAK gene family and expression analysis under plant hormone treatment in cotton. BMC. Genom. 2021, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Bleecker, A.B. Receptor-like kinases from Arabidopsis form amonophyletic gene family related to animal receptor kinases. Proc. Natl. Acad. Sci. USA 2001, 98, 10763–10768. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Liu, Y.; Guo, Z.; Mao, J.; Chen, B. Genome-wide annotation and expression responses to biotic stresses of the wall-associated kinase-Rcecptor-like kinase (WAK-RLK) gene family in apple (Malus domestica). Eur. J. Plant Pathol. 2018, 153, 771–785. [Google Scholar] [CrossRef]
- Kohorn, B.D. WAKs; cell wall associated kinases. Curr. Opin. Cell Biol. 2001, 13, 529–533. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, W.; Ren, Z.; Wang, X.; Zhao, J.; Pei, X.; Liu, Y.; He, K.; Zhang, F.; Hou, W.; et al. Characterization and expression analysis of wall-associated kinase (WAK) and WAK-like family in cotton. Int. J. Biol. Macromol. 2021, 187, 867–879. [Google Scholar] [CrossRef]
- Anderson, C.M.; Wagner, T.A.; Perret, M.; He, Z.H.; He, D.; Kohorn, B.D. WAKs: Cell wall-associated kinases linking the cytoplasm to the extracellular matrix. Plant Mol. Biol. 2001, 47, 197–206. [Google Scholar] [CrossRef]
- Sivaguru, M.; Ezaki, B.; He, Z.H.; Tong, H.; Osawa, H.; Baluska, F.; Volkmann, D.; Matsumoto, H. Aluminum-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003, 132, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Zhou, L.; Jamieson, P.; Zhang, L.; Zhao, Z.; Babilonia, K.; Shao, W.; Wu, L.; Mustafa, R.; Amin, I.; et al. The cotton wall-associated kinase GhWAK7A mediates responses to fungal wilt pathogens by complexing with the chitin sensory receptors. Plant Cell. 2020, 32, 3978–4001. [Google Scholar] [CrossRef]
- Qi, H.; Zhu, X.; Guo, F.; Lv, L.; Zhang, Z. The wall-associated receptor-like kinase TaWAK7D is required for defense responses to Rhizoctonia cerealis in Wheat. Int. J. Mol. Sci. 2020, 22, 5629. [Google Scholar] [CrossRef]
- Verica, J.A.; He, Z.H. The cell wall-associated kinase (WAK) and WAK-like kinase gene family. Plant Physiol. 2002, 129, 455–459. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, L.F.V.; Christoff, A.P.; de Lima, J.C.; de Ross, B.C.F.; Sachetto-Martins, G.; Margis-Pinherio, M.; Margis, R. The wall-associated kinase gene family in rice genomes. Plant Sci. 2014, 229, 181–192. [Google Scholar] [CrossRef]
- Hou, X.; Tong, H.; Selby, J.; Dewitt, J.; Peng, X.; He, Z.H. Involvement of a cell wall-associated kinase, WAKL4, in Arabidopsis mineral responses. Plant Physiol. 2005, 139, 1704–1716. [Google Scholar] [CrossRef] [Green Version]
- Lally, D.; Ingmire, P.; Tong, H.Y.; He, Z.H. Antisense expression of a cell wall-associated protein kinase, WAK4, inhibits cell elongation and alters morphology. Plant Cell. 2001, 13, 1317–1331. [Google Scholar] [CrossRef]
- Wagner, T.A.; Kohorn, B.D. Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell. 2001, 13, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhou, S.Y.; Zhao, W.S.; Su, S.C.; Peng, Y.L. A novel wall-associated receptor-like protein kinase gene, OsWAK1, plays important roles in rice blast disease resistance. Plant Mol. Biol. 2009, 69, 337–346. [Google Scholar] [CrossRef]
- Feng, H.; Li, C.; Zhou, J.; Yuan, Y.; Feng, Z.; Shi, Y.; Zhao, L.; Zhang, Y.; Wei, F.; Zhu, H. A cotton WAKL protein interacted with a DnaJ protein and was involved in defense against Verticillium dahliae. Int. J. Biol. Macromol. 2021, 167, 633–643. [Google Scholar] [CrossRef]
- Yang, P.; Praz, C.; Li, B.; Singla, J.; Robert, C.; Kessel, B.; Scheuermann, D.; Lüthi, L.; Ouzunova, M.; Erb, M.; et al. Fungal resistance mediated by maize wall-associated kinase ZmWAK-RLK1 correlates with reduced benzoxazinoid content. New Phytol. 2019, 221, 976–987. [Google Scholar] [CrossRef] [Green Version]
- Saintenac, C.; Lee, W.S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Powers, S.J.; Bergès, H.; Phillips, A.L.; Uauy, C.; et al. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat. Genet. 2018, 50, 368–374. [Google Scholar] [CrossRef]
- Martínez-García, P.J.; Crepeau, M.W.; Puiu, D.; Gonzalez-Ibeas, D.; Whalen, J.; Stevens, K.A.; Paul, R.; Butterfield, T.S.; Brittom, M.T.; Reagan, R.L.; et al. The walnut (Juglans regia) genome sequence reveals diversity in genes coding for the biosynthesis of non-structural polyphenols. Plant J. 2016, 87, 507–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrano, A.; Britton, M.; Zaini, P.A.; Zimin, A.V.; Workman, R.E.; Puiu, D.; Bianco, L.; Pierro, E.A.D.; Allen, B.J.; Chakraborty, S.; et al. High-quality chromosome-scale assembly of the walnut (Juglans regia L.) reference genome. Gigascience 2020, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Xi, R.M.; She, R.X.; Chen, P.P.; Yan, Y.J.; Yang, G.; Dang, M.; Yue, M.; Pei, D.; Woestek, K.; et al. Improved de novo chromosome-level genome assembly of the vulnerable walnut tree Juglans mandshurica reveals gene family evolution and possible genome basis of resistance to lesion nematode. Mol. Ecol. Resour. 2021, 21, 2063–2076. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Z.; Tian, Y.; Zhang, S.; Li, D.; Dong, W.; Zhang, C.; Zhang, Z. Characterization of wall-associated kinase/wall-associated kinase-like (WAK/WAKL) family in rose (Rosa chinensis) reveals the role of RcWAK4 in Botrytis resistance. BMC Plant Biol. 2021, 21, 526. [Google Scholar] [CrossRef]
- Tripathi, R.K.; Aguirre, J.A.; Singh, J. Genome-wide analysis of wall associated kinase (WAK) gene family in barley. Genomics 2021, 113, 523–530. [Google Scholar] [CrossRef]
- Plomion, C.; Aury, J.M.; Amselem, J.; Leroy, T.; Murat, F.; Duplessis, S.; Faye, S.; Francillonne, N.; Labadie, K.; Le Provost, G.; et al. Oak genome reveals facets of long lifespan. Nat. Plants 2018, 4, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Gou, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Feng, H.; Liu, X.; Dong, Y.; Wang, Q.; Yang, K.Q. Genome-wide identification and characterization of long non-coding RNAs conferring resistance to Colletotrichum gloeosporioides in walnut (Juglans regia). BMC Genom. 2021, 22, 15. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, V.; Gupta, A.K. Wall associated kinases from plants—An overview. Physiol. Mol. Biol. Plants 2008, 14, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Song, Y.; Chen, D.; Zhang, Y.; Zhang, Q.; Yi, Y.; Qu, G. Genome-wide identification, classification, characterization, and expression analysis of the wall-associated kinase family during fruit development and under wound stress in tomato (Solanum lycopersicum L.). Genes 2020, 11, 1186. [Google Scholar] [CrossRef]
- Liang, Y.; Wan, N.; Cheng, Z.; Mo, Y.; Liu, B.; Liu, H.; Raboanatahiry, N.; Yin, Y.; Li, M. Whole-genome identification and expression pattern of the vicinal oxygen chelate family in rapeseed (Brassica napus L.). Front. Plant Sci. 2017, 8, 745. [Google Scholar] [CrossRef] [Green Version]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Shiu, S.H.; Karlowski, W.M.; Pan, R.; Tzeng, Y.H.; Mayer, K.F.; Li, W.H. Comparative analysis of the receptor-like kinase family in Arabidopsis and rice. Plant Cell. 2004, 16, 1220–1234. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [Green Version]
- Roulin, A.; Auer, P.L.; Libault, M.; Schlueter, J.; Farmer, A.; May, G.; Stacey, G.; Doerge, R.W.; Jackson, S.A. The fate of duplicated genes in a polyploid plant genome. Plant J. 2013, 73, 143–153. [Google Scholar] [CrossRef]
- Verica, J.A.; Chae, L.; Tong, H.; Ingmire, P.; He, Z.H. Tissue-specific and developmentally regulated expression of a cluster of tandemly arrayed cell wall-associated kinase-like kinase genes in Arabidopsis. Plant Physiol 2003, 133, 1732–1746. [Google Scholar] [CrossRef] [Green Version]
- Jo, B.S.; Choi, S.S. Introns: The functional benefits of introns in genomes. Genom. Inform. 2015, 13, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukheriee, D.; Saha, D.; Acharya, D.; Mukheriee, A.; Chakraborty, S.; Ghosh, T.C. The role of introns in the conservation of the metabolic genes of Arabidopsis thaliana. Genomics 2018, 110, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Kalsotra, A.; Cooper, T.A. Functional consequences of developmentally regulated alternative splicing. Nat. Rev. Genet. 2011, 12, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.Y.; Lin, S.L. Intronic microRNAs. Biochem. Biophys. Res. Commun. 2005, 326, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Liu, B.; Liu, L.; Song, S. Jasmonate action in plant growth and development. J. Exp. Bot. 2017, 68, 1349–1359. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | No. of Amino Acids | Mol. Wt (kDa) | Isoelectric Point (pI) | Instability Index (II) | Aliphatic Index | Grand Average of Hydropathicity (GRAVY) | Subcellular Localizationa |
|---|---|---|---|---|---|---|---|
| JrWAK1 | 690 | 77,758.55 | 5.97 | 40.56 | 80.94 | −0.31 | plas |
| JrWAK2 | 839 | 91,544.55 | 6.36 | 33.11 | 82.81 | −0.33 | plas |
| JrWAK3 | 778 | 85,990.06 | 6.36 | 37.49 | 92.57 | −0.19 | vacu |
| JrWAK4 | 836 | 92,128.95 | 6.67 | 35.95 | 83.83 | −0.29 | plas |
| JrWAK5 | 778 | 85,292.05 | 6.25 | 31.33 | 87.90 | −0.21 | vacu |
| JrWAK6 | 768 | 85,036.38 | 6.96 | 37.39 | 91.76 | −0.19 | plas |
| JrWAK7 | 716 | 79,111.38 | 8.12 | 34.46 | 89.15 | −0.26 | vacu |
| JrWAK8 | 713 | 79,985.06 | 5.93 | 36.64 | 83.52 | −0.22 | vacu |
| JrWAK9 | 713 | 79,776.70 | 5.64 | 33.72 | 82.85 | −0.24 | vacu |
| JrWAK10 | 763 | 84,326.97 | 5.43 | 36.41 | 84.84 | −0.20 | plas |
| JrWAK11 | 707 | 78,434.90 | 8.48 | 42.21 | 87.96 | −0.21 | plas |
| JrWAKL1 | 673 | 74,644.43 | 5.27 | 43.53 | 87.74 | −0.16 | extr |
| JrWAKL2 | 708 | 79,775.49 | 7.86 | 38.86 | 84.66 | −0.23 | chlo |
| JrWAKL3 | 739 | 83,259.24 | 7.71 | 36.16 | 84.79 | −0.26 | extr |
| JrWAKL4 | 644 | 70,964.82 | 8.83 | 37.45 | 92.34 | −0.13 | vacu |
| JrWAKL5 | 827 | 91,687.81 | 7.63 | 36.59 | 90.50 | −0.16 | plas |
| JrWAKL6 | 694 | 76,326.10 | 5.56 | 36.81 | 83.16 | −0.18 | plas |
| JrWAKL7 | 707 | 78,717.69 | 6.33 | 32.77 | 86.53 | −0.14 | plas |
| JrWAKL8 | 723 | 81,305.71 | 6.01 | 33.79 | 81.29 | −0.23 | plas |
| JrWAKL9 | 513 | 57,573.86 | 7.82 | 27.57 | 86.80 | −0.08 | chlo |
| JrWAKL10 | 705 | 79,273.50 | 6.54 | 28.82 | 85.97 | −0.18 | plas |
| JrWAKL11 | 603 | 67,641.00 | 7.76 | 31.67 | 82.12 | −0.35 | nucl |
| JrWAKL12 | 681 | 76,621.84 | 6.37 | 48.65 | 83.85 | −0.33 | plas |
| JrWAKL13 | 678 | 75,503.57 | 5.91 | 43.32 | 82.05 | −0.24 | chlo |
| JrWAKL14 | 787 | 87,360.88 | 6.51 | 33.75 | 83.60 | −0.23 | plas |
| JrWAKL15 | 669 | 74,597.36 | 5.94 | 39.87 | 81.46 | −0.22 | vacu |
| JrWAKL16 | 670 | 74,773.49 | 7.78 | 46.52 | 82.19 | −0.30 | plas |
| JmWAK1 | 758 | 84,080.04 | 6.35 | 35.01 | 84.37 | −0.21 | plas |
| JmWAK2 | 749 | 83,946.50 | 5.86 | 35.86 | 82.26 | −0.26 | vacu |
| JmWAK3 | 1396 | 155,950.00 | 7.06 | 38.40 | 87.49 | −0.25 | plas |
| JmWAK4 | 763 | 84,642.55 | 7.88 | 40.86 | 86.09 | −0.27 | plas |
| JmWAK5 | 779 | 85,485.49 | 6.56 | 35.61 | 88.19 | −0.22 | plas |
| JmWAKL1 | 629 | 69,698.84 | 6.64 | 51.86 | 94.10 | 0.01 | plas |
| JmWAKL2 | 632 | 69,194.91 | 6.20 | 36.32 | 87.66 | −0.16 | plas |
| JmWAKL3 | 1182 | 133,611.68 | 6.61 | 32.72 | 88.15 | −0.22 | plas |
| JmWAKL4 | 881 | 99,728.80 | 6.22 | 35.90 | 85.27 | −0.23 | chlo |
| JmWAKL5 | 706 | 78,535.72 | 4.86 | 36.53 | 83.81 | −0.21 | plas |
| JmWAKL6 | 682 | 78,128.27 | 5.79 | 52.94 | 83.05 | −0.22 | plas |
| JmWAKL7 | 880 | 98,454.69 | 8.40 | 39.81 | 90.69 | −0.23 | plas |
| JmWAKL8 | 662 | 73,877.35 | 7.11 | 42.40 | 88.34 | −0.30 | plas |
| JmWAKL9 | 959 | 106,500.53 | 5.56 | 40.59 | 95.62 | −0.14 | plas |
| Gene Name | Whole Genome Duplication (WGD) | Tandem Duplication (TD) | Dispersed Duplication (DSD) | Proximal Duplication (PD) |
|---|---|---|---|---|
| JrWAK1 | √ | |||
| JrWAK2 | √ | |||
| JrWAK3 | √ | |||
| JrWAK4 | √ | |||
| JrWAK5 | √ | |||
| JrWAK6 | √ | |||
| JrWAK7 | √ | |||
| JrWAK8 | √ | |||
| JrWAK9 | √ | |||
| JrWAK10 | √ | √ | ||
| JrWAK11 | √ | |||
| JrWAKL1 | ||||
| JrWAKL2 | √ | |||
| JrWAKL3 | √ | |||
| JrWAKL4 | √ | |||
| JrWAKL5 | √ | |||
| JrWAKL6 | √ | √ | ||
| JrWAKL7 | √ | |||
| JrWAKL8 | √ | |||
| JrWAKL9 | √ | |||
| JrWAKL10 | √ | |||
| JrWAKL11 | √ | |||
| JrWAKL12 | √ | |||
| JrWAKL13 | √ | |||
| JrWAKL14 | √ | |||
| JrWAKL15 | ||||
| JrWAKL16 | √ | |||
| JmWAK1 | √ | |||
| JmWAK2 | √ | |||
| JmWAK3 | √ | |||
| JmWAK4 | √ | |||
| JmWAK5 | √ | |||
| JmWAKL1 | √ | |||
| JmWAKL2 | √ | |||
| JmWAKL3 | √ | |||
| JmWAKL4 | √ | |||
| JmWAKL5 | √ | |||
| JmWAKL6 | √ | |||
| JmWAKL7 | √ | |||
| JmWAKL8 | √ | |||
| JmWAKL9 | √ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Ma, J.; Liu, H.; Ou, M.; Ye, H.; Zhao, P. Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica. Genes 2022, 13, 134. https://doi.org/10.3390/genes13010134
Li M, Ma J, Liu H, Ou M, Ye H, Zhao P. Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica. Genes. 2022; 13(1):134. https://doi.org/10.3390/genes13010134
Chicago/Turabian StyleLi, Mengdi, Jiayu Ma, Hengzhao Liu, Mengwei Ou, Hang Ye, and Peng Zhao. 2022. "Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica" Genes 13, no. 1: 134. https://doi.org/10.3390/genes13010134