
Select Porcine Elongation Factor 1α Sequences Mediate Stable High-Level and Upregulated Expression of Heterologous Genes in Porcine Cells in Response to Primate Serum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Promoter and Expression Vector Construction
2.2. Cell Culture, Transfection, Selection, Serum Induction, and Genotyping
2.3. Luciferase Assay
2.4. Real-Time Quantitative PCR
2.5. Western Blot Analysis
2.6. Flow Cytometry
2.7. Statistical Analysis
3. Results
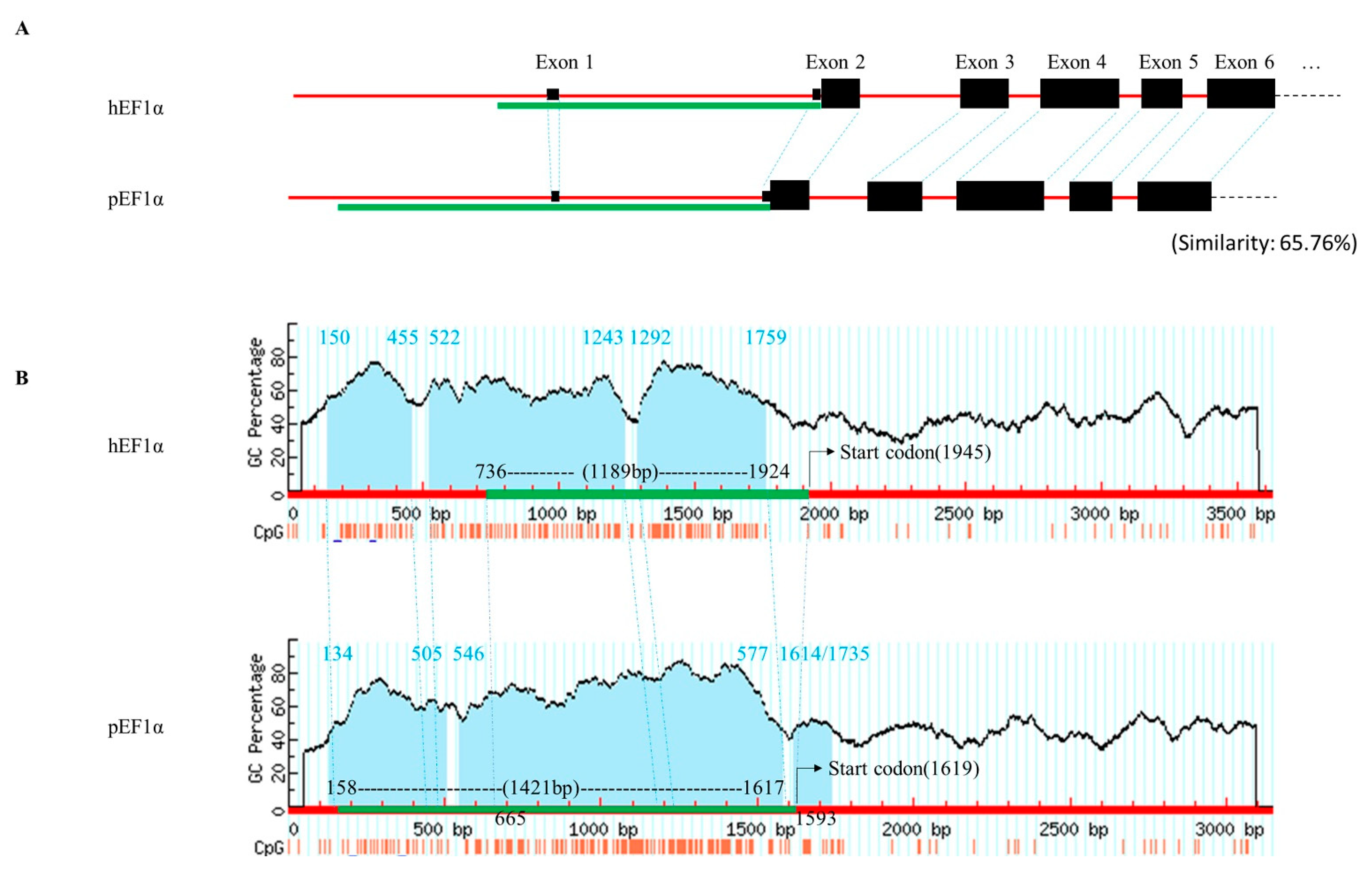
3.1. Sequence Analysis of the pEF1α Upstream Region
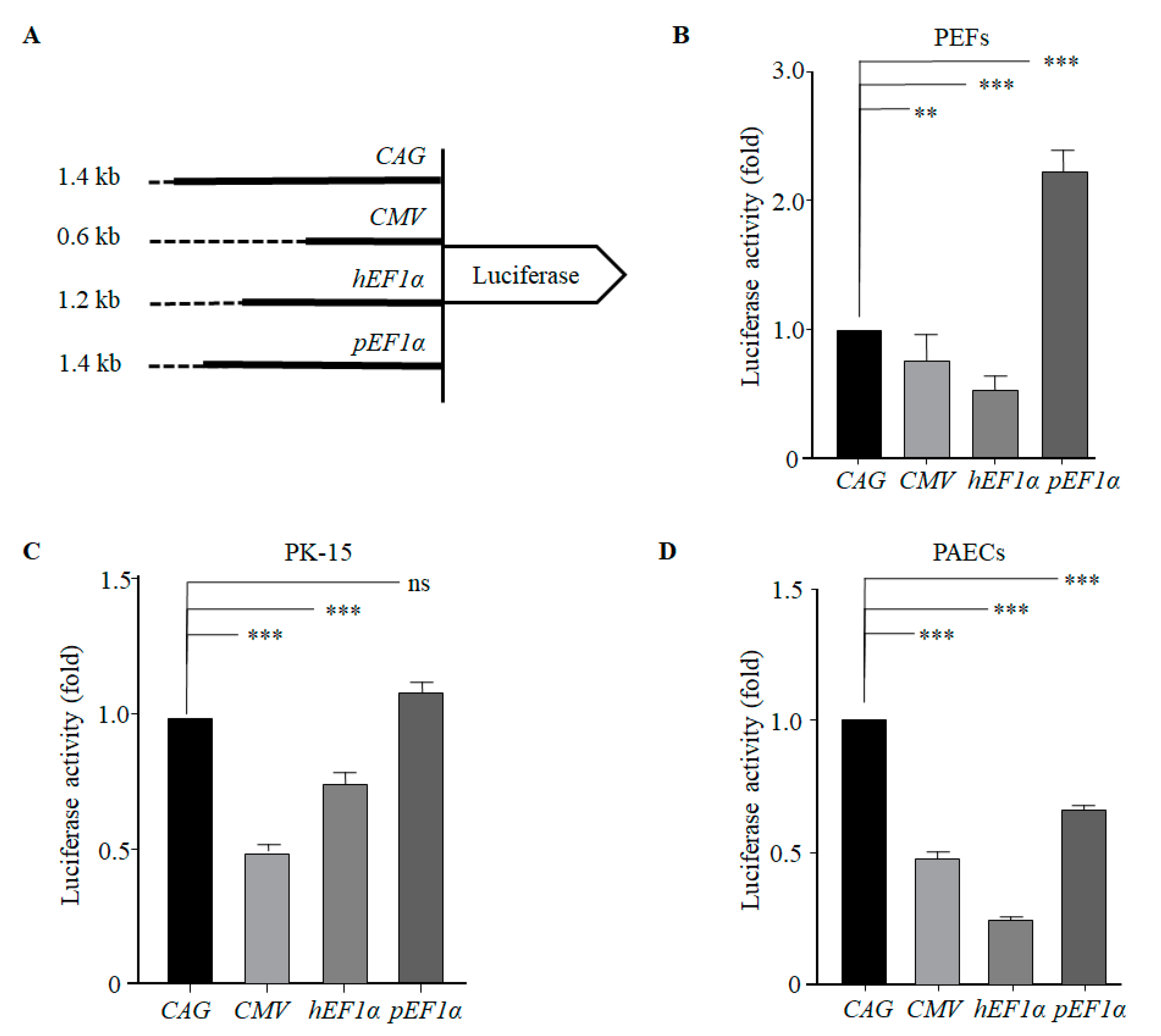
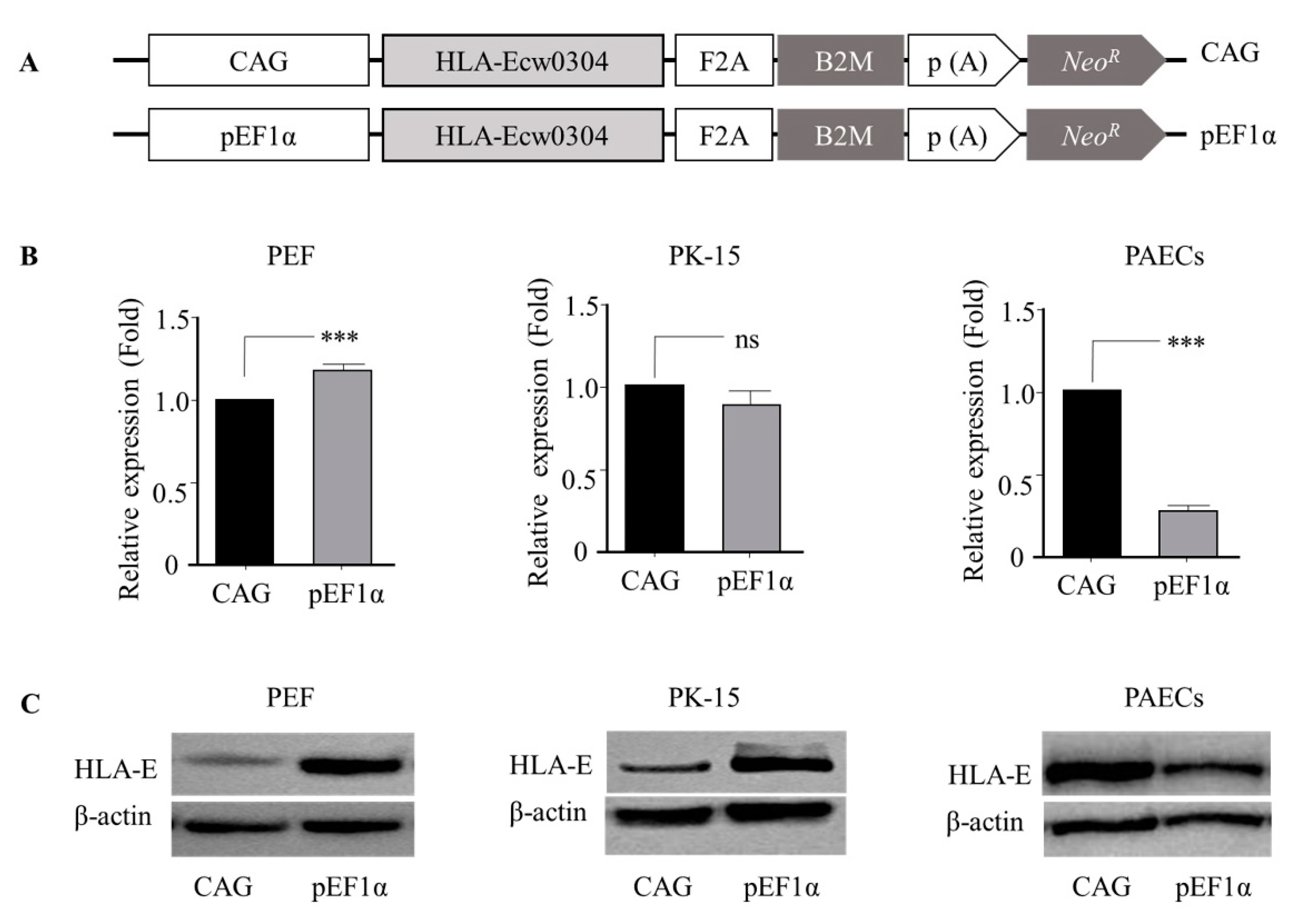
3.2. The pEF1α Promoter Led to High-Level Heterologous Gene Expression in Porcine Cells
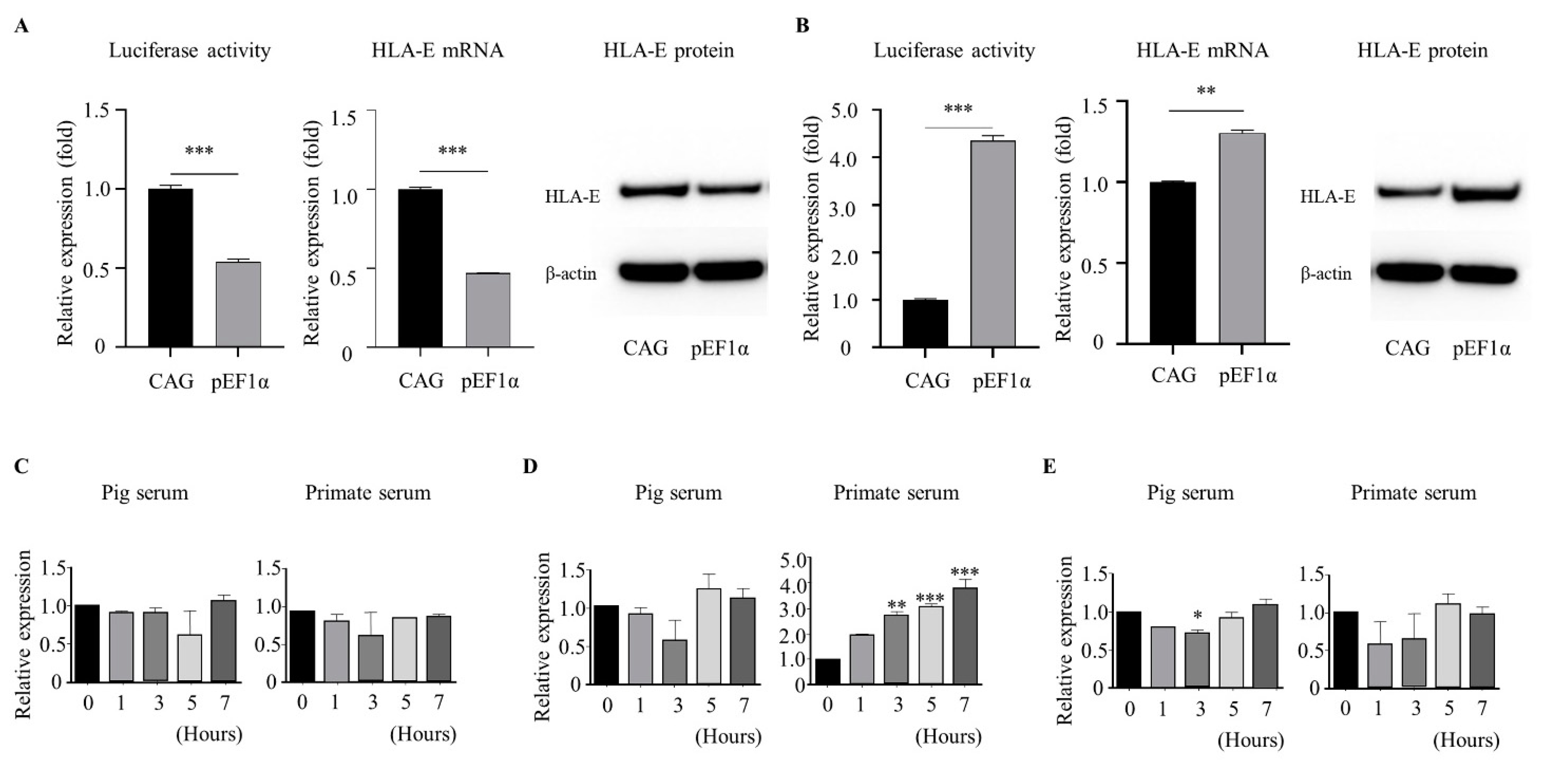
3.3. The pEF1α Promoter Induced Heterologous Gene Expression in Response to Xenogeneic Serum
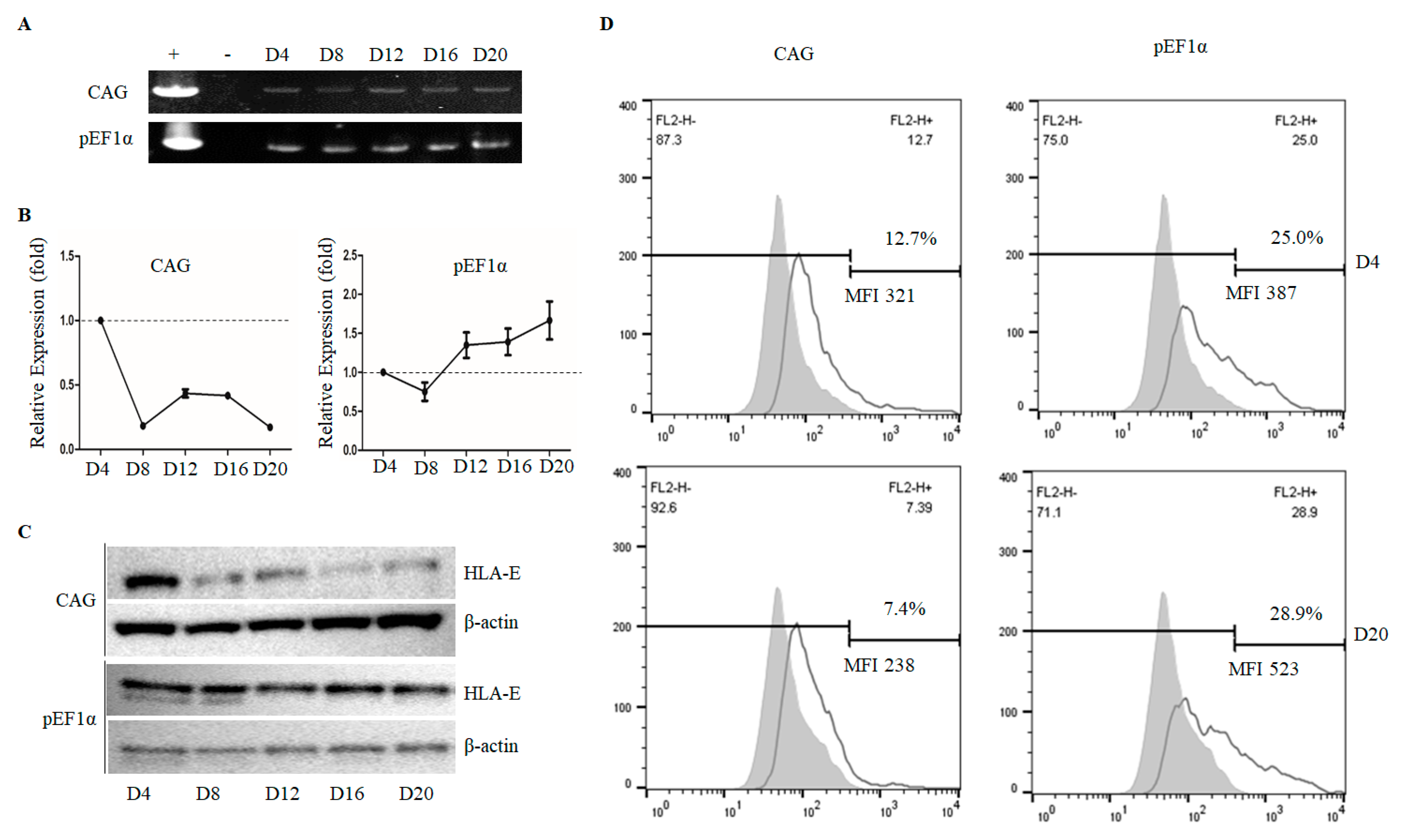
3.4. The pEF1α Promoter, but Not the CAG Promoter, Retained High-Level HLA-E Expression during Long-Term Culture
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cooper, D.K.C.; Hara, H.; Iwase, H.; Yamamoto, T.; Li, Q.; Ezzelarab, M.; Federzoni, E.; Dandro, A.; Ayares, D. Justification of specific genetic modifications in pigs for clinical organ xenotransplantation. Xenotransplantation 2019, 26, e12516. [Google Scholar] [CrossRef]
- Buermann, A.; Petkov, S.; Petersen, B.; Hein, R.; Lucas-Hahn, A.; Baars, W.; Brinkmann, A.; Niemann, H.; Schwinzer, R. Pigs expressing the human inhibitory ligand PD-L1 (CD 274) provide a new source of xenogeneic cells and tissues with low immunogenic properties. Xenotransplantation 2018, 25, e12387. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Xu, W.; Kan, Y.; Zhao, H.Y.; Zhou, Y.; Song, X.; Wu, J.; Xiong, J.; Goswami, D.; Yang, M.; et al. Extensive germline genome engineering in pigs. Nat. Biomed. Eng. 2021, 5, 134–143. [Google Scholar] [CrossRef]
- Fischer, K.; Kraner-Scheiber, S.; Petersen, B.; Rieblinger, B.; Buermann, A.; Flisikowska, T.; Flisikowski, K.; Christan, S.; Edlinger, M.; Baars, W.; et al. Efficient production of multi-modified pigs for xenotransplantation by ‘combineering’, gene stacking and gene editing. Sci. Rep. 2016, 6, 29081. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Li, H.P.; Hung, Y.H.; Leu, Y.W.; Wu, W.H.; Wang, F.S.; Lee, K.D.; Chang, P.J.; Wu, C.S.; Lu, Y.J.; et al. Targeted methylation of CMV and E1A viral promoters. Biochem. Biophys. Res. Commun. 2010, 402, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, T.; Zhang, Q.K.; Jiang, Y.; Xu, D.G.; Zhang, M.; Shen, W.; Pan, Q.J. Unstable expression of transgene is associated with the methylation of CAG promoter in the offspring from the same litter of homozygous transgenic mice. Mol. Biol. Rep. 2014, 41, 5177–5186. [Google Scholar] [CrossRef]
- Aigner, B.; Klymiuk, N.; Wolf, E. Transgenic pigs for xenotransplantation: Selection of promoter sequences for reliable transgene expression. Curr. Opin. Organ Transplant. 2010, 15, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Wuensch, A.; Baehr, A.; Bongoni, A.K.; Kemter, E.; Blutke, A.; Baars, W.; Haertle, S.; Zakhartchenko, V.; Kurome, M.; Kessler, B.; et al. Regulatory sequences of the porcine THBD gene facilitate endothelial-specific expression of bioactive human thrombomodulin in single- and multitransgenic pigs. Transplantation 2014, 97, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Bacman, S.R.; Gammage, P.A.; Minczuk, M.; Moraes, C.T. Chapter 19—Manipulation of mitochondrial genes and mtDNA heteroplasmy. In Methods in Cell Biology; Pon, L.A., Schon, E.A., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 155, pp. 441–487. [Google Scholar]
- Kallunki, T.; Barisic, M.; Jäättelä, M.; Liu, B. How to Choose the Right Inducible Gene Expression System for Mammalian Studies? Cells 2019, 8, 796. [Google Scholar] [CrossRef] [Green Version]
- Varley, A.W.; Coulthard, M.G.; Meidell, R.S.; Gerard, R.D.; Munford, R.S. Inflammation-induced recombinant protein expression in vivo using promoters from acute-phase protein genes. Proc. Natl. Acad. Sci. USA 1995, 92, 5346–5350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.; Manske, K.; Seissler, J.; Wohlleber, D.; Simm, N.; Wolf-van Buerck, L.; Knolle, P.; Schnieke, A.; Fischer, K. Cytokine-inducible promoters to drive dynamic transgene expression: The “Smart Graft” strategy. Xenotransplantation 2020, 27, e12634. [Google Scholar] [CrossRef]
- Dinarello, C.A.; van der Meer, J.W. Treating inflammation by blocking interleukin-1 in humans. Semin. Immunol. 2013, 25, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Grenz, A.; Schenk, M.; Zipfel, A.; Viebahn, R. TNF-alpha and its receptors mediate graft rejection and loss after liver transplantation. Clin. Chem. Lab. Med. 2000, 38, 1183–1185. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Söderström, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Sun, W.S.; Chun, J.L.; Kim, D.H.; Ahn, J.S.; Kim, M.K.; Hwang, I.S.; Kwon, D.J.; Hwang, S.; Lee, J.W. Molecular cloning and characterization of porcine ribosomal protein L21. J. Vet. Sci. 2017, 18, 531–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Hwang, I.S.; Vasamsetti, B.M.K.; Rallabandi, H.R.; Park, M.R.; Byun, S.J.; Yang, H.; Ock, S.A.; Lee, H.C.; Woo, J.S.; et al. Codon optimized membrane cofactor protein expression in α 1, 3 galactosyltransferase knockout pig cells improve protection against cytotoxicity of monkey serum. 3 Biotech 2020, 10, 108. [Google Scholar] [CrossRef]
- Park, M.Y.; Krishna Vasamsetti, B.M.; Kim, W.S.; Kang, H.J.; Kim, D.Y.; Lim, B.; Cho, K.; Kim, J.S.; Chee, H.K.; Park, J.H.; et al. Comprehensive Analysis of Cardiac Xeno-Graft Unveils Rejection Mechanisms. Int. J. Mol. Sci. 2021, 22, 751. [Google Scholar] [CrossRef] [PubMed]
- Iwase, H.; Liu, H.; Wijkstrom, M.; Zhou, H.; Singh, J.; Hara, H.; Ezzelarab, M.; Long, C.; Klein, E.; Wagner, R.; et al. Pig kidney graft survival in a baboon for 136 days: Longest life-supporting organ graft survival to date. Xenotransplantation 2015, 22, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, L.; Zhang, Y.; He, Y.; Yu, H.; Chen, J.; Liu, D.; Lin, S.; Gao, M.; Zhong, G.; Lei, W.; et al. Selective germline genome edited pigs and their long immune tolerance in Non Human Primates. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Lee, R.; Oh, K.B.; Kim, Y.; Woo, J.S.; Hwang, S.; Im, G.S.; Ock, S.A. Stable Regulation of Senescence-Related Genes in Galactose-alpha1,3-galactose Epitope Knockout and Human Membrane Cofactor Protein hCD46 Pig. Transplant. Proc. 2019, 51, 2043–2050. [Google Scholar] [CrossRef]
- Hwang, S.; Oh, K.B.; Kwon, D.J.; Ock, S.A.; Lee, J.W.; Im, G.S.; Lee, S.S.; Lee, K.; Park, J.K. Improvement of cloning efficiency in minipigs using post-tawed donor cells treated with roscovitine. Mol. Biotechnol. 2013, 55, 212–216. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, H.; Oh, K.B.; Hwang, I.S.; Yang, H.; Park, M.R.; Ock, S.A.; Woo, J.S.; Im, G.S.; Hwang, S. Production and Breeding of Transgenic Cloned Pigs Expressing Human CD73. Dev. Reprod. 2017, 21, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himanen, S.V.; Sistonen, L. New insights into transcriptional reprogramming during cellular stress. J. Cell Sci. 2019, 132, jcs238402. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Cheshire, J.K.; Patel, H.; Woo, P. Cross-talk between transcription factors NF-kappa B and C/EBP in the transcriptional regulation of genes. Int. J. Biochem. Cell Biol. 1997, 29, 1525–1539. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Z.; Tian, Z.; Zhang, X.; Xu, D.; Li, Q.; Zhang, J.; Wang, T. The EF-1α promoter maintains high-level transgene expression from episomal vectors in transfected CHO-K1 cells. J. Cell. Mol. Med. 2017, 21, 3044–3054. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Gene Symbol | Amplicon Size (bp) | Primer (5′→3′) |
|---|---|---|---|
| Real-time PCR | hHLA-E | 501 | GGGCTACCCGAGCCCGTCACCCTGAGATGG |
| TTCAATTCTCTCTCCATTCTTCAGTAAGTC | |||
| pGAPDH | 219 | TCGGAGTGAACGGATTTG | |
| CCTGGAAGATGGTGATGG | |||
| pTF | 181 | AACTGAATGTGACCGTAGAAGCTG | |
| CCTTTATCCACGTCAATCAGAAAC | |||
| pEF1α | 136 | TGGATTGCATTCTACCACCA | |
| ACCATGCCAGGTTTGAGAAC | |||
| pICAM2 | 117 | CGTGCTGCTCTTCTTGTTTG | |
| CACCTCAGCCTCCTCTGAAC | |||
| Genotyping | hHLA-E | 1274 | ATTTCCACACTTCCGTGTCC |
| TTCAATTCTCTCTCCATTCTTCAGTAAGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.-S.; Yang, H.; No, J.G.; Lee, H.; Lee, N.; Lee, M.; Kang, M.-J.; Oh, K.B. Select Porcine Elongation Factor 1α Sequences Mediate Stable High-Level and Upregulated Expression of Heterologous Genes in Porcine Cells in Response to Primate Serum. Genes 2021, 12, 1046. https://doi.org/10.3390/genes12071046
Sun W-S, Yang H, No JG, Lee H, Lee N, Lee M, Kang M-J, Oh KB. Select Porcine Elongation Factor 1α Sequences Mediate Stable High-Level and Upregulated Expression of Heterologous Genes in Porcine Cells in Response to Primate Serum. Genes. 2021; 12(7):1046. https://doi.org/10.3390/genes12071046
Chicago/Turabian StyleSun, Wu-Sheng, Hyeon Yang, Jin Gu No, Haesun Lee, Nahyun Lee, Minguk Lee, Man-Jong Kang, and Keon Bong Oh. 2021. "Select Porcine Elongation Factor 1α Sequences Mediate Stable High-Level and Upregulated Expression of Heterologous Genes in Porcine Cells in Response to Primate Serum" Genes 12, no. 7: 1046. https://doi.org/10.3390/genes12071046