
Rice (Oryza sativa) TIR1 and 5′adamantyl-IAA Significantly Improve the Auxin-Inducible Degron System in Schizosaccharomyces pombe


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Cloning and Plasmid Construction
2.3. S. pombe arg3-D4 Integration Expression Vectors
2.4. AID-Tagging Constructs
2.5. Yeast Transformations
2.6. Spot Tests
2.7. Preparation of Total Cell Extract and Western Blot Analysis
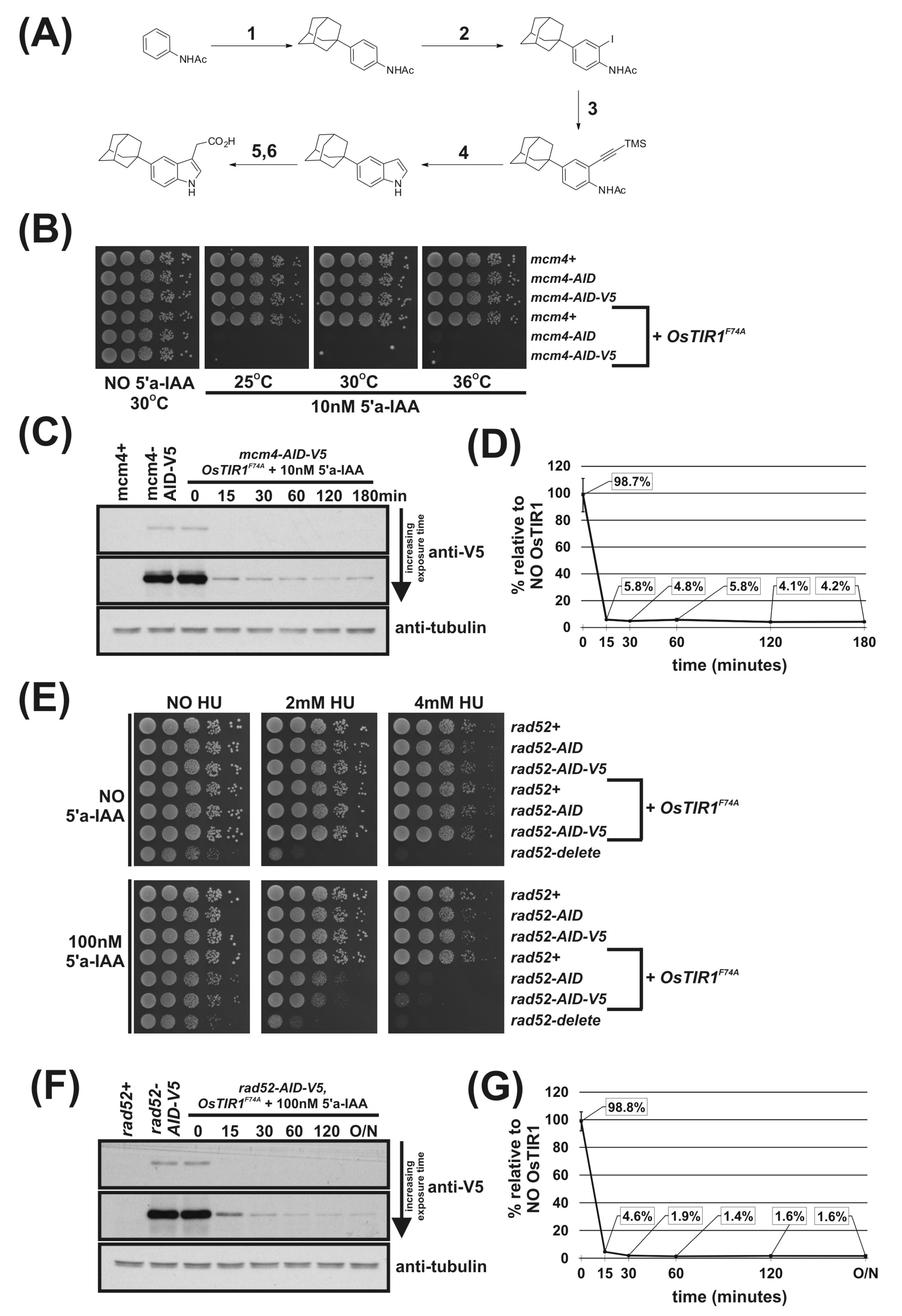
2.8. Synthesis of 2-[5-(adamantan-1-yl)-1H-indol-3-yl]Acetic Acid (5′-adamantyl-IAA)
2.8.1. N-(4-(adamantan-1-yl)phenyl)acetamide

2.8.2. N-(4-(adamantan-1-yl)-2-iodophenyl)acetamide

2.8.3. N-(4-(adamantan-1-yl)-2-((trimethylsilyl)ethynyl)phenyl)acetamide

2.8.4. 5-(Adamantan-1-yl)-1H-indole

2.8.5. Methyl 2-(5-(adamantan-1-yl)-1H-indol-3-yl)-2-oxoacetate

2.8.6. 2-[5-(Adamantan-1-yl)-1H-indol-3-yl]Acetic Acid

3. Results
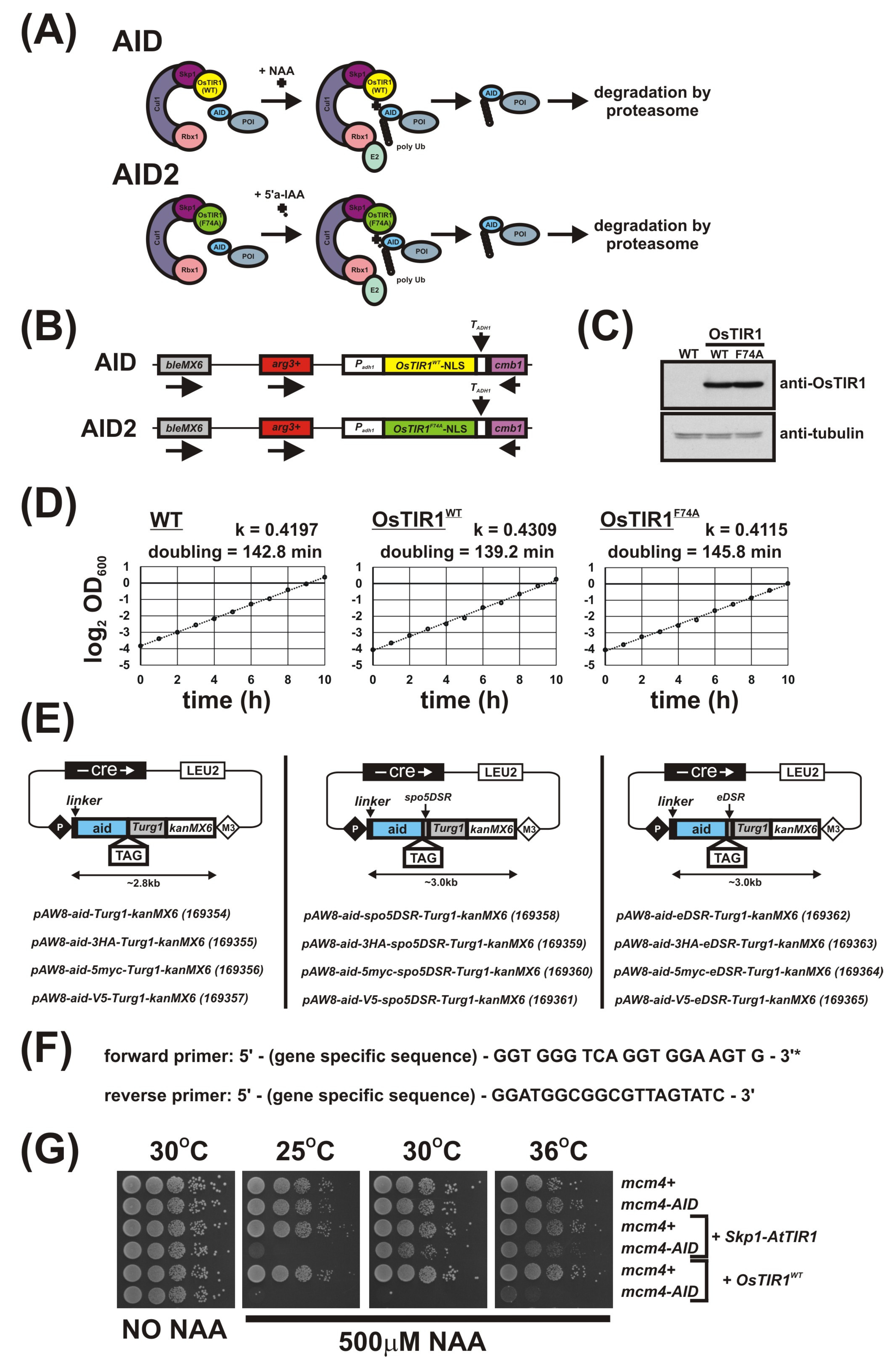
3.1. Integration of OsTIR1WT and OsTIR1F74A Expression Constructs at the arg3-D4 Locus
3.2. Plasmids for C-Terminal AID Tagging
3.3. Testing the New S. pombe AID/OsTIR1(WT) System
3.4. Further Characterisation of the S. pombe AID/OsTIR1WT System
3.5. Using the AID2/OsTIR1F74A System in S. pombe
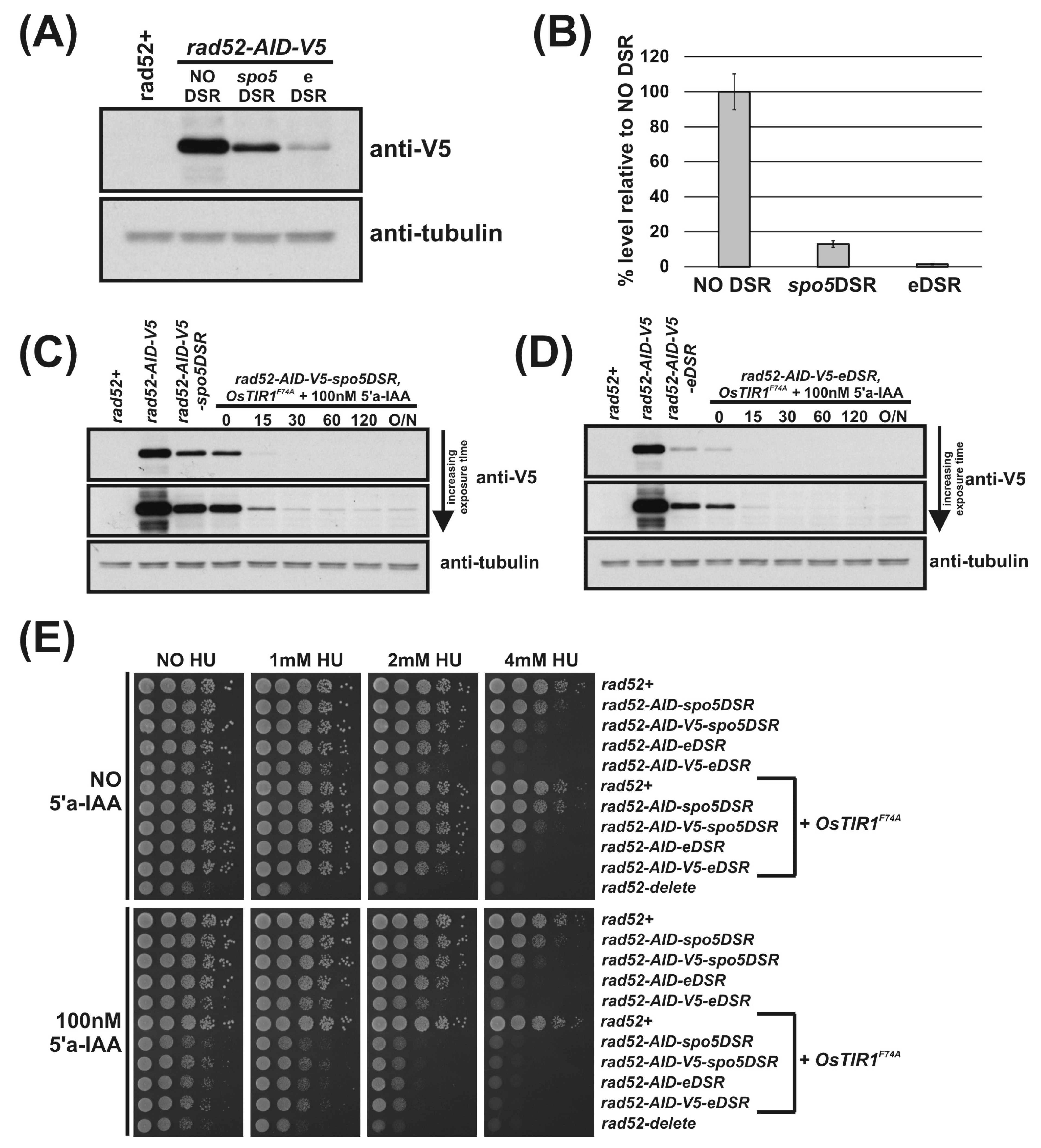
3.6. Use of DSR Elements Allows Generation of Stricter S. pombe Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nishimura, K.; Fukagawa, T.; Takisawa, H.; Kakimoto, T.; Kanemaki, M. An auxin-based degron system for the rapid depletion of proteins in nonplant cells. Nat. Methods 2009, 6, 917–922. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef]
- Costa, E.A.; Subramanian, K.; Nunnari, J.; Weissman, J.S. Defining the physiological role of SRP in protein-targeting efficiency and specificity. Science 2018, 359, 689–692. [Google Scholar] [CrossRef] [Green Version]
- Nora, E.P.; Goloborodko, A.; Valton, A.L.; Gibcus, J.H.; Uebersohn, A.; Abdennur, N.; Dekker, J.; Mirny, L.A.; Bruneau, B.G. Targeted Degradation of CTCF Decouples Local Insulation of Chromosome Domains from Genomic Compartmentalization. Cell 2017, 169, 930–944. [Google Scholar] [CrossRef] [Green Version]
- Muhar, M.; Ebert, A.; Neumann, T.; Umkehrer, C.; Jude, J.; Wieshofer, C.; Rescheneder, P.; Lipp, J.J.; Herzog, V.A.; Reichholf, B.; et al. SLAM-seq defines direct gene-regulatory functions of the BRD4-MYC axis. Science 2018, 360, 800–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yesbolatova, A.; Saito, Y.; Kitamoto, N.; Makino-Itou, H.; Ajima, R.; Nakano, R.; Nakaoka, H.; Fukui, K.; Gamo, K.; Tominari, Y.; et al. The auxin-inducible degron 2 technology provides sharp degradation control in yeast, mammalian cells, and mice. Nat. Commun. 2020, 11, 5701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ward, J.D.; Cheng, Z.; Dernburg, A.F. The auxin-inducible degradation (AID) system enables versatile conditional protein depletion in C. elegans. Development 2015, 142, 4374–4384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bence, M.; Jankovics, F.; Lukácsovich, T.; Erdélyi, M. Combining the auxin-inducible degradation system with CRISPR/Cas9-based genome editing for the conditional depletion of endogenous Drosophila melanogaster proteins. FEBS J. 2017, 284, 1056–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleinjan, D.A.; Wardrope, C.; Nga Sou, S.; Rosser, S.J. Drug-tunable multidimensional synthetic gene control using inducible degron-tagged dCas9 effectors. Nat. Commun. 2017, 8, 1191. [Google Scholar] [CrossRef] [Green Version]
- Kanke, M.; Nishimura, K.; Kanemaki, M.; Kakimoto, T.; Takahashi, T.S.; Nakagawa, T.; Masukata, H. Auxin-inducible protein depletion system in fission yeast. BMC Cell Biol. 2011, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Yamagishi, Y.; Sakuno, T.; Shimura, M.; Watanabe, Y. Heterochromatin links to centromeric protection by recruiting shugoshin. Nature 2008, 455, 251–255. [Google Scholar] [CrossRef]
- Kanke, M.; Kodama, Y.; Takahashi, T.S.; Nakagawa, T.; Masukata, H. Mcm10 plays an essential role in origin DNA unwinding after loading of the CMG components. EMBO J. 2012, 31, 2182–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handa, T.; Kanke, M.; Takahashi, T.S.; Nakagawa, T.; Masukata, H. DNA polymerization-independent functions of DNA polymerase epsilon in assembly and progression of the replisome in fission yeast. Mol. Biol. Cell 2012, 23, 3240–3253. [Google Scholar] [CrossRef]
- Kakui, Y.; Rabinowitz, A.; Barry, D.J.; Uhlmann, F. Condensin-mediated remodeling of the mitotic chromatin landscape in fission yeast. Nat. Genet. 2017, 49, 1553–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moiseeva, V.; Amelina, H.; Collopy, L.C.; Armstrong, C.A.; Pearson, S.R.; Tomita, K. The telomere bouquet facilitates meiotic prophase progression and exit in fission yeast. Cell Discov. 2017, 3, 17041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyan, K.M.; McKenna, B.D.; Anderson, W.D.; Duarte, F.M.; Core, L.; Guertin, M.J. An improved auxin-inducible degron system preserves native protein levels and enables rapid and specific protein depletion. Genes Dev. 2019, 33, 1441–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, N.; Takahashi, K.; Iwasaki, R.; Yamada, R.; Yoshimura, M.; Endo, T.A.; Kimura, S.; Zhang, H.; Nomoto, M.; Tada, Y.; et al. Chemical hijacking of auxin signaling with an engineered auxin-TIR1 pair. Nat. Chem. Biol. 2018, 14, 299–305. [Google Scholar] [CrossRef]
- Yamada, R.; Murai, K.; Uchida, N.; Takahashi, K.; Iwasaki, R.; Tada, Y.; Kinoshita, T.; Itami, K.; Torii, K.U.; Hagihara, S. A Super Strong Engineered Auxin-TIR1 Pair. Plant Cell Physiol. 2018, 59, 1538–1544. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, K.; Yamada, R.; Hagihara, S.; Iwasaki, R.; Uchida, N.; Kamura, T.; Takahashi, K.; Torii, K.U.; Fukagawa, T. A super-sensitive auxin-inducible degron system with an engineered auxin-TIR1 pair. Nucleic Acids Res. 2020, 48, e108. [Google Scholar] [CrossRef] [PubMed]
- Harigaya, Y.; Tanaka, H.; Yamanaka, S.; Tanaka, K.; Watanabe, Y.; Tsutsumi, C.; Chikashige, Y.; Hiraoka, Y.; Yamashita, A.; Yamamoto, M. Selective elimination of messenger RNA prevents an incidence of untimely meiosis. Nature 2006, 442, 45–50. [Google Scholar] [CrossRef]
- Yamanaka, S.; Yamashita, A.; Harigaya, Y.; Iwata, R.; Yamamoto, M. Importance of polyadenylation in the selective elimination of meiotic mRNAs in growing S. pombe cells. EMBO J. 2010, 29, 2173–2181. [Google Scholar] [CrossRef]
- Yamashita, A.; Shichino, Y.; Tanaka, H.; Hiriart, E.; Touat-Todeschini, L.; Vavasseur, A.; Ding, D.Q.; Hiraoka, Y.; Verdel, A.; Yamamoto, M. Hexanucleotide motifs mediate recruitment of the RNA elimination machinery to silent meiotic genes. Open Biol. 2012, 2, 120014. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.T.; Daigaku, Y.; Mohebi, S.; Etheridge, T.J.; Chahwan, C.; Murray, J.M.; Carr, A.M. Optimisation of the Schizosaccharomyces pombe urg1 expression system. PLoS ONE 2013, 8, e83800. [Google Scholar] [CrossRef] [Green Version]
- Waddell, S.; Jenkins, J.R. arg3+, a new selection marker system for Schizosaccharomyces pombe: Application of ura4+ as a removable integration marker. Nucleic Acids Res. 1995, 23, 1836–1837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, S.; Klar, A.; Nurse, P. Molecular Genetic Analysis of Fission Yeast Schizosaccharomyces Pombe; Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology; Elsevier: Amsterdam, Netherlands, 1991; pp. 795–823. [Google Scholar]
- Zach, R.; Carr, A.M. Increased expression of Polδ does not alter the canonical replication program in vivo. Wellcome Open Res. 2021, 6, 44. [Google Scholar] [CrossRef]
- Watson, A.T.; Garcia, V.; Bone, N.; Carr, A.M.; Armstrong, J. Gene tagging and gene replacement using recombinase-mediated cassette exchange in Schizosaccharomyces pombe. Gene 2008, 407, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Bähler, J.; Wu, J.Q.; Longtine, M.S.; Shah, N.G.; McKenzie, A.; Steever, A.B.; Wach, A.; Philippsen, P.; Pringle, J.R. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 1998, 14, 943–951. [Google Scholar] [CrossRef]
- Hentges, P.; Van Driessche, B.; Tafforeau, L.; Vandenhaute, J.; Carr, A.M. Three novel antibiotic marker cassettes for gene disruption and marker switching in Schizosaccharomyces pombe. Yeast 2005, 22, 1013–1019. [Google Scholar] [CrossRef]
- Watson, A.T.; Werler, P.; Carr, A.M. Regulation of gene expression at the fission yeast Schizosaccharomyces pombe urg1 locus. Gene 2011, 484, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Yokobayashi, S.; Watanabe, Y. The kinetochore protein Moa1 enables cohesion-mediated monopolar attachment at meiosis I. Cell 2005, 123, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Gray, W.M.; del Pozo, J.C.; Walker, L.; Hobbie, L.; Risseeuw, E.; Banks, T.; Crosby, W.L.; Yang, M.; Ma, H.; Estelle, M. Identification of an SCF ubiquitin-ligase complex required for auxin response in Arabidopsis thaliana. Genes Dev. 1999, 13, 1678–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraschin, F.S.; Memelink, J.; Offringa, R. Auxin-induced, SCF(TIR1)-mediated poly-ubiquitination marks AUX/IAA proteins for degradation. Plant J. 2009, 59, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Morawska, M.; Ulrich, H.D. An expanded tool kit for the auxin-inducible degron system in budding yeast. Yeast 2013, 30, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Fukagawa, T. An efficient method to generate conditional knockout cell lines for essential genes by combination of auxin-inducible degron tag and CRISPR/Cas9. Chromosome Res. 2017, 25, 253–260. [Google Scholar] [CrossRef]
- Zasadzińska, E.; Huang, J.; Bailey, A.O.; Guo, L.Y.; Lee, N.S.; Srivastava, S.; Wong, K.A.; French, B.T.; Black, B.E.; Foltz, D.R. Inheritance of CENP-A Nucleosomes during DNA Replication Requires HJURP. Dev. Cell 2018, 47, 348–362.e7. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.L.; Osman, F.; Dixon, J.; Whitby, M.C. DNA repair by a Rad22-Mus81-dependent pathway that is independent of Rhp51. Nucleic Acids Res. 2004, 32, 5570–5581. [Google Scholar] [CrossRef] [Green Version]
- Marguerat, S.; Schmidt, A.; Codlin, S.; Chen, W.; Aebersold, R.; Bähler, J. Quantitative analysis of fission yeast transcriptomes and proteomes in proliferating and quiescent cells. Cell 2012, 151, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Kubota, T.; Nishimura, K.; Kanemaki, M.T.; Donaldson, A.D. The Elg1 replication factor C-like complex functions in PCNA unloading during DNA replication. Mol. Cell 2013, 50, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Miyazawa-Onami, M.; Iida, T.; Araki, H. iAID: An improved auxin-inducible degron system for the construction of a ‘tight’ conditional mutant in the budding yeast Saccharomyces cerevisiae. Yeast 2015, 32, 567–581. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genotype (h+/h- Denote Mating Type) |
|---|---|
| AW279 | h+ |
| AW1502 | arg3-D4 h+ |
| AW1581 | rad52::kanMX6 h+ |
| AW1655 | bleMX6-arg3-D4 h- |
| AW1658 | arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h- |
| AW1660 | arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h- |
| AW1680 | rad52-HA-AID-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h- |
| AW1681 | rad52-HA-AID-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1, h+ |
| AW1682 | rad52-HA-AID-spo5DSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1, h+ |
| AW1683 | rad52-HA-AID-spo5DSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1, h- |
| AW1703 | arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1762 | arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h+ |
| AW1893 | mcm4-AID-Turg1:kanMX6, h+ |
| AW1899 | mcm4-AID-V5-Turg1:kanMX6, h+ |
| AW1901 | rad52-AID-Turg1:kanMX6, h+ |
| AW1907 | rad52-AID-V5-Turg1:kanMX6, h+ |
| AW1915 | rad52-AID-spo5DSR-Turg1:kanMX6 h+ |
| AW1921 | rad52-AID-V5-spo5DSR-Turg1:kanMX6 h+ |
| AW1923 | mcm4-AID-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h+ |
| AW1925 | mcm4-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1935 | mcm4-AID-V5-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h+ |
| AW1937 | mcm4-AID-V5-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1942 | ade6::ade6+-Padh15-skp1-AtTIR1-2NLS-9myc h+ |
| AW1949 | mcm4-AID-Turg1:kanMX6, ade6::ade6+-Padh15-skp1-AtTIR1-2NLS-9myc h+ |
| AW1962 | rad52-AID-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1968 | rad52-AID-V5-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1970 | rad52-AID-spo5DSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1976 | rad52-AID-V5-spo5DSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW1990 | rad52-AID-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h+ |
| AW1996 | rad52-AID-V5-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1WT-TADH1 h+ |
| AW2011 | rad52-AID-eDSR-Turg1:kanMX6 h+ |
| AW2017 | rad52-AID-V5-eDSR-Turg1:kanMX6 h+ |
| AW2024 | rad52-AID-eDSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| AW2026 | rad52-AID-V5-eDSR-Turg1:kanMX6, arg3::bleMX6-arg3+-Padh1-OsTIR1F74A-TADH1 h+ |
| Name | Sequence (5′ to 3′) |
|---|---|
| P1 | AGAAATCTATAGAAAAAAAGCTAGCGTGACGCAGACA |
| P2 | CTCCCGGGAGTGCATGCCAGCATATGTATGTGGTTAGAAAAAAGAAAAGACTTAAAAG |
| P3 | AGAAATCTATAGAAAAAAAGCTAGCGTGACGCAGACATTCGAATGGCATGCCC |
| P4 | CTCCCGGGAGTGCATGCCAGCATATGTATGTGGTTAGAAAAAAGAAAAGACTTAAAAGTTTGTGATAGTCAAGACAATGGAATTCTCTTGCTTAAAGAAAAGCGAAGGCA |
| P5 | GAGAGCTCCGTCTGCAGCGAGTCGACACTTCTAAATAAGCGAATTTCTTATGATTTATGA |
| P6 | ACACTCTACTTGCCCAGATCACTAGTATATTACCCTGTTATCCCTAGCG |
| P7 | CATACATTATACGAAGTTATGCATGCTCGGTGGGTCAGGTGGAAGTGGATCTGGTGCTATGATGGGCAGTGTCGA |
| P8 | AAATTCGCTTATTTAGAAGTGCTAGCTCAAGCTCTGCTCTTGCACTTC |
| P9 | AGTGCAAGAGCAGAGCTTGAGCTAGCAGTCCCGGGGCTTGCCCATCTGTTTTAGACGT |
| P10 | GGGGACGAGGCAAGCTAAACAGATCTCGAGAAGAAGGCCCCGCTG |
| P11 | AGTGCAAGAGCAGAGCTTGAGCTAGCACTACGCCATATCATGCCCA |
| P12 | TCTAAAACAGATGGGCAAGCCCCGGGGCTTTGTCTAACAGGTTTTATGTTGGTTTAAGT |
| P13 | AGAAGTGCAAGAGCAGAGCTGGTGCTGGAGCAGGCGCCTACCCATACGATGTTCCTGACTATGCG |
| P14 | GATGGGCAAGCCCCGGGACTGCTAGCGGCGCGCCTCAGCACTGAGC |
| P15 | TGGGCATGATATGGCGTAGTGCTAGCGGCGCGCCTCAGCACTGAGC |
| P16 | AGAAGTGCAAGAGCAGAGCTGGAGCAGGCGCCGGTGAACAAAAGTTGATTTCTGAAG |
| P17 | GATGGGCAAGCCCCGGGACTGCTAGCGGCGCGCCTTACAAGTCTTCCTC |
| P18 | TGGGCATGATATGGCGTAGTGCTAGCGGCGCGCCTTACAAGTCTTCCTC |
| P19 | TATACATTATTTAATACTAGTCCTAACTGACACAGTACAATATTCATTATTTCTATGCAAGCCAGTTCAATATCTTGATTGTTTAGCTTGCCTCGTCCCC |
| P20 | TCATCAGGGTCATTGGGACTATTCAACGCGAAATCAAATAGAAAACAAAATATTGTTTCAAAAGAATGCTTTCATGTATAGGATGGCGGCGTTAGTATCG |
| P21 | GTGCTTTGGAAAGGCGAGGACGTATTAAGGTTATTACCAGTGCTGGACATCGCATTGTACGTTCAATTGCACAGACTGATGGTGGGTCAGGTGGAAGTG |
| P22 | ATTATGCTCTGTAGTCTTTGATTTTCAACAACGACCTATGGTTTATGGCTCATGAATAATACCAGCTTATTCGCTAAAAAAGGATGGCGGCGTTAGTATC |
| P23 | GAACAAATTCTGATCCTCAGTCGGCAATGAGGTCGCGAGAAAACTACGATGCTACGGTGGATAAGAAAGCCAAAAAAGGAGGTGGGTCAGGTGGAAGTG |
| P24 | AGATCTACCGTTTAAACAAATCATTAGTCATAAAACAGAAAATACTTGGTAAAAAACAAGTTGCCAATCATCACATTTTGCCTCATTACTTGGATGGCGGCGTTAGTATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watson, A.T.; Hassell-Hart, S.; Spencer, J.; Carr, A.M. Rice (Oryza sativa) TIR1 and 5′adamantyl-IAA Significantly Improve the Auxin-Inducible Degron System in Schizosaccharomyces pombe. Genes 2021, 12, 882. https://doi.org/10.3390/genes12060882
Watson AT, Hassell-Hart S, Spencer J, Carr AM. Rice (Oryza sativa) TIR1 and 5′adamantyl-IAA Significantly Improve the Auxin-Inducible Degron System in Schizosaccharomyces pombe. Genes. 2021; 12(6):882. https://doi.org/10.3390/genes12060882
Chicago/Turabian StyleWatson, Adam T., Storm Hassell-Hart, John Spencer, and Antony M. Carr. 2021. "Rice (Oryza sativa) TIR1 and 5′adamantyl-IAA Significantly Improve the Auxin-Inducible Degron System in Schizosaccharomyces pombe" Genes 12, no. 6: 882. https://doi.org/10.3390/genes12060882