
Years of Schooling Could Reduce Epigenetic Aging: A Study of a Mexican Cohort

 ,
, 
 , and
, and
Abstract
:1. Introduction
1.1. Social Epigenetics
1.2. Tlaltizapan Cohort
2. Materials and Methods
2.1. Sample Population
2.2. Clinical and Anthropometric Measurements
2.2.1. Biochemical Profile
2.2.2. Anthropometric Measurements
2.2.3. Body Composition and Physical Performance
2.2.4. Health Status
2.3. DNA Methylation
2.3.1. Epigenetic Clocks
2.4. Statistical Analysis
2.4.1. Clinical Study, Epigenetic Clocks, and Anthropometric Comparison between the Tlatizapan Cohort and the Urban-Raised Cohort
2.4.2. Analysis Only on the Tlatizapan Cohort
Epigenetic Clocks
Epigenome-Wide Association Analysis
2.5. Bioethical Considerations
3. Results
3.1. Differences between the Urban-Raised and the Tlatizapan Cohorts
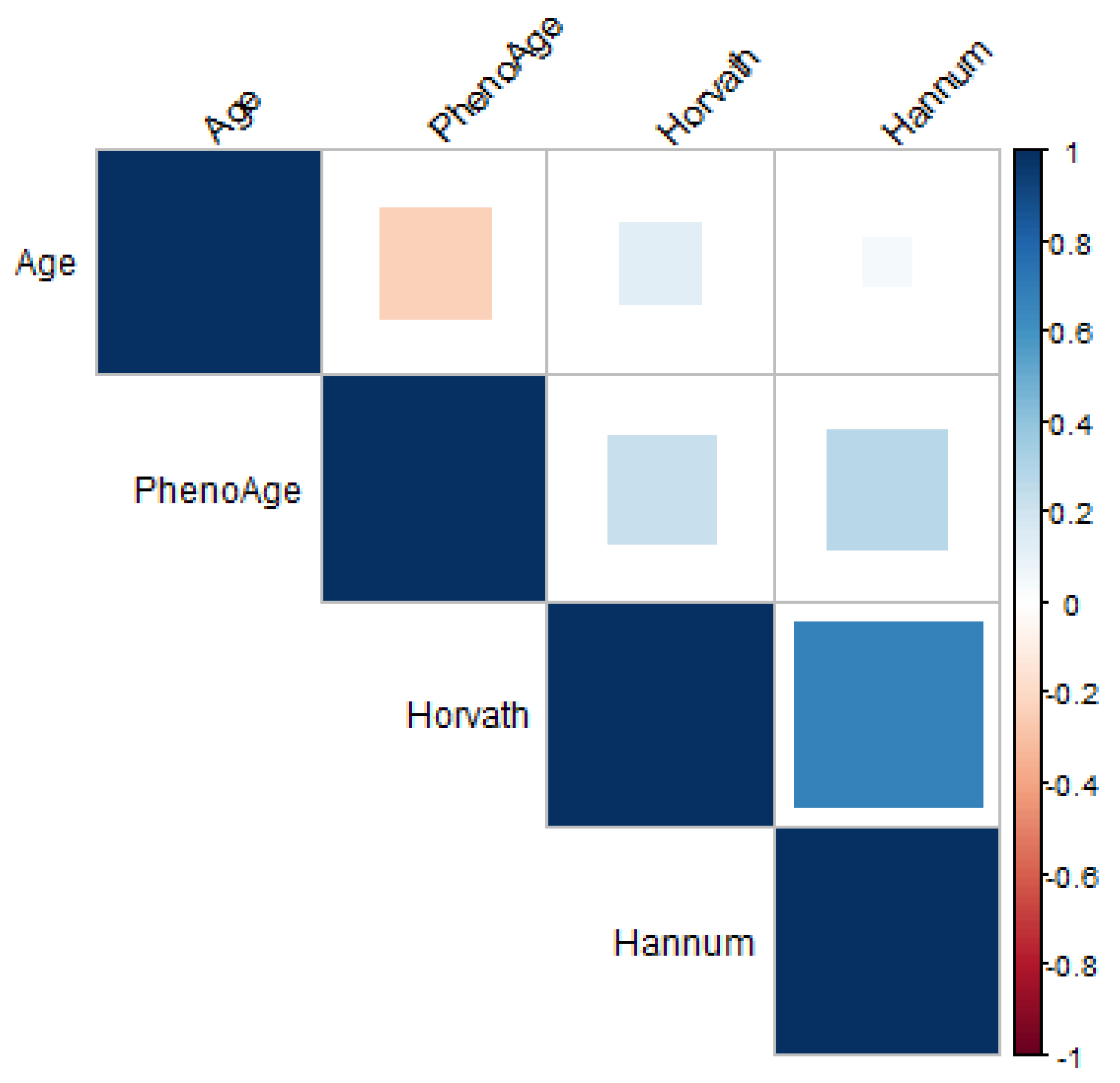
Evaluation of Different Epigenetic Clocks
3.2. Analysis of the Tlatizapan Cohort
3.2.1. Effect of Clinical Variables on Individuals with Accelerated Epigenetic Aging
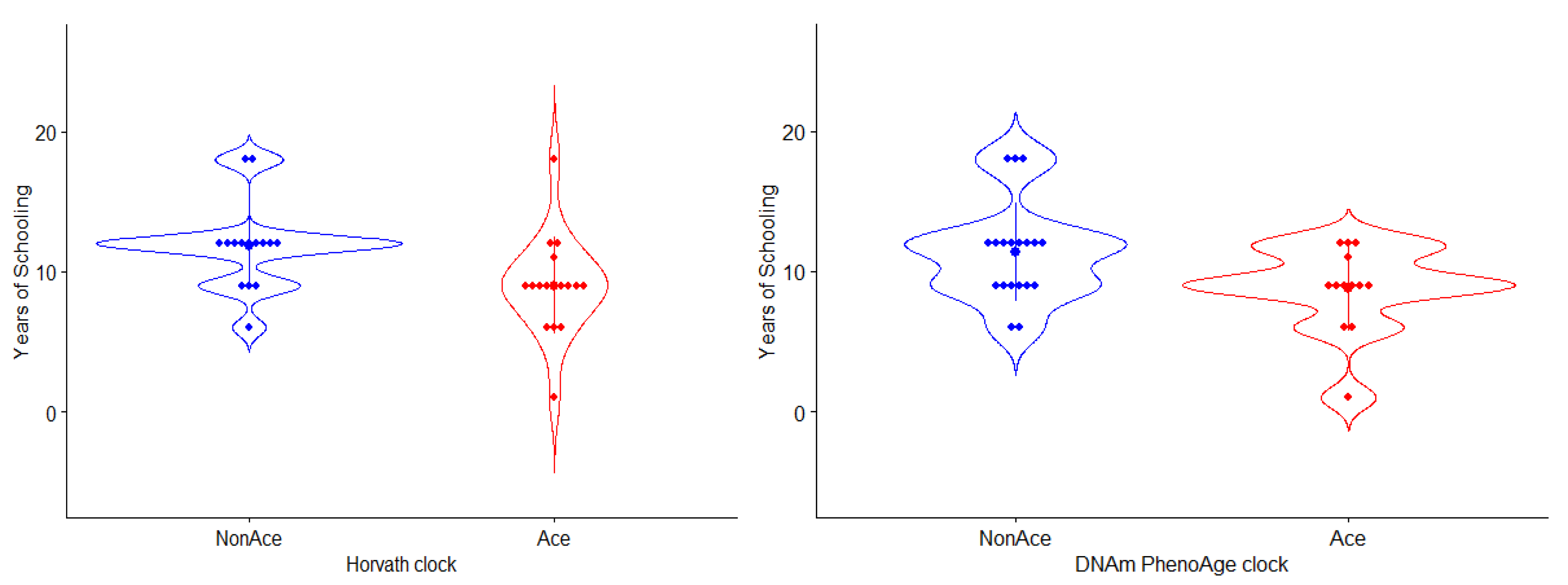
3.2.2. Evaluation of Epigenetic Changes in Individuals with Long and Short Durations of Schooling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gluckman, P.D.; Hanson, M.A.; Spencer, H.G.; Bateson, P. Environmental Influences during Development and Their Later Consequences for Health and Disease: Implications for the Interpretation of Empirical Studies. Proc. R. Soc. B Biol. Sci. 2005, 272, 671–677. [Google Scholar] [CrossRef] [Green Version]
- Schell, L.M.; Gallo, M.V.; Ravenscroft, J. Environmental Influences on Human Growth and Development: Historical Review and Case Study of Contemporary Influences. Ann. Hum. Biol. 2009, 36, 459–477. [Google Scholar] [CrossRef]
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Aging, Inflammation and the Environment. Exp. Gerontol. 2018, 105, 10–18. [Google Scholar] [CrossRef]
- Christensen, K.; McGue, M. Healthy Ageing, the Genome and the Environment. Nat. Rev. Endocrinol. 2016, 12, 378–380. [Google Scholar] [CrossRef] [Green Version]
- Lovely, C.; Rampersad, M.; Fernandes, Y.; Eberhart, J. Gene-Environment Interactions in Development and Disease: Gene-Environment Interactions in Development and Disease. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e247. [Google Scholar] [CrossRef]
- Vrieze, S.I.; Iacono, W.G.; McGue, M. Confluence of Genes, Environment, Development, and Behavior in a Post Genome-Wide Association Study World. Dev. Psychopathol. 2012, 24, 1195–1214. [Google Scholar] [CrossRef] [Green Version]
- Strife, S.; Downey, L. Childhood Development and Access to Nature: A New Direction for Environmental Inequality Research. Organ. Environ. 2009, 22, 99–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Verjan, J.C.; Barrera-Vázquez, O.S.; García-Velázquez, L.; Samper-Ternent, R.; Arroyo, P. Epigenetic Variations Due to Nutritional Status in Early-life and Its Later Impact on Aging and Disease. Clin. Genet. 2020, 98, 313–321. [Google Scholar] [CrossRef] [PubMed]
- McEniry, M. Early-Life Conditions and Older Adult Health in Low- and Middle-Income Countries: A Review. J. Dev. Orig. Health Dis. 2013, 4, 10–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, A.; Shipley, M.; Singh-Manoux, A.; Marmot, M.G. Successful Aging: The Contribution of Early-Life and Midlife Risk Factors: SUCCESSFUL AGING. J. Am. Geriatr. Soc. 2008, 56, 1098–1105. [Google Scholar] [CrossRef] [Green Version]
- Tani, Y.; Fujiwara, T.; Kondo, K. Adverse Childhood Experiences and Dementia: Interactions With Social Capital in the Japan Gerontological Evaluation Study Cohort. Am. J. Prev. Med. 2021, 61, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T. Survival Costs of Reproduction in Drosophila. Exp. Gerontol. 2011, 46, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.N.; Barker, D.J.P. Type 2 (Non-Insulin-Dependent) Diabetes Mellitus: The Thrifty Phenotype Hypothesis. Diabetologia 1992, 35, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Almond, D.; Currie, J. Killing Me Softly: The Fetal Origins Hypothesis. J. Econ. Perspect. 2011, 25, 153–172. [Google Scholar] [CrossRef] [Green Version]
- Tobi, E.W.; Goeman, J.J.; Monajemi, R.; Gu, H.; Putter, H.; Zhang, Y.; Slieker, R.C.; Stok, A.P.; Thijssen, P.E.; Müller, F.; et al. DNA Methylation Signatures Link Prenatal Famine Exposure to Growth and Metabolism. Nat. Commun. 2014, 5, 5592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jowett, A.J. The Growth of China’s Population, 1949–1982 (With Special Reference to the Demographic Disaster of 1960–61). Geogr. J. 1984, 150, 155. [Google Scholar] [CrossRef]
- Jowett, A.J. The Demographic Responses to Famine: The Case of China 1958? 61. GeoJournal 1991, 23. [Google Scholar] [CrossRef]
- Ahmed, F. Epigenetics: Tales of Adversity. Nature 2010, 468, S20. [Google Scholar] [CrossRef] [Green Version]
- Schulz, L.C. The Dutch Hunger Winter and the Developmental Origins of Health and Disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16757–16758. [Google Scholar] [CrossRef] [Green Version]
- Gensous, N.; Franceschi, C.; Santoro, A.; Milazzo, M.; Garagnani, P.; Bacalini, M.G. The Impact of Caloric Restriction on the Epigenetic Signatures of Aging. Int. J. Mol. Sci. 2019, 20, 2022. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Song, J.; Li, Y.; Dong, B.; Zou, Z.; Ma, J. Early-Life Exposure to the Chinese Famine Is Associated with Higher Methylation Level in the INSR Gene in Later Adulthood. Sci. Rep. 2019, 9, 3354. [Google Scholar] [CrossRef]
- Cohen, D.A.; Babey, S. Reducing Environmental Risks to Modify Lifestyle Behaviors. Diabetes Spectr. 2015, 28, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer Is a Preventable Disease That Requires Major Lifestyle Changes. Pharm. Res. 2008, 25, 2097–2116. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H.; Martin, S. Environmental/Lifestyle Factors in the Pathogenesis and Prevention of Type 2 Diabetes. BMC Med. 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Kremers, S.P.J.; Eves, F.F.; Andersen, R.E. Environmental Changes to Promote Physical Activity and Healthy Dietary Behavior. J. Environ. Public Health 2012, 2012, 470858. [Google Scholar] [CrossRef]
- Loi, M.; Del Savio, L.; Stupka, E. Social Epigenetics and Equality of Opportunity. Public Health Ethics 2013, 6, 142–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrill, S.M.; Gladish, N.; Kobor, M.S. Social Environment and Epigenetics. In Behavioral Neurogenomics; Binder, E.B., Klengel, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; Volume 42, pp. 83–126. ISBN 978-3-030-31264-0. [Google Scholar]
- Cunliffe, V.T. The Epigenetic Impacts of Social Stress: How Does Social Adversity Become Biologically Embedded? Epigenomics 2016, 8, 1653–1669. [Google Scholar] [CrossRef] [Green Version]
- Curley, J.P.; Jensen, C.L.; Mashoodh, R.; Champagne, F.A. Social Influences on Neurobiology and Behavior: Epigenetic Effects during Development. Psychoneuroendocrinology 2011, 36, 352–371. [Google Scholar] [CrossRef] [Green Version]
- Moosavi, A.; Motevalizadeh Ardekani, A. Role of Epigenetics in Biology and Human Diseases. Iran. Biomed. J. 2016, 20, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Barouki, R.; Melén, E.; Herceg, Z.; Beckers, J.; Chen, J.; Karagas, M.; Puga, A.; Xia, Y.; Chadwick, L.; Yan, W.; et al. Epigenetics as a Mechanism Linking Developmental Exposures to Long-Term Toxicity. Environ. Int. 2018, 114, 77–86. [Google Scholar] [CrossRef]
- BIOS Consortium; van Dongen, J.; Bonder, M.J.; Dekkers, K.F.; Nivard, M.G.; van Iterson, M.; Willemsen, G.; Beekman, M.; van der Spek, A.; van Meurs, J.B.J.; et al. DNA Methylation Signatures of Educational Attainment. NPJ Sci. Learn. 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BIOS Consortium; Karlsson Linnér, R.; Marioni, R.E.; Rietveld, C.A.; Simpkin, A.J.; Davies, N.M.; Watanabe, K.; Armstrong, N.J.; Auro, K.; Baumbach, C.; et al. An Epigenome-Wide Association Study Meta-Analysis of Educational Attainment. Mol. Psychiatry 2017, 22, 1680–1690. [Google Scholar] [CrossRef] [Green Version]
- Pickersgill, M. Epigenetics, Education, and the Plastic Body: Changing Concepts and New Engagements. Res. Educ. 2020, 107, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Hu, H. The Relationship and Mechanism between Education and Functional Health Status Transition among Older Persons in China. BMC Geriatr. 2018, 18, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-Wide Methylation Profiles Reveal Quantitative Views of Human Aging Rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA Methylation Age of Human Tissues and Cell Types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, A.T.; Quach, A.; Wilson, J.G.; Reiner, A.P.; Aviv, A.; Raj, K.; Hou, L.; Baccarelli, A.A.; Li, Y.; Stewart, J.D.; et al. DNA Methylation GrimAge Strongly Predicts Lifespan and Healthspan. Aging 2019, 11, 303–327. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An Epigenetic Biomarker of Aging for Lifespan and Healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E. Assessment of Epigenetic Clocks as Biomarkers of Aging in Basic and Population Research. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 463–465. [Google Scholar] [CrossRef]
- Cravioto, J.; Birch, H.; De Licardie, E.; Rosales, L.; Vega, L. The Ecology of Growth and Development in a Mexican Preindustrial Community Report 1: Method and Findings from Birth to One Month of Age. Monogr. Soc. Res. Child Dev. 1969, 34, 1–76. [Google Scholar] [CrossRef]
- Galvan-Ramos, R.; Cravioto, J.; Gutierrez, G.; Gomez, F.; Frenk, S. Operation Zacatepec. III. Comparison of an Indirect and a Direct Method in Evaluation of the Nutritional Status of Children of a Rural Community. Bol. Méd. Hosp. Infant. México 1958, 15, 855–873. [Google Scholar]
- Vega Franco, L.; Cravioto, J.; Roca de Licardie, E. Ecology of the Size of Newborn Infants in a Preindustrial Community. Bol. Méd. Hosp. Infant. México 1974, 31, 199–206. [Google Scholar]
- Sandler, S.G. Management of Patients with Hematologic Malignancies and Aplastic Anemia Who Are Refractory to Platelet Transfusions. Haematol. Budap 1998, 29, 1–11. [Google Scholar]
- World Health Organization. Prevention of Diabetes Mellitus. Report of a WHO Study Group; World Health Organization: Geneva, Switzerland, 1994; Volume 844, pp. 1–100. [Google Scholar]
- Muñoz, O.M.; García, Á.A.; Fernández-Ávila, D.; Higuera, A.; Ruiz, Á.J.; Aschner, P.; Toro, J.M.; Arteaga, J.M.; Merchán, A.; Sánchez, G.; et al. Guía de práctica clínica para la prevención, detección temprana, diagnóstico, tratamiento y seguimiento de las dislipidemias: Evaluación del riesgo cardiovascular. Rev. Colomb. Cardiol. 2015, 22, 263–269. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Physical Status: The Use and Interpretation of Anthropometry. Report of a WHO Expert Committee; World Health Organization: Geneva, Switzerland, 1995; Volume 854, pp. 1–452. [Google Scholar]
- Shepherd, J.A.; Ng, B.K.; Sommer, M.J.; Heymsfield, S.B. Body Composition by DXA. Bone 2017, 104, 101–105. [Google Scholar] [CrossRef]
- Guralnik, J.M.; Simonsick, E.M.; Ferrucci, L.; Glynn, R.J.; Berkman, L.F.; Blazer, D.G.; Scherr, P.A.; Wallace, R.B. A Short Physical Performance Battery Assessing Lower Extremity Function: Association With Self-Reported Disability and Prediction of Mortality and Nursing Home Admission. J. Gerontol. 1994, 49, M85–M94. [Google Scholar] [CrossRef]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State”. A Practical Method for Grading the Cognitive State of Patients for the Clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Delbaere, K.; Close, J.C.T.; Mikolaizak, A.S.; Sachdev, P.S.; Brodaty, H.; Lord, S.R. The Falls Efficacy Scale International (FES-I). A Comprehensive Longitudinal Validation Study. Age Ageing 2010, 39, 210–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Laird, P.W.; Shen, H. Comprehensive Characterization, Annotation and Innovative Use of Infinium DNA Methylation BeadChip Probes. Nucleic Acids Res. 2016, 45, e22. [Google Scholar] [CrossRef]
- Nordlund, J.; Bäcklin, C.L.; Wahlberg, P.; Busche, S.; Berglund, E.C.; Eloranta, M.-L.; Flaegstad, T.; Forestier, E.; Frost, B.-M.; Harila-Saari, A.; et al. Genome-Wide Signatures of Differential DNA Methylation in Pediatric Acute Lymphoblastic Leukemia. Genome Biol. 2013, 14, r105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pidsley, R.; Zotenko, E.; Peters, T.J.; Lawrence, M.G.; Risbridger, G.P.; Molloy, P.; Van Djik, S.; Muhlhausler, B.; Stirzaker, C.; Clark, S.J. Critical Evaluation of the Illumina MethylationEPIC BeadChip Microarray for Whole-Genome DNA Methylation Profiling. Genome Biol. 2016, 17, 208. [Google Scholar] [CrossRef] [Green Version]
- Morris, T.J.; Butcher, L.M.; Feber, A.; Teschendorff, A.E.; Chakravarthy, A.R.; Wojdacz, T.K.; Beck, S. ChAMP: 450k Chip Analysis Methylation Pipeline. Bioinforma. Oxf. Engl. 2014, 30, 428–430. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Du, P.; Zhang, X.; Huang, C.-C.; Jafari, N.; Kibbe, W.A.; Hou, L.; Lin, S.M. Comparison of Beta-Value and M-Value Methods for Quantifying Methylation Levels by Microarray Analysis. BMC Bioinform. 2010, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansell, G.; Gorrie-Stone, T.J.; Bao, Y.; Kumari, M.; Schalkwyk, L.S.; Mill, J.; Hannon, E. Guidance for DNA Methylation Studies: Statistical Insights from the Illumina EPIC Array. BMC Genom. 2019, 20, 366. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene Set Analysis Toolkit with Revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- McGuinness, D.; McGlynn, L.M.; Johnson, P.C.; MacIntyre, A.; Batty, G.D.; Burns, H.; Cavanagh, J.; Deans, K.A.; Ford, I.; McConnachie, A.; et al. Socio-Economic Status Is Associated with Epigenetic Differences in the PSoBid Cohort. Int. J. Epidemiol. 2012, 41, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perng, W.; Rozek, L.S.; Mora-Plazas, M.; Duchin, O.; Marin, C.; Forero, Y.; Baylin, A.; Villamor, E. Micronutrient Status and Global DNA Methylation in School-Age Children. Epigenetics 2012, 7, 1133–1141. [Google Scholar] [CrossRef]
- Chen, Y.; Li, Y.; Hsieh, T.; Wang, C.; Cheng, K.; Wang, L.; Lin, T.; Cheung, C.H.A.; Wu, C.; Chiang, H. Aging-induced Akt Activation Involves in Aging-related Pathologies and Aβ-induced Toxicity. Aging Cell 2019, 18. [Google Scholar] [CrossRef]
- Griffin, R.J.; Moloney, A.; Kelliher, M.; Johnston, J.A.; Ravid, R.; Dockery, P.; O’Connor, R.; O’Neill, C. Activation of Akt/PKB, Increased Phosphorylation of Akt Substrates and Loss and Altered Distribution of Akt and PTEN Are Features of Alzheimer’s Disease Pathology. J. Neurochem. 2005, 93, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Nyeko, R.; Kalyesubula, I.; Mworozi, E.; Bachou, H. Lactose Intolerance among Severely Malnourished Children with Diarrhoea Admitted to the Nutrition Unit, Mulago Hospital, Uganda. BMC Pediatr. 2010, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, B.; Dudley, M.; Nichols, V.; Putman, M.; Avery, S.; Fraley, J.; Quaroni, A.; Shiner, M.; Carrazza, F. Effects of Malnutrition on Expression and Activity of Lactase in Children. Gastroenterology 1997, 112, 742–751. [Google Scholar] [CrossRef]
- Horvath, S.; Gurven, M.; Levine, M.E.; Trumble, B.C.; Kaplan, H.; Allayee, H.; Ritz, B.R.; Chen, B.; Lu, A.T.; Rickabaugh, T.M.; et al. An Epigenetic Clock Analysis of Race/Ethnicity, Sex, and Coronary Heart Disease. Genome Biol. 2016, 17, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Tlaltizapan Cohort (n = 32) | Urban Raised (n = 7) | Stat (p-Value) | |
|---|---|---|---|
| Age | 52.26 (0.42) | 62.41 (0.35) | 3.98 (0.0105) |
| Gender | |||
| Female | 19 (59.37) | 5 (71.43) | 0.03 (0.8690) |
| Male | 13 (40.63) | 2 (28.57) | 0.03 (0.8690) |
| Years of schooling | 10.31 (3.55) | 13.86 (8.99) | 1.03 (0.3421) |
| BMI | 29.35 (4.61) | 29.12 (4.61) | −0.12 (0.9063) |
| Visceral fat | 3.09 (1.01) | 3.17 (0.86) | 0.21 (0.8379) |
| Biochemical variables | |||
| Glucose | 108.34 (49.87) | 125.43 (69.01) | 0.62 (0.5535) |
| Triglycerides | 212.59 (210.05) | 230.71 (190.92) | 0.22 (0.8281) |
| Total Cholesterol | 196.75 (29.94) | 236.29 (33.14) | 2.91 (0.0189) |
| HDL | 44.07 (12.84) | 45.81 (12.09) | 0.34 (0.7400) |
| LDL | 111.98 (38.60) | 144.30 (44.98) | 1.76 (0.1155) |
| Creatinine | 0.64 (0.18) | 0.71 (0.14) | 1.15 (0.2763) |
| Uric Acid | 5.62 (0.65) | 5.21 (1.42) | −1.17 (0.2535) |
| Prealbumine | 24.92 (9.53) | 28.82 (5.61) | 1.37 (0.1970) |
| Reactive Protein C | 0.40 (0.17) | 0.38 (0.08) | −0.55 (0.5935) |
| Transferrine | 247.76 (87.39) | 276.40 (42.01) | 1.24 (0.2341) |
| Glycosylated Haemoglobin | 6.45 (1.77) | 5.73 (1.22) | −1.22 (0.2525) |
| Fibrinogen | 330.75 (50.13) | 353.83 (24.79) | 1.71 (0.1078) |
| Iron Fixing | 357.06 (60.39) | 388.67 (78.15) | 0.94 (0.3828) |
| Mental Health | |||
| Mini-mental | 25.56 (2.65) | 26.42 (1.81) | 1.04 (0.3167) |
| Depression | 6 (18.75) | 2 (28.57) | 0.00 (0.9472) |
| Drugs | 9 (28.13) | 5 (71.43) | 5.18 (0.1591) |
| Tobacco Index | 1.50 (5.45) | 0.86 (1.18) | −0.61 (0.5477) |
| Alcohol | 28 (87.50) | 4 (57.14) | 1.83 (0.1763) |
| Epigenetic Clocks | |||
| Horvath | 58.37 (3.61) | 63.84 (4.56) | 2.96 (0.0138) |
| Horvath acceleration | −0.14 (3.56) | 0.71 (4.24) | 0.47 (0.6579) |
| Hannum | 56.77 (4.02) | 60.16 (5.40) | 1.75 (0.1096) |
| Hannum acceleration | 0.05 (3.60) | −0.25 (4.45) | −0.15 (0.8814) |
| DNAm PhenoAge | 49.95 (3.62) | 54.74 (4.87) | 2.06 (0.0591) |
| PhenoAge acceleration | 0.11 (6.64) | −0.58 (4.02) | −0.34 (0.7383) |
| 1 Position | CpG Site | 2 LogFC | p-Value | High Avg | Low Avg | Gene | 3 Gene Loc | CGI 4 |
|---|---|---|---|---|---|---|---|---|
| 2:38496264 | cg19269093 | −0.0453 | 2.1563 × 10−5 | 0.8747 | 0.8294 | IGR | OpenSea | |
| 2:136595281 | cg04750100 | −0.0892 | 2.5258 × 10−5 | 0.4171 | 0.3279 | LCT | TSS1500 | OpenSea |
| 2:237163447 | cg25305153 | −0.0779 | 3.3160 × 10−5 | 0.8126 | 0.7347 | ASB18 | Body | OpenSea |
| 3:65561644 | cg05244979 | −0.0510 | 3.4089 × 10−5 | 0.6869 | 0.6359 | MAGI1 | Body | OpenSea |
| 4:113970506 | cg02815171 | 0.1165 | 4.5334 × 10−5 | 0.4218 | 0.5383 | ANK2 | TSS1500 | OpenSea |
| 7:156716133 | cg03184819 | −0.0907 | 4.9140 × 10−5 | 0.2529 | 0.1622 | IGR | OpenSea | |
| 9:4435234 | cg08538646 | −0.0717 | 4.2326 × 10−5 | 0.8585 | 0.7868 | IGR | OpenSea | |
| 9:117818174 | cg07712264 | −0.0555 | 4.0462 × 10−5 | 0.7059 | 0.6504 | TNC | Body | OpenSea |
| 10:25460855 | cg15018193 | −0.0439 | 1.1161 × 10−5 | 0.7583 | 0.7144 | LOC100128811 | Body | Self |
| 11:102124935 | cg06458665 | −0.0397 | 2.2954 × 10−5 | 0.8635 | 0.8237 | IGR | OpenSea | |
| 12:26451968 | cg13537590 | −0.0500 | 1.0817 × 10−5 | 0.7114 | 0.6614 | IGR | OpenSea | |
| 22:50710746 | cg22416596 | −0.0404 | 4.9783 × 10−5 | 0.6884 | 0.6481 | IGR | Shore |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomez-Verjan, J.C.; Esparza-Aguilar, M.; Martín-Martín, V.; Salazar-Perez, C.; Cadena-Trejo, C.; Gutierrez-Robledo, L.M.; Martínez-Magaña, J.J.; Nicolini, H.; Arroyo, P. Years of Schooling Could Reduce Epigenetic Aging: A Study of a Mexican Cohort. Genes 2021, 12, 1408. https://doi.org/10.3390/genes12091408
Gomez-Verjan JC, Esparza-Aguilar M, Martín-Martín V, Salazar-Perez C, Cadena-Trejo C, Gutierrez-Robledo LM, Martínez-Magaña JJ, Nicolini H, Arroyo P. Years of Schooling Could Reduce Epigenetic Aging: A Study of a Mexican Cohort. Genes. 2021; 12(9):1408. https://doi.org/10.3390/genes12091408
Chicago/Turabian StyleGomez-Verjan, Juan Carlos, Marcelino Esparza-Aguilar, Verónica Martín-Martín, Cecilia Salazar-Perez, Cinthya Cadena-Trejo, Luis Miguel Gutierrez-Robledo, José Jaime Martínez-Magaña, Humberto Nicolini, and Pedro Arroyo. 2021. "Years of Schooling Could Reduce Epigenetic Aging: A Study of a Mexican Cohort" Genes 12, no. 9: 1408. https://doi.org/10.3390/genes12091408